Article body
La sortiline est une protéine d’environ 100 kDa qui possède un seul domaine transmembranaire. Le nombre de partenaires auxquels cette protéine est capable de s’associer est impressionnant : d’abord la RAP (receptor associated protein), une protéine de 40 kDa qui a servi à purifier et à cloner pour la première fois le gène codant pour la sortiline [1]. Puis, la neurotensine (NT), un neuropeptide de 13 acides aminés grâce auquel nous avons purifié par chromatographie d’affinité l’un des récepteur de la NT, le NTR3, qui s’avéra être identique à la sortiline [2]. Vient ensuite la lipoprotéine lipase (LpL), une protéine de 50 kDa que la sortiline est capable de lier et d’internaliser [3], puis le propeptide de 44 acides aminés libéré par la furine lors de la maturation de la prosortiline [4]. Enfin, le proNGF (précurseur du NGF, nerve growth factor, une protéine de 31 kDa) interagit lui aussi avec la sortiline pour délivrer un signal de mort cellulaire [5].
La structure primaire de la sortiline, et notamment la petite taille de sa queue carboxyterminale cytoplasmique, lui interdit toute possibilité de transmettre seule un signal intracellulaire. La recherche de ses activités à la surface cellulaire passait donc obligatoirement par l’identification de partenariats avec d’autres protéines membranaires. C’est dans ce contexte que se situent les résultats obtenus par A. Nykjaer et al. qui montrent que les effets proapoptotiques du proNGF passent par une interaction directe entre le p75NTR, l’un des récepteur du NGF, et la sortiline [5].
Alors que le NGF était connu comme un facteur de survie, de croissance et de différenciation des neurones, le proNGF, libéré comme tel [6], était décrit comme un facteur de mort cellulaire. L’action positive du NGF sur la survie et la croissance cellulaires requiert son association physique à deux récepteurs des neurotrophines, le récepteur p75NTR et TrkA, un autre récepteur du NGF. Le problème était que la mise en jeu du p75NTR semblait aussi responsable de la mort de plusieurs types de cellules dont les neurones. Cette contradiction est résolue par A. Nykjaer, qui démontre clairement que la sortiline est le co-récepteur du p75NTR pour l’action proapoptotique activée par le proNGF alors que l’association du NGF au complexe TrKA-p75NTR conduit à la survie cellulaire. De façon remarquable, l’effet proapoptotique du proNGF lié au complexe sortiline-p75NTR peut être inhibé par la NT, ce qui prouve sans ambiguïté l’importance de la sortiline dans cette fonction. Ainsi le NGF et son précurseur sont capables de moduler l’équilibre entre la survie et la mort cellulaire selon le complexe moléculaire auquel ils se lient.
Cette association fonctionnelle de la sortiline avec un autre récepteur membranaire n’est pas la première décrite. En effet, la sortiline forme également un complexe avec le récepteur 1 de la NT (NTR1, un récepteur à sept domaines transmembranaires) dans les cellules d’adénocarcinome de côlon humain HT29 [7]. Dans ce cas, la sortiline module l’intensité du signal induit par la NT puisque l’interaction sortiline-NTR1 diminue l’activité de la voie de signalisation intrinsèque du NTR1, activité qui fait intervenir les MAP kinases et la phospholipase C. D’autre part, dans les cellules microgliales humaines qui expriment la sortiline, la NT est capable d’activer la migration cellulaire par un mécanisme qui emprunte la voie de la protéine-kinase B (PI3-kinase) [8]. Le partenaire éventuel de la sortiline impliqué dans cet effet n’a pas encore été identifié (Figure 1).
Figure 1
La sortiline, un co-récepteur associé à de multiples fonctions.
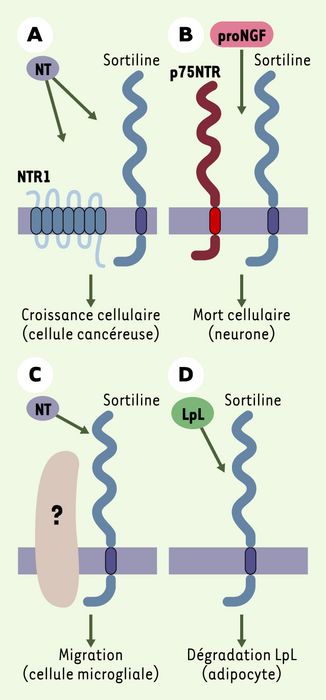
A. La sortiline est associée avec le NTR1 pour induire les effets de prolifération de la NT dans les cellules cancéreuses (HT29). B. A l’inverse, le complexe p75NTR/sortiline formé dans les neurones est responsable de la mort cellulaire provoquée par le proNGF. C. Dans les cellules microgliales humaines, la NT active la migration cellulaire par l’intermédiaire de la sortiline, probablement en association avec une protéine non identifiée capable de transduire le signal. D. Dans les adipocytes, la sortiline internalise la LpL (lipoprotéine lipase) et permet ainsi sa dégradation. Cette fonction ne nécessite pas forcément l’association avec un partenaire.
Les propriétés décrites ci-dessus sont la conséquence de la présence de la sortiline à la surface cellulaire. Cependant, cette expression membranaire ne correspond qu’à environ 10 % du contenu cellulaire en sortiline, le reste de la protéine étant localisé au niveau du Golgi [1]. C’est cette fraction intracellulaire d’environ 90 % qui est responsable de l’autre fonction de la sortiline : la régulation du trafic intracellulaire. En effet, l’extrémité cytoplasmique de la sortiline interagit avec le domaine VHS de la protéine de tri cytosolique GGA2 (Golgi-localizing, γ-adaptin ear homology domain, ADP-ribosylation factor-binding protein) [9]. Cette interaction permet en outre à la sortiline de contribuer à l’adressage des SAP (sphingolipid activator proteins) dans les lysosomes [10]. La sortiline, en tant que protéine « cargo », ferait partie intégrante d’une machinerie intracellulaire, située principalement au niveau du réseau transgolgien, qui servirait à l’adressage de multiples protéines vers les lysosomes et autres compartiments vésiculaires (Figure 2). La localisation de la sortiline dans des vésicules contenant des transporteurs de réserve comme le transporteur de glucose Glut4 [11] conforte cette fonction de tri intracellulaire.
Figure 2
Modèle d’un exemple de « cargo » d’adressage.
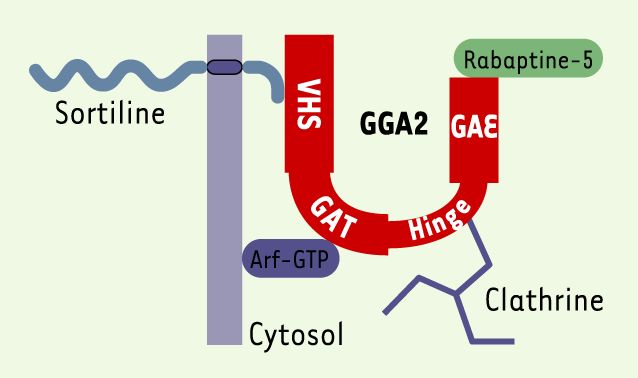
Le coeur de cette machinerie, située au niveau du TGN (trans-Golgi network), serait la protéine GGA2 (rouge) dont chaque domaine (VHS, GAT, Hinge et GAE) peut interagir avec différents partenaires. La sortiline reconnait le domaine VHS. L’ensemble de ces interactions faciliterait la formation des vésicules en réponse à l’échange de GTP sur Arf (ADP-ribosylation factor).
Le double rôle de la sortiline comme co-récepteur membranaire et comme protéine impliquée dans le tri cellulaire est donc en parfait accord avec sa double localisation. Tous les partenaires de la sortiline ont-ils été identifiés ? Rien n’est moins sûr. Par exemple, nous avons montré que la sortiline était impliquée dans les effets de la NT sur la croissance de cellules cancéreuses d’origine prostatique, pancréatique ou colonique [12]. Plusieurs de ces cellules n’exprimant pas le NTR1, le partenaire éventuel de la sortiline reste à identifier. La liste des fonctions de cette protéine à multiples facettes n’est donc probablement pas close.
Appendices
Références
- 1. Petersen CM, Nielsen MS, Nykjaer A, et al. Molecular identification of a novel candidate sorting receptor purified from human brain by receptor-associated protein affinity chromatography. J Biol Chem 1997 ; 272 : 3599-3605.
- 2. Mazella J, Zsürger N, Navarro V, et al. The 100-kDa neurotensin receptor is gp95/sortilin, a non-G-protein-coupled receptor. J Biol Chem 1998 ; 273 : 26273-6.
- 3. Nielsen MS, Jacobsen C, Olivecrona G, et al. Sortilin/neurotensin receptor-3 binds and mediates degradation of lipoprotein lipase. J Biol Chem 1999 ; 274 : 8832-6.
- 4. Munck Petersen C, Nielsen MS, Jacobsen C, et al. Propeptide cleavage conditions sortilin/neurotensin receptor-3 for ligand binding. EMBO J 1999 ;18 : 595-604.
- 5. Nykjaer A, Lee R, Teng KK, et al. Sortilin is essential for proNGF-induced neuronal cell death. Nature 2004 ; 427 : 843-8.
- 6. Dicou E, Lee J, Brachet P. Synthesis of nerve growth factor mRNA and precursor protein in the thyroid and parathyroid glands of the rat. Proc Natl Acad Sci 1986 ; 83 : 7084-8.
- 7. Martin S, Navarro V, Vincent JP, Mazella J. Neurotensin receptor-1 and -3 complex modulates the cellular signaling of neurotensin in the HT29 cell line. Gastroenterology 2002 ; 123 : 1135-43.
- 8. Martin S, Vincent JP, Mazella J. Involvement of the neurotensin receptor-3 in the neurotensin-induced migration of human microglia. J Neurosci 2003 ; 23 : 1198-1205.
- 9. Nielsen MS, Madsen P, Christensen EI, et al. The sortilin cytoplasmic tail conveys Golgi-endosome transport and binds the VHS domain of the GGA2 sorting protein. EMBO J 2001 ; 20 : 2180-90.
- 10. Lefrancois S, Zeng J, Hassan AJ, et al. The lysosomal trafficking of sphingolipid activator proteins (SAPs) is mediated by sortilin. EMBO J 2003 ; 22 : 6430-7.
- 11. Lin BZ, Pilch PF, Kandror KV. Sortilin is a major protein component of Glut4-containing vesicles. J Biol Chem 1997 ; 272 : 24145-7.
- 12. Dal Farra C, Sarret P, Navarro V, et al. Involvement of the neurotensin receptor subtype NTR3 in the growth effect of neurotensin on cancer cell lines. Int J Cancer 2001 ; 92 : 503-9.
List of figures
Figure 1
La sortiline, un co-récepteur associé à de multiples fonctions.
A. La sortiline est associée avec le NTR1 pour induire les effets de prolifération de la NT dans les cellules cancéreuses (HT29). B. A l’inverse, le complexe p75NTR/sortiline formé dans les neurones est responsable de la mort cellulaire provoquée par le proNGF. C. Dans les cellules microgliales humaines, la NT active la migration cellulaire par l’intermédiaire de la sortiline, probablement en association avec une protéine non identifiée capable de transduire le signal. D. Dans les adipocytes, la sortiline internalise la LpL (lipoprotéine lipase) et permet ainsi sa dégradation. Cette fonction ne nécessite pas forcément l’association avec un partenaire.
Figure 2
Modèle d’un exemple de « cargo » d’adressage.
Le coeur de cette machinerie, située au niveau du TGN (trans-Golgi network), serait la protéine GGA2 (rouge) dont chaque domaine (VHS, GAT, Hinge et GAE) peut interagir avec différents partenaires. La sortiline reconnait le domaine VHS. L’ensemble de ces interactions faciliterait la formation des vésicules en réponse à l’échange de GTP sur Arf (ADP-ribosylation factor).