Abstracts
Résumé
Dans le noyau de la cellule, le nucléole est le centre de synthèse des ribosomes. Domaine nucléaire dynamique, son activité reflète un équilibre entre le niveau de synthèse des ARN ribosomiques (ARNr), directement lié à la croissance et à la prolifération cellulaires, l’efficacité de la maturation des ARNr, et le transport des sous-unités ribosomiques vers le cytoplasme. Le nucléole disparaît avant la division cellulaire et réapparaît juste après, l’assemblage du nucléole étant un événement très précoce en sortie de mitose. La fabrication des ribosomes est interrompue pendant la mitose, mais les machineries nucléolaires sont transmises aux cellules filles. Le nucléole est également un domaine nucléaire multifonctionnel qui joue un rôle important dans l’organisation nucléaire. Des mutations au niveau de gènes codant pour des protéines nucléolaires sont associées à des maladies humaines.
Summary
In eukaryotes, the nucleolus is the ribosome factory. The nucleolus is a very active large nuclear domain resulting from the equilibrium between level of ribosomal gene transcription, efficiency of rRNA processing and transport of the ribosomal subunits (40S and 60S) towards the cytoplasm. The ribosome production is regulated and is linked with cell growth and cell proliferation. The ribosome production is stopped during mitosis but the nucleolar machineries are inherited in daughter cells and the nucleolar reassembly is a very early event at the exit of mitosis. The nucleolus is also a multifunctional domain involved in nuclear architecture and specific interaction with some nuclear bodies. Finally, several human diseases appear to result from mutations of nucleolar proteins.
Article body
Le nucléole est une structure nucléaire proéminente, visible en microscopie conventionnelle par contraste de phase (Figure 1A), ce qui lui a valu son nom d’organite bien qu’il ne soit entouré d’aucune membrane. Connu depuis les années 1960 comme étant le centre de synthèse des ARN ribosomiques (ARNr) et d’assemblage des sous-unités ribosomiques, le nucléole contient les gènes codant pour les ARNr ainsi que des particules contenant les ARNr 18S et 28S à différents stades de maturation et d’assemblage avec les protéines ribosomiques [1]. Depuis, le nucléole a fait l’objet de nombreuses recherches visant à comprendre la biogenèse des ribosomes [2, 3] et les relations entre la fonction nucléolaire et la croissance cellulaire [4]. Plus récemment, le nucléole a été impliqué dans d’autres fonctions cellulaires, comme l’assemblage ou la maturation de complexes impliquant des ARN différents des ARNr (signal recognition particle ou SRP, ARN de transfert, ARN U6, complexe télomérase), le cycle cellulaire ou le vieillissement [5, 6]. Les données de la protéomique nucléolaire mettent en évidence une complexité inconnue jusqu’ici [7] et indiquent que le nucléole est au centre d’échanges entre différents domaines nucléaires [8, 9], dont on commence à découvrir l’organisation [10, 11]. Les principes qui régissent l’expression des gènes ribosomiques étant généralisables à d’autres types de gènes, l’étude du nucléole permet une meilleure compréhension de l’organisation fonctionnelle du noyau.
Figure 1
Structure du nucléole.

A. Cellules humaines en culture (HeLa) observées en contraste de phase. Les nucléoles sont les masses très contrastées visibles dans les noyaux. B. Organisation du nucléole observée en microscopie électronique à transmission et B’ représentation schématique de cette image. Sur cette coupe fine, on distingue plusieurs centres fibrillaires (CF, jaune) entourés de composant fibrillaire dense (CFD, vert). Ces structure sont enchâssées dans le composant granulaire (CG, rose). Le contact entre nucléole et enveloppe nucléaire formant une invagination est visible sur la partie inférieure de l’image (blanc). La chromatine condensée est figurée en gris moucheté. échelle: A = 10 µm, B = 0,5 µm.
Production de ribosomes par le nucléole
Le nucléole est un domaine très dynamique dont l’apparence varie en fonction de son activité [12]. Chez la grande majorité des eucaryotes y compris la levure, quand la biogenèse des ribosomes est active, le nucléole est organisé en trois composants majeurs visibles en microscopie électronique: les centres fibrillaires sont des régions claires dont la taille varie de 0,1 à 1 µm; ils sont entourés par le composant fibrillaire dense, dont le nom est directement lié à son aspect contrasté et à sa texture; ces deux composants sont enchâssés dans le composant granulaire constitué de granules de 15 à 20 nm (Figure 1B, 1B’) [2, 3, 13].
Les ribosomes sont constitués de deux sous-unités, une petite de 40S et une grande de 60S qui sont produites dans le nucléole et transportées séparément dans le cytoplasme (Figure 2). La sous-unité de 40S est constituée de l’ARNr 18S auquel sont associées 33 protéines ribosomiques. La sous-unité de 60S est formée par les ARNr 28S, 5,8S et 5S auxquels sont associées 49 protéines ribosomiques. Les ARNr 28S, 18S et 5,8S sont synthétisés, maturés et assemblés avec les protéines ribosomiques dans le nucléole à partir de transcrits primaires (ARNr 47S chez les mammifères) issus de la transcription des gènes ribosomiques par l’ARN polymérase I (Pol I). Les étapes les plus tardives sont réalisées au cours du transport des sous-unités ribosomiques dans le nucléoplasme ou même dans le cytoplasme. L’ARNr 5S est associé à la sous-unité ribosomique 60S dans le nucléole après avoir été synthétisé dans le nucléoplasme par l’ARN polymérase III (Figure 2).
Figure 2
Biogenèse des ribosomes.
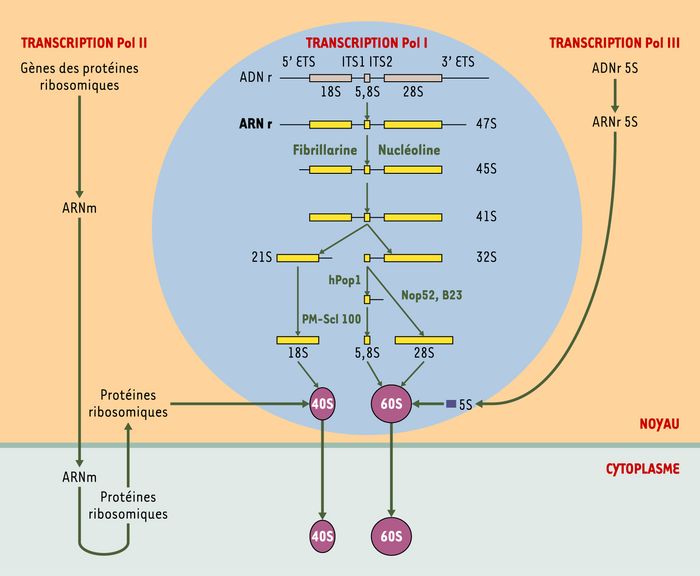
Le nucléole (cercle bleu) est le site de synthèse et de maturation des ARN ribosomiques (ARNr) 28S, 18S et 5,8S et le site d’association avec l’ARNr 5S. C’est également le lieu où les ARNr s’associent avec les protéines ribosomiques. La synthèse des ARNr (rectangles jaunes) est suivie de leur maturation par élimination, par étapes successives, des séquence ETS (external transcribed sequence) et ITS (internal transcribed sequence) (traits fins noirs) par des complexes agissant séquentiellement. Il faut noter que la fabrication des ribosomes nécessite la mise en oeuvre des trois machineries de transcription. Les gènes (ADN) des ARNr 18S, 5,8S et 28S sont transcrits par l’ARN polymérase I (Pol I), l’ARNr 5S par l’ARN polymérase III (Pol III) et ceux codant pour les protéines ribosomiques par l’ARN polymérase II (Pol II). Certaines des protéines impliquées dans la maturation sont indiquées en lettres vertes. hPop1: processing of precursor RNAs; Nop52: nucleolar protein 52; B23: protéine nucléolaire; PM-Scl 100: 100kDa-polymyositis/scleroderma antigen.
La quantité de ribosomes produits dépend de l’efficacité de ces machineries nucléolaires mais aussi du nombre de gènes ribosomiques activés.
Gènes ribosomiques
Ce sont des gènes répétés, regroupés dans des régions chromosomiques appelées régions organisatrices du nucléole ou NOR (nucleolar organizer region). Le nucléole s’organise autour du NOR et, dans certaines espèces comme chez l’homme, il y a coopération entre plusieurs NOR pour former un nucléole. Le nombre de gènes ribosomiques varie de 100 à plusieurs centaines chez les mammifères. Chez l’homme, les 400 copies sont regroupées sur les chromosomes 13, 14, 15, 21 et 22. Cependant, toutes les copies ne sont pas actives: même lorsqu’une forte production de ribosomes est nécessaire, on estime que seulement 60% des gènes ribosomiques sont actifs dans les cellules de mammifères en phase exponentielle de croissance [14] et 46% chez la levure [15].
L’organisation en tandem des gènes ribosomiques (séquences répétées orientées dans le même sens) est retrouvée chez tous les eucaryotes et, chez la levure, semble contrôler l’activité et l’organisation nucléolaires [13, 16]. Certaines copies de gènes ribosomiques pourraient être sous forme hétérochromatique, favorable au regroupement des gènes ribosomiques et à l’intégrité du nucléole. Il faut noter que les gènes actifs et inactifs sont intercalés chez la levure [17] comme dans les cellules de mammifères [18]. Pendant la mitose, la chromatine des NOR présente deux configurations, vraisemblablement en rapport avec des états différents des gènes ribosomiques dans un même NOR [19].
Transcription des gènes ribosomiques
L’initiation de la transcription des gènes ribosomiques se situe à la jonction centres fibrillaires-composant fibrillaire dense (Figure 1B’). Elle dépend de Pol I et de facteurs spécifiques des gènes ribosomiques (Figure 3) [4, 20]. Récemment, il a été montré que le facteur TFIIH (transcription factor II H), impliqué dans l’initiation de la transcription par Pol II, est essentiel à la transcription par la Pol I [21]. TFIIH, impliqué également dans la réparation de la chromatine, pourrait participer au couplage entre transcription et réparation des gènes ribosomiques.
Figure 3
La transcription de l’ADN ribosomique.

1. L’ADN ribosomique (ADNr) est associé aux facteurs de transcription UBF (upstream binding factor) (vert) et SL1 (selectivity factor 1) (jaune) dans la région du promoteur. 2. L’ARN polymérase I (PolI) est recrutée sur le site d’initiation de la transcription (flèche rouge) par l’intermédiaire du facteur RRN3 (bleu) [51]. 3. Au cours de l’élongation, Pol I se déplace (sens de la flèche verte) jusqu’au site de terminaison associé au facteur de terminaison TTF1 (transcription termination factor 1) (rose), qui interagit également avec le promoteur [52]. Le point d’interrogation indique que la question de la participation de la dimérisation de TTF1 à l’organisation en boucle des gènes ribosomiques reste à élucider.
Maturation des ARN ribosomiques
La maturation des ARNr est déclenchée dans le composant fibrillaire dense et se poursuit pendant la migration des ARNr vers le composant granulaire (Figure 1). Elle est très complexe puisqu’elle implique plus d’une centaine de protéines et au moins autant de petits ARN nucléolaires (small nucleolar RNA ou snoARN en français). La maturation des ARNr nécessite deux étapes cruciales: (1) la modification des ARNr par plusieurs dizaines de méthylations et pseudo-uridylations; (2) le clivage des ARNr primaires à différents sites [22, 23]. Le schéma général de ce processus complexe est très similaire de la levure à l’homme. En particulier, l’ordre des clivages et le rôle de guide des snoARN responsables des modifications des ARNr sont conservés au cours de l’évolution [23]. Différents complexes interviennent de façon ordonnée pour éliminer les régions transcrites externes (5’ et 3’) et internes de l’ARNr primaire, qui ne seront pas conservées dans les ARNr matures (Figure 2). Les clivages qui vont conduire à l’élimination des régions transcrites internes font intervenir des exonucléases [24], des hélicases (huit hélicases impliquées dans la maturation précoce et neuf dans la maturation tardive chez E. coli) [25] et différentes protéines dont le rôle exact n’est pas connu. Les étapes de maturation qui donnent naissance à la grande sous-unité de 60S étant plus longues et intervenant après les premiers clivages, les protéines impliquées dans ces étapes ont été qualifiées de machinerie de maturation tardive [26]. La caractérisation des intermédiaires d’assemblage de la sous-unité de 60S a montré, par une approche protéomique, que de nombreuses protéines interviennent successivement sur un ARNr au même stade de maturation, soit à des étapes précises, soit en restant associées pendant plusieurs étapes [27].
Continuité entre les étapes
Il est essentiel pour la cellule de maintenir une continuité entre les différentes étapes du processus de synthèse des ARNr, notamment entre transcription et maturation. Bien qu’il y ait une association co-transcriptionnelle des ARNr avec certains complexes de maturation précoce, c’est seulement après la terminaison de la transcription que le clivage de l’ARN 47S commence. Chez la levure, le clivage du transcrit primaire (ARNr 35S) commence 10 secondes après la fin de la transcription [28] et la clôture de la transcription pourrait être un facteur de contrôle de qualité pendant la biogenèse des ribosomes. Cependant, la recherche d’un signal permettant d’associer transcription et maturation n’a donné aucun résultat. Récemment, nous avons montré qu’il est possible de déconnecter la fin de la transcription de l’ADNr de la maturation des ARN 47S grâce à des inhibiteurs de kinases [29]. On peut donc supposer qu’il s’agit d’un processus contrôlé sans que l’on sache actuellement par quel mécanisme.
Une perturbation ou un blocage de la transcription de l’ARNr induit une ségrégation des composants du nucléole [30, 31] caractérisée par un regroupement des partenaires associés à une étape et à leur juxtaposition. Dans ce cas, les protéines de la machinerie de transcription forment un croissant caractéristique situé au-dessus des deux masses formées respectivement par les machineries de maturation précoce et tardive. Cette image typique, observable en microscopie optique, indique une inhibition de transcription des gènes ribosomiques [32]. Le prototype de la ségrégation nucléolaire est obtenu par traitement des cellules par l’actinomycine D, qui, à faible dose, induit un blocage spécifique de la transcription par Pol I. Si la maturation de l’ARNr est perturbée alors que la transcription est maintenue, il y a séparation des machineries de maturation et de transcription. Lorsque cette perturbation est induite par des inhibiteurs de kinases [29, 33], les gènes ribosomiques se déploient dans le nucléoplasme et les sites de transcription apparaissent comme un collier de perles [34]. Dans ce cas, il n’y a plus de nucléole caractéristique mais une structure couramment appelée nucléole en collier de perles, visible dans le nucléoplasme par contraste de phase [33].
Cycle cellulaire et nucléole
Chez les eucaryotes supérieurs, la mitose correspond à une période d’inactivation et/ou de délocalisation des machineries nucléaires, et l’organisation fonctionnelle du noyau doit être reconstituée après chaque mitose. Le nucléole, assemblé en fin de mitose (Figure 4), est actif pendant l’interphase et désassemblé en début de mitose. Dans le cas des mitoses sans rupture de l’enveloppe nucléaire, comme chez la levure S. cerevisiae, l’activité nucléaire persiste et le nucléole n’est pas désorganisé pendant la mitose.
Figure 4
Nucléole et cycle cellulaire.
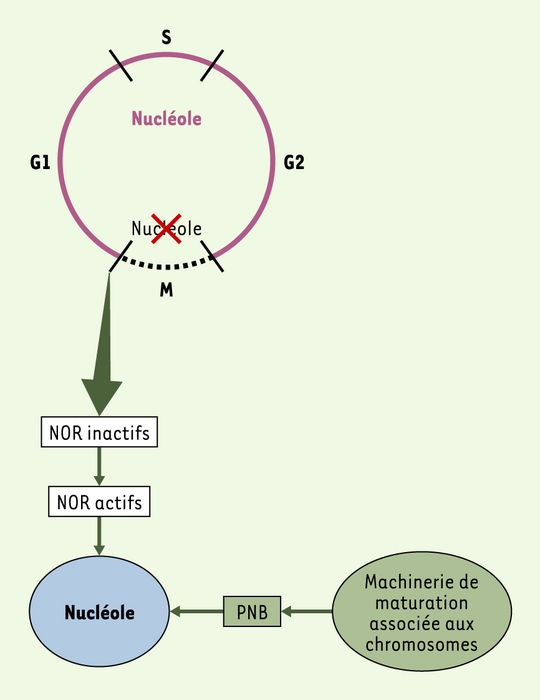
Le nucléole est présent pendant toute l’interphase (G1 à G2) (cercle rose) et est absent pendant la mitose (pointillé noir). Il se dissocie à la transition G2/M quand la machinerie de maturation se disperse à la périphérie des chromosomes et quand, de façon concomitante, la transcription l’ARN polymérase I (Pol I) est réprimée. Cependant, la machinerie de transcription Pol I reste assemblée sur les NOR (nucleolar organizer region). Pour réassembler un nouveau nucléole en sortie de mitose, il faut activer la transcription Pol I (NOR actifs) et recruter la machinerie de maturation sur les sites de transcription. Ce recrutement requiert la formation de corps nucléaires appelés PNB (pre-nucleolar bodies).
L’assemblage du nucléole est réalisé très précocement en sortie de mitose à partir des machineries ou des complexes transmis de la cellule mère aux cellules filles. Dans les cellules humaines, la transcription par Pol I commence en télophase et le recrutement de la machinerie de maturation dure en moyenne 90 minutes, puis plusieurs NOR s’associent pour former un nucléole.
Les facteurs UBF (upstream binding factor), SL1 (selectivity factor 1), TTF1 (transcription termination factor I) et le complexe Pol I restent associés aux gènes ribosomiques pendant la mitose [19, 35, 36], mais sous une forme inactive. C’est la kinase CDK1-cycline B qui est responsable de cette inactivation, l’inhibition de cette kinase étant capable de restaurer la transcription par Pol I dans des cellules mitotiques [37]. Ainsi, en sortie de mitose, quand la dégradation de la cycline B supprime la répression mitotique de la machinerie Pol I, la transcription des ARNr reprend (Figure 4).
Pendant la mitose, les machineries de maturation des ARNr sont distribuées autour des chromosomes. En sortie de mitose, la concentration de ces machineries à la surface des chromosomes crée des masses morphologiquement identifiables, connues sous le nom de PNB (pre-nucleolar bodies), qui pourraient être des plates-formes d’assemblage des machineries de maturation. Les protéines des PNB sont relocalisées sur les sites de transcription par tris successifs, comme le montre l’analyse en temps réel dans des cellules vivantes qui a permis de visualiser des transmissions directionnelles entre PNB et sites de transcription [26]. La formation des PNB en sortie de mitose dépend de l’inactivation de CDK1, comme le ciblage des machineries de maturation précoces, tandis que le ciblage des machineries de maturation tardives est contrôlé par une autre kinase encore non identifiée [29].
Comment sont recrutées les machineries de maturation des ARNr au moment de l’assemblage d’un nucléole en fin de mitose? La cinétique observée suggère que ce recrutement dépendrait de l’activation de la transcription [3]. Pourtant, l’élément déterminant semble être la présence d’ARNr et non la transcription. En effet, même lorsque la transcription par Pol I est bloquée dans les cellules en sortie de mitose, on observe le recrutement, autour des gènes ribosomiques, des protéines de la machinerie de maturation associées à des pré-ARNr stables transmis pendant la mitose [32]. De même, au cours du développement de l’embryon de xénope, la présence d’ARNr provenant du stock maternel est capable de recruter la machinerie de maturation autour des gènes ribosomiques en l’absence de transcription [38, 39]. L’ARNr semble donc jouer un rôle crucial dans la structuration du nucléole ((→) m/s 2003, n°6-7, p.673).
L’activité du nucléole est d’autant plus importante que la prolifération cellulaire est rapide, comme par exemple dans les cellules cancéreuses, et elle est fortement réprimée, voire arrêtée dans les cellules différenciées [4, 20]. L’efficacité de la machinerie de transcription peut être contrôlée au niveau de la formation du complexe de préinitiation, de l’initiation de la transcription par Pol I via l’expression de facteurs de transcription spécifiques [40, 41] ou au niveau de l’activité des gènes ribosomiques. Le contrôle de l’activité de transcription par Pol I en fonction du cycle cellulaire peut dépendre de la protéine du rétinoblastome, un suppresseur de tumeur qui joue un rôle crucial dans le contrôle du cycle cellulaire, et qui peut inhiber l’activité de transcription Pol I en agissant directement sur UBF [42]. Par ailleurs, une maturation anormale des ARNr induit l’accumulation du suppresseur de tumeur p53 et par conséquent l’arrêt du cycle cellulaire [43, 44], ce qui constitue un autre lien entre cycle cellulaire et biogenèse des ribosomes.
Enfin, des mutations au niveau de gènes codant pour des protéines du nucléole sont associées à des maladies humaines. Dans certains cas, les symptômes de la maladie pourraient être la conséquence directe de mutations qui affectent la production de ribosomes (voir Encadré).
Conclusions
Le nucléole est un domaine nucléaire résultant de l’activité de production de ribosomes. L’équilibre entre transcription, maturation et transport des sous-unités ribosomiques crée un domaine fonctionnel de grande taille qui constitue une zone de séquestration ou d’exclusion de molécules participant à des fonctions différentes de la biogenèse des ribosomes. Le nucléole représente la plus forte concentration d’ARN dans la cellule, associé à des machineries sophistiquées impliquant un grand nombre de snoARN. Cette concentration localisée semble permettre l’assemblage de ribonucléoprotéines différentes des ribosomes. En outre, le nucléole établit de nombreux contacts et échanges avec plusieurs corps nucléaires. La compréhension des fonctions nucléolaires intégrées dans le contexte nucléaire devrait permettre de répondre à plusieurs questions non résolues concernant, notamment, le rôle des protéines ribosomiques dans la dynamique d’assemblage des nucléoles, le contrôle du transport des sous-unités ribosomiques et l’existence d’un contrôle de qualité des ribosomes produits.
Appendices
Remerciements
Les auteurs remercient N. Angelier, C. Carles, A.L. Haenni et P.Roussel pour les suggestions concernant le manuscrit ainsi que Sophie Le Panse pour le cliché de microscopie électronique. Ce travail a bénéficié des subventions du Centre National de la Recherche Scientifique (UMR 7592) et de l’Association pour la Recherche sur le Cancer (contrat n°4290).
Références
- 1. Hadjiolov AA. The nucleolus and ribosome biogenesis. Wien-New York: Springer-Verlag, 1985: 268 p.
- 2. Shaw PJ, Jordan EG. The nucleolus. Annu Rev Cell Dev Biol 1995; 11: 93-121.
- 3. Scheer U, Hock R. Structure and function of the nucleolus. Curr Opin Cell Biol 1999; 11: 385-90.
- 4. Comai L. The nucleolus: a paradigm for cell proliferation and aging. Braz JMed Biol Res 1999; 32: 1473-8.
- 5. Pederson T. The plurifunctional nucleolus. Nucleic Acids Res 1998; 26: 3871-6.
- 6. Visintin R, Amon A. The nucleolus: the magician’s hat for cell cycle tricks. Curr Opin Cell Biol 2000; 12: 372-7.
- 7. Andersen JS, Lyon CE, Fox AH, et al. Directed proteomic analysis of the human nucleolus. Curr Biol 2002; 12: 1-11.
- 8. Leung AK, Lamond AI. In vivo analysis of NHPX reveals a novel nucleolar localization pathway involving a transient accumulation in splicing speckles. J Cell Biol 2002; 157: 615-29.
- 9. Fox AH, Lam YW, Leung AKL, et al. Paraspeckles: a novel nuclear domain. Curr Biol 2002; 12: 13-25.
- 10. Cremer T, Cremer C. Chromosome territories, nuclear architecture and gene regulation in mammalian cells. Nat Rev Genet 2001; 2: 292-301.
- 11. Strouboulis J, Wolffe AP. Functional compartmentalization of the nucleus. J Cell Sci 1996; 109: 1991-2000.
- 12. Mélèse T, Xue Z. The nucleolus: an organelle formed by the act of building a ribosome. Curr Opin Cell Biol 1995; 7: 319-24.
- 13. Trumtel S, Léger-Silvestre I, Gleizes PE, Teulières F, Gas N. Assembly and functional organization of the nucleolus: ultrastructural analysis of Saccharomyces cerevisiae mutants. Mol Biol Cell 2000; 11: 2175-89.
- 14. Conconi A, Widmer RM, Koller T, Sogo JM. Two different chromatin structures coexist in ribosomal RNA genes throughout the cell cycle. Cell 1989; 57: 753-61.
- 15. Dammann R, Lucchini R, Koller T, Sogo JM. Chromatin structures and transcription of rDNA in yeast Saccharomyces cerevisiae. Nucleic Acids Res 1993; 21: 2331-8.
- 16. Shou W, Sakamoto KM, Keener J, et al. Net1 stimulates RNA polymerase I transcription and regulates nucleolar structure independently of controlling mitotic exit. Mol Cell 2001; 8: 45-55.
- 17. Dammann R, Lucchini R, Koller T, Sogo JM. Transcription in the yeast rRNA gene locus: distribution of the active gene copies and chromatin structure of their flanking regulatory sequences. Mol Cell Biol 1995; 15: 5294-303.
- 18. Junéra HR, Masson C, Géraud G, Suja J, Hernandez-Verdun D. Involvement of in situ conformation of ribosomal genes and selective distribution of UBF in rRNA transcription. Mol Biol Cell 1997; 8: 145-56.
- 19. Gébrane-Younès J, Fomproix N, Hernandez-Verdun D. When rDNA transcription is arrested during mitosis, UBF is still associated with non-condensed rDNA. J Cell Sci 1997; 110: 2429-40.
- 20. Grummt I. Regulation of mammalian ribosomal gene transcription by RNA polymerase I. Prog Nucleic Acids Res Mol Biol 1999; 62: 109-54.
- 21. Iben S, Tschochner H, Bier M, et al. TFIIH plays an essential role in RNA polymerase I transcription. Cell 2002; 109: 297-306.
- 22. Venema J, Tollervey D. Ribosome synthesis in Saccharomyces cerevisiae. Annu Rev Genet 1999; 33: 261-311.
- 23. Filipowicz W, Pogacic V. Biogenesis of small nucleolar robonucleoproteins. Curr Opin Cell Biol 2002; 14: 319-27.
- 24. Mitchell P, Petfalski E, Tollervey D. The 3’ end of the yeast 5.8 rRNA is generated by an exonuclease processing mechanism. Genes Dev 1996; 10: 502-13.
- 25. Tanner K, Linder P. DExD/H box RNA helicases: from generic motors to specific dissociation functions. Mol Cell 2001; 8: 251-62.
- 26. Savino TM, Gébrane-Younès J, De Mey J, Sibarita JB, Hernandez-Verdun D. Nucleolar assembly of the rRNA processing machinery in living cells. J Cell Biol 2001; 153: 1097-110.
- 27. Harnpicharnchai P, Jakovljevic J, Horsey E, et al. Composition and functional characterization of yeast 66S ribosome assembly intermediates. Mol Cell 2001; 8: 505-15.
- 28. Allmang C, Tollervey D. The role of the 3’ external transcribed spacer in yeast pre-rRNA processing. J Mol Biol 1998; 278: 67-78.
- 29. Sirri V, Hernandez-Verdun D, Roussel P. Cyclin-dependent kinases govern formation and maintenance of the nucleolus. J Cell Biol 2002; 156: 969-81.
- 30. Puvion-Dutilleul F, Mazan S, Nicoloso M, Pichard E, Bachellerie JP, Puvion E. Alterations of nucleolar ultrastructure and ribosome biogenesis by actinomycin D. Implications for U3 snRNP function. Eur J Cell Biol 1992; 58: 149-62.
- 31. Puvion-Dutilleul F, Puvion E, Bachellerie JP. Early stages of pre-rRNA formation within the nucleolar ultrastructure of mouse cells studied by in situ hybridization with 5’ETS leader probe. Chromosoma 1997; 105: 496-505.
- 32. Dousset T, Wang C, Verheggen C, Chen D, Hernandez-Verdun D, Huang S. Initiation of nucleolar assembly is independent of RNA polmerase I transcription. Mol Biol Cell 2000; 11: 2705-17.
- 33. Le Panse S, Masson C, Héliot L, Chassery JM, Junéra HR, Hernandez-Verdun D. 3-D organization of single ribosomal transcription units after DRB inhibition of RNA polymerase II transcription. J Cell Sci 1999; 112: 2145-54.
- 34. Haaf T, Ward DC. Inhibition of RNA polymerase II transcription causes chromatin decondensation, loss of nucleolar structure, and dispersion of chromosomal domains. Exp Cell Res 1996; 224: 163-73.
- 35. Roussel P, André C, Comai L, Hernandez-Verdun D. The rDNA transcription machinery is assembled during mitosis in active NORs and absent in inactive NORs. J Cell Biol 1996; 133: 235-46.
- 36. Sirri V, Roussel P, Hernandez-Verdun D. The mitotically phosphorylated form of the transcription termination factor TTF-1 is associated with the repressed rDNA transcription machinery. J Cell Sci 1999; 112: 3259-68.
- 37. Sirri V, Roussel P, Hernandez-Verdun D. In vivo release of mitotic silencing of ribosomal gene transcription does not give rise to precursor ribosomal RNA processing. J Cell Biol 2000; 148: 259-70.
- 38. Verheggen C, Le Panse S, Almouzni G, Hernandez-Verdun D. Presence of pre-rRNAs before activation of polymerase I transcription in the building process of nucleoli during early development of Xenopus laevis. J Cell Biol 1998; 142: 1167-80.
- 39. Verheggen C, Almouzni G, Hernandez-Verdun D. The ribosomal RNA processing machinery is recruited to the nucleolar domain before RNA polymerase I during Xenopus laevis development. J Cell Biol 2000; 149: 293-305.
- 40. Voit R, Hoffmann M, Grummt I. Phosphorylation by G1-specific cdk-cyclin complexes activates the nucleolar transcription factor UBF. EMBO J 1999; 18: 1891-9.
- 41. Panov KI, Friedrich JK, Zomerdijk JCBM. A step subsequent to preinitiation complex assembly at the ribosomal RNA gene promoter is rate limiting for human RNA polymerase I-dependent transcription. Mol Cell Biol 2001; 21: 2641-9.
- 42. Cavanaugh AH, Hempel WM, Taylor LJ, Rogalsky V, Todorov G, Rothblum LI. Activity of RNA polymerase I transcription factor UBF blocked by rb gene product. Nature 1995; 374: 177-80.
- 43. David-Pfeuty T, Nouvian-Dooghe Y, Sirri V, Roussel P, Hernandez-Verdun D. Common and reversible regulation of wild-type p53 function and of ribosomal biogenesis by protein kinases in human cells. Oncogene 2001; 20: 5951-63.
- 44. Pestov DG, Strezoska Z, Lau LF. Evidence of p53-dependent cross-talk between ribosome biogenesis and cell cycle: effects of nucleolar protein Bop1 on G1/S transition. Mol Cell Biol 2001; 21: 4246-55.
- 45. Marciniak RA, Lombard DB, Johnson FB, Guarente L. Nucleolar localization of the Werner syndrome protein in human cells. Proc Natl Acad Sci USA 1998; 95: 6887-92.
- 46. Sinclair DA, Mills K, Guarente I. Accelerated aging and nucleolar fragmentation in yeast sgs1 mutants. Science 1997; 277: 1313-6.
- 47. Shiratori M, Suzuki T, Itoh C, Goto M, Furuichi Y, Matsumoto T. WRN helicase accelarates the transcription of ribosomal RNA as a component of an RNA polymerase I-associated complex. Oncogene 2002; 21: 2447-54.
- 48. Dixon J, Edwards SJ, Anderson I, Brass A, Scambler PJ, Dixon MJ. Identification of the complete coding requence and genomic organization of the treacher Collins syndrome gene. Genome res 1997; 7: 223-34.
- 49. Isaac C, Marsh KL, Paznekas WA, et al. Characterization of the nucleolar gene product, treacle, in Treacher Collins syndrome. Mol Biol Cell 2000; 11: 3061-71.
- 50. Dez C, Henras A, Faucon B, Lafontaine D, Caizergues-Ferrer M, Henry Y. Stable expression in yeast of the mature form of human telomerase RNA depends on its association with the box H/ACA small nucleolar RNP proteins Cbf5p, Nhp2p and Nop10p. Nucleic acids Res 2001; 29: 598-603.
- 51. Miller G, Panov KI, Friedrich JK, Trinkle-Mulcahy L, Lamond AI, Zomerdijk JC. hRRN3 is essential in the SL1-mediated recruitment of RNA polymerase I to rRNA gene promoters. EMBO J 2001; 20: 1373-82.
- 52. Längst G, Becker PB, Grummt I. TTF-1 determines the chromatin architecture of the active rDNA promoter. EMBO J 1998; 17: 3135-43.
List of figures
Figure 1
Structure du nucléole.
A. Cellules humaines en culture (HeLa) observées en contraste de phase. Les nucléoles sont les masses très contrastées visibles dans les noyaux. B. Organisation du nucléole observée en microscopie électronique à transmission et B’ représentation schématique de cette image. Sur cette coupe fine, on distingue plusieurs centres fibrillaires (CF, jaune) entourés de composant fibrillaire dense (CFD, vert). Ces structure sont enchâssées dans le composant granulaire (CG, rose). Le contact entre nucléole et enveloppe nucléaire formant une invagination est visible sur la partie inférieure de l’image (blanc). La chromatine condensée est figurée en gris moucheté. échelle: A = 10 µm, B = 0,5 µm.
Figure 2
Biogenèse des ribosomes.
Le nucléole (cercle bleu) est le site de synthèse et de maturation des ARN ribosomiques (ARNr) 28S, 18S et 5,8S et le site d’association avec l’ARNr 5S. C’est également le lieu où les ARNr s’associent avec les protéines ribosomiques. La synthèse des ARNr (rectangles jaunes) est suivie de leur maturation par élimination, par étapes successives, des séquence ETS (external transcribed sequence) et ITS (internal transcribed sequence) (traits fins noirs) par des complexes agissant séquentiellement. Il faut noter que la fabrication des ribosomes nécessite la mise en oeuvre des trois machineries de transcription. Les gènes (ADN) des ARNr 18S, 5,8S et 28S sont transcrits par l’ARN polymérase I (Pol I), l’ARNr 5S par l’ARN polymérase III (Pol III) et ceux codant pour les protéines ribosomiques par l’ARN polymérase II (Pol II). Certaines des protéines impliquées dans la maturation sont indiquées en lettres vertes. hPop1: processing of precursor RNAs; Nop52: nucleolar protein 52; B23: protéine nucléolaire; PM-Scl 100: 100kDa-polymyositis/scleroderma antigen.
Figure 3
La transcription de l’ADN ribosomique.
1. L’ADN ribosomique (ADNr) est associé aux facteurs de transcription UBF (upstream binding factor) (vert) et SL1 (selectivity factor 1) (jaune) dans la région du promoteur. 2. L’ARN polymérase I (PolI) est recrutée sur le site d’initiation de la transcription (flèche rouge) par l’intermédiaire du facteur RRN3 (bleu) [51]. 3. Au cours de l’élongation, Pol I se déplace (sens de la flèche verte) jusqu’au site de terminaison associé au facteur de terminaison TTF1 (transcription termination factor 1) (rose), qui interagit également avec le promoteur [52]. Le point d’interrogation indique que la question de la participation de la dimérisation de TTF1 à l’organisation en boucle des gènes ribosomiques reste à élucider.
Figure 4
Nucléole et cycle cellulaire.
Le nucléole est présent pendant toute l’interphase (G1 à G2) (cercle rose) et est absent pendant la mitose (pointillé noir). Il se dissocie à la transition G2/M quand la machinerie de maturation se disperse à la périphérie des chromosomes et quand, de façon concomitante, la transcription l’ARN polymérase I (Pol I) est réprimée. Cependant, la machinerie de transcription Pol I reste assemblée sur les NOR (nucleolar organizer region). Pour réassembler un nouveau nucléole en sortie de mitose, il faut activer la transcription Pol I (NOR actifs) et recruter la machinerie de maturation sur les sites de transcription. Ce recrutement requiert la formation de corps nucléaires appelés PNB (pre-nucleolar bodies).