Article body
De nombreux polluants semblent favoriser l’apparition de cancers et altérer les processus du développement et les fonctions reproductives mâles et femelles. Ces composés ont été appelés des perturbateurs endocriniens, et bien que leurs mécanismes d’action soient multiples, ils semblent agir principalement par liaison au récepteur de l’oestradiol et par la modification du métabolisme de cette hormone [1]. L’exemple type est celui de certains pesticides organochlorés dont beaucoup ont été progressivement interdits. Les dioxines peuvent être considérées comme des perturbateurs endocriniens mais leurs mécanismes d’action sont plus complexes. En effet, la plupart des travaux montrent qu’elles sont anti-oestrogéniques, et antagonisent la prolifération des cellules mammaires exposées aux hormones et l’expression de gènes oestrogéno-dépendants [2]. En revanche, d’autres travaux montrent des effets mimétiques des oestrogènes, en particulier sur les cellules utérines. Les dioxines semblent jouer un rôle dans la pathogénie des endométrioses chez la femme [3]; cette relation est suspectée principalement en raison de l’élévation de la quantité de ces polluants chez les femmes souffrant de cette maladie, et de l’apparition d’endométrioses chez des singes exposés à un traitement de plusieurs années par les dioxines. Les travaux sur l’interaction dioxine-oestradiol ont produit des résultats contradictoires probablement en raison de la complexité des effets hormonaux de la dioxine et de leur caractère paradoxal.
La dioxine agit principalement en interagissant avec le récepteur intracellulaire AHR (aryl hydrocarbon receptor) qui fait partie de la famille PAS-HLH (Per-Arnt-Sim homology-helix-loop-helix) des facteurs transcriptionnels. Son mécanisme d’action ressemble à celui des récepteurs nucléaires comme le récepteur de l’oestradiol bien qu’il n’y ait aucune homologie de structure entre ces deux familles. Plusieurs auteurs ont recherché une interaction entre le récepteur AHR et le récepteur de l’oestradiol ER pour expliquer les effets antagonistes des deux voies de signalisation. Bien que certains auteurs aient montré une telle interaction [4, 5], leurs résultats n’ont pu être confirmés par d’autres et la controverse a duré plusieurs années [6, 7]. Le récent article d’équipes japonaises et françaises montre de manière très convaincante l’interaction entre récepteur AHR et ERα ou ERβ et fournit des explications possibles aux résultats contradictoires passés [8, 9]. En effet, F. Ohtake et al. montrent que le 3-méthylcholanthrène, un agoniste du récepteur AHR est capable d’activer un promoteur contenant un élément de réponse aux oestrogènes uniquement en présence des deux récepteurs, AHR et ER (Figure1). Lorsque ce promoteur est activé par l’oestradiol, le 3-méthylcholanthrène a un effet antagoniste. Ces interactions fonctionnelles sont corrélées avec des liaisons physiques entre récepteur AHR et récepteurs ER démontrées par plusieurs techniques comme la co-immunoprécipitation. Cette interaction entre le domaine A/B des récepteurs ER et le domaine PAS du récepteur AHR dépend de la présence du ligand du AHR, mais elle est indépendante de la présence d’oestradiol. Par ailleurs, le partenaire du AHR, la protéine Arnt, dont la structure est semblable, n’interagit pas directement avec le ER. Le modèle proposé prévoit donc que le 3-méthylcholanthrène active le récepteur AHR qui s’associe à Arnt et recrute le récepteur ER en l’absence d’oestradiol pour activer les gènes oestrogéno-sensibles par l’intermédiaire du co-activateur p300. Ce modèle est conforté par des expériences in vivo chez la souris. En effet, les effets trophiques du 3-méthylcholanthrène sur l’utérus sont observés chez la souris ovariectomisée, mais sont absents chez les souris Ahr-/- ainsi que chez les souris Erα-/-.
Figure 1
Interaction entre les récepteurs ER et AHR.
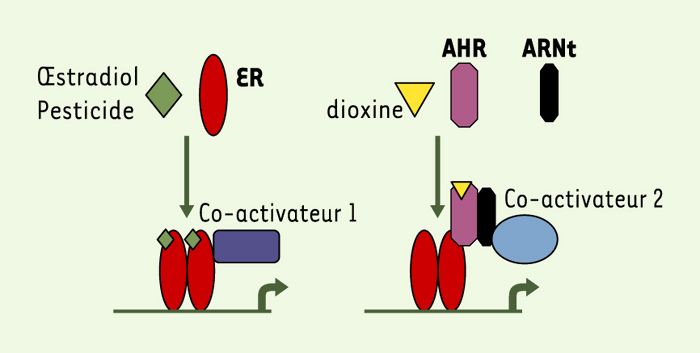
L’oestradiol ou des pesticides organochlorés se lient au récepteurs ER et activent la transcription par l’intermédiaire d’un ou plusieurs co-activateurs (ici dénommés 1). La dioxine se lie à son propre récepteur (AHR lui-même associé à Arnt) ; ce complexe est capable de s’associer aux récepteurs ER même en l’absence d’oestradiol et d’activer la transcription de gènes oestrogéno-dépendants par l’intermédiaire de co-activateurs (ici appelés 2). Il est possible que les co-activateurs 1 et 2 ne soient pas identiques.
Ces études suggèrent un mécanisme original d’activation du récepteur ER en l’absence d’oestradiol puisque le récepteur AHR, lorsqu’il est lié à ses ligands, est capable de s’associer au récepteur ER, en l’absence du ligand de ce dernier, et d’activer la transcription par son intermédiaire (Figure1). Ce mécanisme est différent de celui d’autres perturbateurs endocriniens qui se lient directement au récepteur de l’oestradiol. En présence d’oestradiol, les auteurs montrent bien le rôle antagoniste des ligands du AHR. On peut penser que le complexe AHR-ER altère la sensibilité du ER pour son ligand naturel, soit en diminuant son affinité soit à une étape ultérieure, par exemple en favorisant la liaison de co-activateurs différents. Ces mécanismes restent cependant à élucider. L’apport de ce travail par rapport aux travaux précédents est de montrer la liaison des récepteurs AHR et ER en l’absence d’hormone et de mettre en évidence le comportement différent des ligands du AHR en l’absence ou en présence d’oestradiol. Fonctionnellement, ils ont un comportement d’agonistes partiels, d’où leurs effets variables selon les systèmes expérimentaux étudiés. Ce mode d’action complexe explique probablement les effets globaux relativement subtils des dioxines et des autres agonistes du récepteur AHR.
Appendices
Références
- 1. Massaad C, Entezami F, Massade L, et al. How can chemical compounds alter human fertility? Eur J Obstet Gynecol Reprod Biol 2002 ; 100 : 127-37.
- 2. Krishnan V, Porter W, Santostefano M, Wang X, Safe S. Molecular mechanism of inhibition of estrogen-induced cathepsin D gene expression by 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin (TCDD) in MCF-7 cells. Mol Cell Biol 1995 ; 15 : 6710-9.
- 3. Gibbons A. Dioxin tied to endometriosis. Science 1993 ; 262 : 1373.
- 4. Kharat I, Saatcioglu F. Antiestrogenic effects of 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin are mediated by direct transcriptional interference with the liganded estrogen receptor. Cross-talk between aryl hydrocarbon- and estrogen-mediated signaling. J Biol Chem 1996; 271: 10533-7.
- 5. Klinge CM, Kaur K, Swanson HI. The aryl hydrocarbon receptor interacts with estrogen receptor alpha and orphan receptors COUP-TFI and ERRalpha1. Arch Biochem Biophys 2000 ; 373 : 163-74.
- 6. Duan R, Porter W, Samudio I, Vyhlidal C, Kladde M, Safe S. Transcriptional activation of c-fos protooncogene by 17beta-estradiol: mechanism of aryl hydrocarbon receptor-mediated inhibition. Mol Endocrinol 1999; 13 : 1511-21.
- 7. Rogers JM, Denison MS. Analysis of the antiestrogenic activity of 2, 3, 7, 8-tetrachlorodibenzo-p-dioxin in human ovarian carcinoma BG-1 cells. Mol Pharmacol 2002 ; 61: 1393-403.
- 8. Ohtake F, Takeyama K, Matsumoto T, et al. Modulation of estrogen receptor signalling by association with the activated dioxin receptor. Nature 2003 ; 423 : 545-50.
- 9. Brosens JJ, Parker MG. Gene expression: estrogen receptor hijacked. Nature 2003 ; 423 : 487-8.
List of figures
Figure 1
Interaction entre les récepteurs ER et AHR.
L’oestradiol ou des pesticides organochlorés se lient au récepteurs ER et activent la transcription par l’intermédiaire d’un ou plusieurs co-activateurs (ici dénommés 1). La dioxine se lie à son propre récepteur (AHR lui-même associé à Arnt) ; ce complexe est capable de s’associer aux récepteurs ER même en l’absence d’oestradiol et d’activer la transcription de gènes oestrogéno-dépendants par l’intermédiaire de co-activateurs (ici appelés 2). Il est possible que les co-activateurs 1 et 2 ne soient pas identiques.