Article body
La fibrose est une réponse adaptative pathologique qui détruit non spécifiquement les tissus. Il s’agit d’un processus universel de réparation des tissus qui survient en réaction à de nombreux types d’agressions telles les contraintes mécaniques [1], les brûlures, les radiations ionisantes, l’ischémie, l’inflammation. Ces agressions concourent de manière intriquée à la physiopathologie des maladies infectieuses, tumorales ou auto-immunes, de l’hypertension artérielle et des maladies cardio-vasculaires [2]. Le terme de fibrose décrit précisément l’accumulation nouvelle de protéines de la matrice extra-cellulaire selon un arrangement spatial fibrillaire caractéristique. Il s’agit essentiellement de molécules de collagènes de type I et III (voire de type II, V ou XI) synthétisées sous forme de triples hélices elles-mêmes assemblées en fibrilles par les cellules fibroblastiques [3]. L’apparition des collagènes fibrillaires marque un changement qualitatif et quantitatif de composition des collagènes des tissus [4]. Réciproquement, ces changements de la matrice extracellulaire influencent le phénotype des cellules qui y résident. Ainsi dans le rein normal, le collagène de type I n’existe que dans l’adventice artériel [5]. Son apparition au sein des autres structures de cet organe marque une fibrose tubulo-interstitielle (Figure 1) qui constitue le meilleur marqueur pronostic défavorable d’une évolution vers l’insuffisance rénale terminale, et ce quelle que soit la maladie causale [6]. Ainsi, comme lors des fibroses compliquant les hépatopathies et les pneumopathies chroniques, les séquelles de brûlures ou d’abrasions cutanées-muqueuses ou encore le remodelage cardiaque et vasculaire, le réarrangement de la géométrie de la matrice extracellulaire altère l’organisation fonctionnelle du tissu considéré [4]. De ce fait, ce processus de réparation a des effets fonctionnels délétères qui constituent un enjeu médical majeur. Les fibrilles de collagène ont des capacités d’auto-assemblage qui sont aussi catalysées et stabilisées ou au contraire empêchées par les enzymes de la matrice extracellulaire. Le développement de la fibrose ou sa régression dépend donc ainsi du bilan des équilibres biologiques de ces mécanismes. Il est donc crucial de caractériser les changements extracellulaires et cellulaires qui font du restutio ad integrum de l’architecture et de la fonction tissulaire un défi biomédical.
Figure 1
Image multiphoton d’un rein de souris fibrosé.
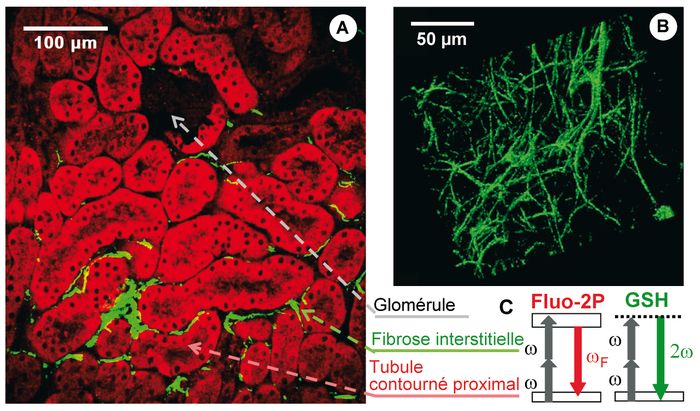
A. Image GSH/fluo-2P d’une coupe épaisse non colorée de cortex rénal. B. Reconstruction 3D de la fibrose interstitielle. C. Codes couleurs pour les modes de contraste multiphoton.
Pour ce faire, un certain nombre d’obstacles méthodologiques sont à surmonter. En effet, les outils de mesure de la fibrose sont peu (coloration au rouge sirius en lumière polarisée couplée à une analyse d’image) ou non quantitatifs (immunohistochimie), ils nécessitent un prélèvement par nature invasif du tissu à analyser et enfin ils ne permettent pas une vision tridimensionnelle des structures et processus dans les tissus complexes.
Dans ce contexte, l’étude des collagènes fibrillaires par la génération de second harmonique (GSH) apparaît très prometteuse. Ce phénomène d’optique non-linéaire consiste en la conversion d’une source laser intense impulsionnelle, de fréquence ω, en son harmonique à 2ω. Au niveau moléculaire, cette réponse quadratique est liée en général à la présence d’électrons polarisables entre un groupement donneur et un groupement accepteur d’électrons. Au niveau macroscopique, la construction cohérente du signal impose l’absence de centrosymétrie, et est plus efficace pour une forte densité d’«harmonophores », puisque le signal est proportionnel au carré de leur densité. En pratique, la GSH est efficace dans des macrostructures denses et ordonnées. Au plan biologique, des signaux GSH intrinsèques ont ainsi été mis en évidence dès les années 1980 dans le collagène fibrillaire [7], et plus récemment dans les myofilaments, les microtubules orientés parallèlement et l’amidon [8].
L’intérêt du signal GSH du collagène fibrillaire réside alors dans son observation en microscopie multiphoton à balayage laser. Ce type de microscopie développé à l’origine par W. Webb et al. (Université Cornell, Ithaca, NY, États-Unis) [9] sur la base de signaux de fluorescence excitée à deux photons (Fluo-2P), s’est peu à peu généralisé à d’autres phénomènes optiques non-linéaires dont la GSH [8]. Il s’agit d’une alternative à la microscopie confocale où la localisation tridimensionnelle du signal provient du fait que les signaux multiphoton dépendent quadratiquement de l'intensité d'excitation. Le principal intérêt de la microscopie multiphoton réside alors dans l’absence de dommages optiques hors du point focal, et dans sa robustesse par rapport aux phénomènes de diffusion qui permet d’imager des tissus biologiques à des profondeurs inégalées en microscopie confocale.
La microscopie multiphoton fondée sur le signal de GSH intrinsèque du collagène fibrillaire semble ainsi la méthode la plus appropriée pour visualiser l’architecture des fibroses, et ceci pour les raisons suivantes :
grande spécificité pour le collagène fibrillaire [10], ce qui permet de quantifier le degré de fibrose ;
faible caractère invasif, lié à la nature non absorptive du processus de GSH, à l’utilisation de sources infrarouges dans la fenêtre de transmission des tissus, et à l’absence de marquage exogène ;
imagerie tridimensionnelle en profondeur ;
possibilité de combiner le signal GSH du collagène à d’autres contrastes, notamment les signaux Fluo-2P d’espèces endogènes telles que le NAD(P)H, les flavines ou l’élastine ou de constructions transgéniques exprimant des sondes fluorescentes métaboliques ou des gènes rapporteurs fluorescents tels que celui de la protéine fluorescente verte (GFP) sous le contrôle de promoteurs d’intérêts.
La Figure 1 illustre ainsi comment la microscopie multiphoton permet de localiser une fibrose interstitielle dans le cortex rénal et de mettre en évidence son architecture tridimensionnelle. Cette méthodologie s’applique dans d’autres contextes, par exemple pour analyser une fibrose pulmonaire
Ainsi, une approche multidisciplinaire, par la confrontation de diverses méthodes de variation aux niveaux cellulaire et extracellulaire, devrait permettre d’améliorer les capacités d’étude de la physiopathologie des fibroses. L’identification des acteurs de telles réorganisations matricielles et de leur retentissement fonctionnel délétère pourrait permettre de sélectionner de futures cibles thérapeutiques pour traiter et prévenir ce processus dégénératif universel.
Appendices
Remerciements
Nous remercions vivement la Fondation pour la Recherche Médicale et la Région Ile de France pour leur soutien à Ana-Maria Pena et Mathias Strupler, respectivement.
Références
- 1. Breen EC. Mechanical strain increases type I collagen expression in pulmonary fibroblasts in vitro. J Appl Physiol 2000 ; 88 : 203-9.
- 2. Hinglais N, Heudes D, Nicoletti A, et al. Colocalization of myocardial fibrosis and inflammatory cells in rats. Lab Invest 1994 ; 70 : 286-94.
- 3. Hulmes DJS. Building collagen molecules, fibrils, and suprafibrillar strcutures. J Struct Biol 2002 ; 137 : 2-10.
- 4. Weber KT. Fibrosis, a common pathway to organ failure: angiotensin II and tissue repair. Semin Nephrol 1997 ; 17 : 467-91.
- 5. Yoshioka K, Tohda M, Takemura T, et al. Distribution of type I collagen in human kidney diseases in comparison with type III collagen. J Pathol 1990 ; 162 : 141-8.
- 6. Nath KA. Tubulointerstitial changes as a major determinant in the progression of renal damage. Am J Kidney Dis 1992 ; 20 : 1-17.
- 7. Freund I, Deutsch M, Sprecher A. Optical second-harmonic microscopy, crossed-beam summation and small-angle scattering in rat-tail tendon. Biophys J 1986 ; 50 : 693-712.
- 8. Zipfel WR, Williams RM, Christie R, et al. Live tissue intrinsic emission microscopy using multiphoton-excited native fluorescence and second harmonic generation. Proc Natl Acad Sci USA 2003 ; 100 : 7075-80.
- 9. Denk W, Strickler JH, Webb WW. Two-photon laser scanning microscopy. Science 1990 ; 248 : 73-6.
- 10. Pena AM, Boulesteix T, Dartigalongue T, Schanne-Klein MC. Chiroptical effects in the second harmonic signal of collagens I and IV. J Am Chem Soc 2005 ; 127 : 10314-22.
List of figures
Figure 1
Image multiphoton d’un rein de souris fibrosé.
A. Image GSH/fluo-2P d’une coupe épaisse non colorée de cortex rénal. B. Reconstruction 3D de la fibrose interstitielle. C. Codes couleurs pour les modes de contraste multiphoton.