
Résumés
Résumé
La perspective biopsychologique systémique des émotions de base est un modèle heuristique permettant de mieux comprendre comment l’individu apprend à s’adapter à son environnement grâce aux différentes émotions qui se sont développées progressivement en même temps que la myélinisation des circuits neurohormonaux, de la naissance jusqu’à environ vingt et un ans. Ce sont ces mêmes émotions, agissant en complémentarité, qui vont permettre à l’individu de maintenir son équilibre tout au long de sa vie.
Cinq émotions de base ont été retenues, car dans la documentation scientifique, cinq circuits neuronaux associés aux émotions sont définis, soit les cinq circuits décrits par Panksepp : celui de l’agressivité (rage-colère), celui du stress (peur-surprise), développé par LeDoux, celui de la récompense (recherche-joie), développé par Tassin, celui de l’empathie (panique-tristesse), développé par Decety, et celui de la conscience (conscience-bonheur), développé par Damasio.
Mots-clés :
- émotions,
- circuits neurohormonaux,
- neurosciences,
- équilibre
Abstract
The systemic biopsychological perspective of basic emotions is a heuristic model that allows a better understanding of how people learn to adapt to their environment through different emotions that developed gradually along neurohormonal circuit myelination from birth until about the age of twenty-one. These same emotions, acting in complementarity, will allow the individual to maintain a balance throughout his life.
Five basic emotions were retained in line with the five emotions related to neuronal circuits, which are defined in the literature, and these are the five circuits described by Panksepp as follows: aggressiveness (Rage, angry), stress (Fear- surprise), developed by LeDoux, reward (Seeking-joy), developed by Tassin, empathy (Panic-sadness), developed by Decety, and consciousness (consciousness-happiness), developed by Damasio.
Several studies on myelination (Kinney, 1988, Parazzini, 2002, Deoni, 2012), Miller, 2012, and Welker, 2012) provide us with a scientific platform to determine the order of development of the neurohormonal circuits underlying basic emotions.
Neurohormonal circuits development begins at conception and will continue up until the age of 20-30 years. This article specifically addresses the first three years of life. It offers a systemic biopsychological perspective of basic emotions developed from the latest data in neuroscience. These informations have been integrated into a coherent whole that allows understanding the origin, the development and the functioning of basic emotions.
In addition to the information output from the thalamus to the midbrain that set in motion the somatic nervous system there exist, according to Roberge (1998), two other brain information sources that are managed by the hypothalamus (the limbic system). These two information sources allow the refining of the behavioural responses and they favour the homeostasis of the organism. The first information source goes from the midbrain to the hypothalamus to activate the peripheral nervous system. The latter is divided into two: the sympathetic (norepinephrine) that accelerates the motor response and the parasympathetic (acetylcholine), which slows it down. These two systems work in tandem. As for the second release of information, it is endocrine, thus it will follow the hypothalamus-pituitary-adrenal axis to cortisol, the hypothalamus-pituitary axis to endorphin and oxytocin and the hypothalamus-pineal axis to melatonin. The different emotional behaviours result from one of these two sources of information or from a combination of these two and are then managed by the limbic system, which is in continuous connection with the neocortex.
In short, no specific centre totally controls human behaviour. Control is achieved through a group of brain structures and relays, permitting adaptive behaviour and maintenance of balance by means of permanent exchanges. Anger, for instance, is a survival emotion, which allows protecting one’s physical integrity. It is very useful as an immediate response in an emergency situation, but it can also be harmful if it is used extensively in all situations, giving way to conduct disorders. Thus, the other neurohormonal circuits will regulate anger.
Emotions are an integral part of human behaviour. They allow the individual to constantly adapt to the physical and social environment. This approach brings a new perspective to understand how each person maintains balance to avoid the onset of clinical disorders. The understanding of neurochemical mechanisms underlying basic emotions opens up the door to several clinical applications.
Keywords:
- emotions,
- neurohormonal circuits,
- neuroscience,
- balance
Corps de l’article
Les recherches en neurosciences ont fait beaucoup de progrès au cours des dernières décennies et ont permis une meilleure compréhension du comportement humain. Plusieurs chercheurs ont fourni ces dernières années des fondements neuroscientifiques aux émotions de base. Panksepp (2011) et Damasio (2010) ont développé chacun un modèle selon trois niveaux. Ekman (2007) a identifié six émotions de base : la colère, le dégoût, la surprise, la peur, la joie et la tristesse. Récemment, Jack (2014) a conclu à l’existence de seulement quatre émotions de base. Le dégoût serait associé à la colère et la peur à la surprise, ce qui apporte un appui aux quatre émotions de base chez les animaux, décrites par Panksepp (1998), soit la rage, la peur, la recherche et la panique. Cinq émotions de base ont été retenues, car dans la documentation scientifique, cinq circuits neuronaux associés aux émotions sont définis, soit les cinq circuits décrits par Panksepp : celui de l’agressivité (rage-colère), celui du stress (peur-surprise), développé par Le Doux, celui de la récompense (recherche-joie), développé par Tassin, celui de l’empathie (panique-tristesse), développé par Decety, et celui de la conscience (conscience-bonheur), développé par Damasio.
L’émotion se définit comme une agitation passagère dans le déploiement normal des tendances relativement à une nouvelle situation. Lorsque des stresseurs viennent perturber l’équilibre, différents circuits neurohormonaux sont sollicités pour permettre le retour à l’homéostasie, amenant le changement à court terme du comportement. L’émotion est donc la réaction à court terme qui permet à l’individu de s’adapter à son environnement en maintenant son équilibre au jour le jour.
Plusieurs recherches sur la myélinisation (Kinney, 1988 ; Parazzini, 2002 ; Deoni, 2012), Miller, 2012 et Welker 2012) apportent des fondements scientifiques permettant de situer l’ordre de développement des circuits neurohormonaux qui sous-tendent les émotions de base. Le classement des émotions de base pour chacun des niveaux du modèle de Panksepp et du modèle de la biopsychologie systémique est présenté au Tableau 1.
Le cerveau fonctionne grâce à des circuits nerveux, hormonaux et immunitaires qui sont appelés circuits neurohormonaux. Ce sont des systèmes programmés à la base par les gènes qui se développent d’abord dans un environnement restreint, soit l’utérus de la mère, et par la suite dans un environnement social dès la naissance.
Le modèle biopsychologique systémique vise à mieux comprendre le développement et le fonctionnement des circuits neurohormonaux soutenant les émotions de base. Le développement commence dès la conception et s’échelonne jusque vers 20-30 ans. Cet article traite spécifiquement des trois premières années de vie. Il propose une perspective biopsychologique systémique des émotions de base élaborée à partir des récentes données en neurosciences. Ces données ont été intégrées en un tout cohérent permettant de comprendre l’origine, le développement et le fonctionnement des émotions de base. Chaque émotion de base est soutenue par des circuits neurohormonaux spécifiques : la colère par les circuits neurohormonaux de l’agressivité, la surprise par ceux du stress, la joie par ceux de la récompense, la tristesse par ceux de l’empathie et le bonheur par ceux de la conscience. Ce modèle va permettre une application clinique dont l’objectif est d’apporter aux clients une compréhension biopsychologique systémique pour les aider à retrouver et à maintenir leur équilibre.
Tableau 1
Différents modèles des émotions et de la conscience relativement à la myélinisation et au modèle biopsychologique systémique
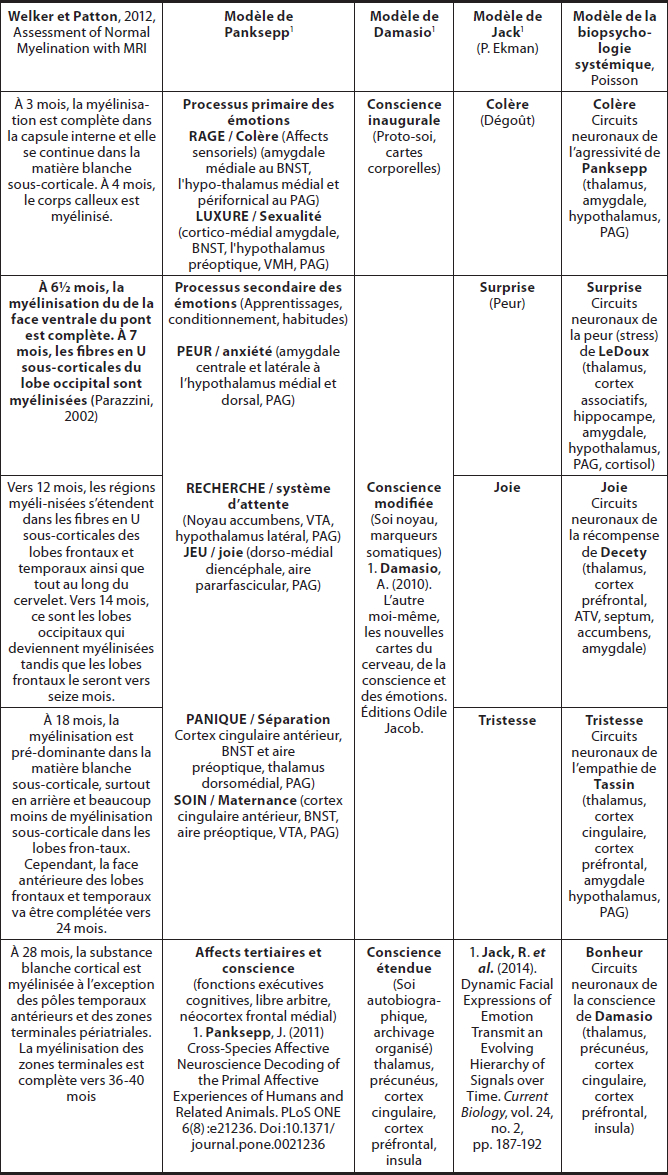
Panksepp et Damasio ont développé un modèle des émotions selon trois niveaux. Une proposition de classement des émotions pour chacun des niveaux y est présentée.
Origines des circuits neurohormonaux
Les gènes sont à la base de la spécificité des circuits neurohormonaux de chaque individu qu’on appelle l’hérédité. Selon Ginger (2002) et Zuckerman (2003), l’hérédité compte pour environ un tiers du comportement humain. Donc, la conception a son importance et peut avoir des répercussions sur le futur développement du foetus. Certaines personnes héritent de prédispositions plus favorables à l’adaptation alors que d’autres reçoivent une hérédité plus fragile entraînant davantage de difficultés à s’adapter.
Les apprentissages lors de la période foetale contribuent pour le deuxième tiers, influençant l’établissement du comportement. Dès les premiers mois de la vie du foetus, le développement des circuits neurohormonaux est soumis au climat utérin. Le troisième tiers influençant l’établissement du comportement débute à partir de la naissance. Le nouveau-né fera des apprentissages en fonction de son environnement physique et social, d’où l’importance d’un milieu stable et stimulant. C’est ainsi que l’environnement va moduler ce que l’individu a reçu génétiquement, tel que décrit par Anacker (2014) en épigénétique.
Panksepp (1998), Giedd (1999) et Schore (2008) ont précisé que les connexions entre les différentes structures du cerveau apparaissent graduellement avec la maturation cérébrale et qu’elles sont le résultat d’une longue sélection au cours de l’évolution.
Développement des circuits neurohormonaux
Le nouveau-né est un être essentiellement sensorimoteur. Toutes les structures neuronales sont présentes à sa naissance, mais les connexions ne sont pas encore toutes complétées. Selon Giedd (1999) et Welker (2012), certaines connexions sont bel et bien établies durant la période foetale, mais d’autres le seront graduellement après la naissance et permettront une plus grande variété de réponses comportementales. La myélinisation est responsable de ce processus.
La myélinisation du cerveau humain débute pendant la vie foetale et se poursuit après la naissance d’une manière prédéterminée et bien définie. Les études d’imagerie par résonance magnétique de Dietrich (1988) et Staudt (1993) ont montré que la myélinisation de la matière blanche sous-corticale part de la circonvolution péricentrale vers les lobes dans l’ordre suivant : lobes occipitaux et pariétaux vers cinq à six mois, lobes frontaux et temporaux vers neuf à onze mois et continue dans la deuxième décennie de la vie. En résumé, selon Parazzini (2002), la myélinisation se fait du centre vers la périphérie, de caudale (arrière) à rostrale (avant), de dorsale (supérieure) à ventrale (inférieure). Premièrement, les régions sensorielles visuelles et auditives sont myélinisées avant les régions motrices ; deuxièmement, les régions proches du sous-système neuroanatomie sont myélinisées avant les composantes plus distantes ; troisièmement, les voies de projection sont généralement myélinisées avant les voies associatives. La substance blanche péritrigonale est complètement myélinisée vers l’âge de 20 mois tout comme les lobes frontaux et temporaux, mais la myélinisation n’est pas complétée dans les lobes pariétaux. La myélinisation semble complète vers l’âge de 36 à 40 mois.
La myélinisation étant commencée lors de la période foetale, le nouveau-né expérimente dès le cinquième mois de vie foetal un premier sens, le toucher. Puis c’est le sens de l’ouïe qui devient fonctionnel vers le sixième mois, et ensuite le sens de l’odorat et du goût au début du troisième trimestre. La vue sera le dernier sens à prendre place chez le foetus, mais deviendra prédominante peu après la naissance.
Figure 1
Principaux circuits neuronaux du système nerveux somatique schématisés par Poisson

Selon Welker (2012), les régions myélinisées chez le nouveau-né sont les structures médullaires, la protubérance dorsale, les pédoncules cérébelleux inférieurs et supérieurs, le mésencéphale et le thalamus ventral latéral. La myélinisation est aussi intense dans les voies corticales vers la région périrolandique. Comme illustré à la Figure 1, les connexions se sont donc établies à partir des sens jusqu’aux cortex sensoriels primaires en passant par le thalamus sauf pour l’odorat qui passe directement au cortex olfactif avant de revenir au thalamus. Le thalamus est un centre d’intégration et de régulation de l’information. Il achemine celle-ci aux différents cortex primaires et de là, l’information passe aux cortex associatifs afin d’obtenir davantage d’informations pour permettre la réponse la plus adaptée. Cette première sortie d’information par le système nerveux somatique est tout simplement une réponse motrice automatique au niveau du mésencéphale, soit un réflexe. C’est de cette façon que se développent des routes et des autoroutes de l’information, appelées « circuits neuronaux ».
Les principaux circuits neurohormonaux de l’agressivité (colère)
Les circuits neurohormonaux de l’agressivité sont les premiers à se développer chez le nouveau-né. Ils favorisent l’émergence d’une première émotion, la colère. Elle permet au nouveau-né d’exprimer ses besoins physiologiques par des cris et des pleurs et ainsi d’alerter l’adulte responsable. Une réponse favorable à un besoin physiologique non répondu permettra de rétablir l’équilibre rompu. C’est le seul moyen que possède le nouveau-né pour communiquer sa douleur, car il est totalement dépendant des autres. Il fait un premier apprentissage de la colère pour exprimer ses besoins de survie, ce qui est déjà une première marque d’affirmation.
À deux mois, selon Welker (2012), il y a un haut signal de myélinisation des parties postérieures des capsules internes et du pédoncule cérébelleux. À trois mois, la myélinisation est complète dans la capsule interne et se continue dans la matière blanche cérébelleuse profonde et dans la substance blanche sous-corticale. À quatre mois, le corps calleux est myélinisé de la région antérieure à la région postérieure.
Ainsi, les premières connexions deviennent fonctionnelles entre le thalamus et le système limbique, comme illustré à la Figure 2. Le thalamus qui reçoit l’information à partir des différents cortex associatifs la transmet à l’insula antérieure qui est un centre d’évaluation des états viscéraux et au cortex somatosensoriel secondaire qui est un centre de l’évaluation de la douleur physique. Ces deux structures sont en étroite relation. Selon l’intensité du signal d’une ou de ces deux structures, l’amygdale, qui est souvent désignée comme le signal d’alarme, va être activée et enclenchera, en passant par l’hypothalamus, une réponse motrice. Panksepp (1998) décrit que les connexions du circuit neuronal de la rage passent de l’amygdale à l’hypothalamus médian puis au mésencéphale (substance grise périaqueducale, PAG) qui enclenche une réponse motrice automatique. Lorsqu’un besoin physiologique n’est pas comblé, Lacombe (2007) précise que l’amygdale va commander à l’hypothalamus, en passant par le mésencéphale (PAG), d’activer le système sympathique (augmentation de la pression sanguine, transformation des lipides en glucides, etc.). Celui-ci, à son tour, par les fibres préganglionnaires, va faire produire de l’adrénaline dans les médullosurrénales pour une réponse comportementale rapide, telle que la lutte ou la fuite, afin de combler ce besoin physiologique non répondu.
Figure 2
Principaux circuits neurohormonaux de l’agressivité (colère) selon Panksepp (1998) adaptés et schématisés par Poisson
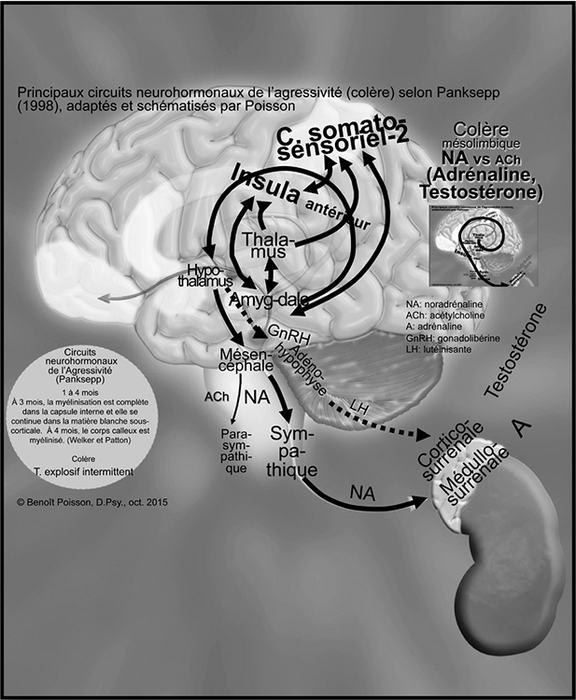
Les flèches  représentent les voies noradrénergiques. Les flèches pointillées
représentent les voies noradrénergiques. Les flèches pointillées
 représentent l’axe hypothalamo-hypophyso-surrénalien.
représentent l’axe hypothalamo-hypophyso-surrénalien.
Ce sont les voies noradrénergiques périphériques, prédominant dans ces circuits, qui sont à la base de la colère. Par contre, elles sont contrebalancées par les voies cholinergiques du système nerveux parasympathique. C’est ainsi qu’apparaît un premier niveau d’équilibre. En effet, lorsqu’il entre en fonction, le système parasympathique, connu comme le frein, permet de récupérer, de refaire des réserves et par conséquent, de rétablir l’homéostasie.
Les principaux circuits neurohormonaux du stress (surprise)
Les deuxièmes circuits neurohormonaux à prendre place sont ceux du stress. Ils permettent au nourrisson de reconnaître la différence entre maman, papa et les autres. Cela l’amène à réagir à ce qui est inconnu en sursautant ou en figeant. Il ne sourit plus spontanément à n’importe qui et il pourra détourner le regard pour fuir un nouveau visage. C’est ce qui est souvent appelé « période sauvage ». C’est de cette façon que cette deuxième émotion, la surprise, aide le nourrisson à apprendre à reconnaître ce qui lui est familier, ce qui lui procure la sécurité.
Selon Welker (2012), à six mois et demi, il y a un haut signal de myélinisation dans l'ensemble du corps calleux et la myélinisation de la face ventrale du pont est complétée. À sept mois, selon Parazzini (2002), les fibres en U sous-corticales du lobe occipital sont myélinisées.
Ainsi, les connexions neuronales entre le thalamus et le mésolimbique continuent de s’établir chez le nourrisson, comme illustré à la Figure 3. Selon LeDoux (1998), dans une situation de danger, mais non de survie, l’information passe du thalamus aux cortex associatifs unimodaux, ce qui permet de préciser l’objet du danger et d’envoyer cette information plus précise à l’amygdale. Parallèlement, à partir des cortex associatifs unimodaux, l’information se rend aux cortex associatifs polymodaux, ce qui permet de déterminer le concept, et à l’hippocampe, ce qui permet de préciser le contexte. Par la suite, ces analyses de l’information sont transmises à l’amygdale qui est davantage en mesure de commander la réponse la plus adaptée pour faire face à cette situation.
Lorsque l’hippocampe identifie un danger imminent, il envoie un message à l’amygdale, qui selon Ledoux (1998), va commander à l’hypothalamus d’activer le système sympathique en passant par le mésencéphale. Parallèlement, en sécrétant de la corticolibérine (CRF), l’hypothalamus active l’hypophyse qui va sécréter, à son tour, de l’adrénocorticotrophine (ACTH), stimulant la production de cortisol au sein des corticosurrénales pour maintenir le plus d’énergie possible afin de faire face à cette menace. C’est l’axe hypothalamo-hypophyso-surrénalien (HHS).
Ainsi, ces circuits neurohormonaux sont à la base de la surprise qui maintient un état de stress grâce à l’activation du système sympathique et à l’augmentation du cortisol. Cela permet au nourrisson de maintenir l’énergie nécessaire pour faire face au danger tant qu’il ne sera pas écarté. C’est le système inhibiteur de l’action (SIA) élaboré par Laborit (1970). Cependant, le cortisol est en interaction avec le système immunitaire. Lorsque le cortisol est en trop grande quantité, le système immunitaire s’affaiblit. Selon Lupien (2010), si une situation de stress persiste trop longtemps et que le niveau de cortisol devient trop élevé, cela peut conduire à l’épuisement et aux maladies psychosomatiques, telles que décrites par Selye (1955).
Figure 3
Principaux circuits neurohormonaux du stress (surprise) selon LeDoux (1998) schématisés par Poisson

Les flèches  représentent les voies noradrénergiques et les flèches pointillées
représentent les voies noradrénergiques et les flèches pointillées
 représentent l’axe hypothalamo-hypophyso-surrénalien.
représentent l’axe hypothalamo-hypophyso-surrénalien.
Lorsque le danger est écarté, le cortisol, qui a un effet de rétrocontrôle inhibiteur, va retourner un message à l’hippocampe lui indiquant que la situation est sous contrôle. À son tour, l’hippocampe va transmettre le message à l’amygdale pour faire cesser la production de CRF et la libération de l’ACTH, et permettre alors le retour à l’homéostasie. C’est ce deuxième niveau d’équilibre, grâce à l’axe HHS, qui permet au nourrisson de maintenir un sain développement.
Les principaux circuits neurohormonaux de la récompense (joie)
Le développement des circuits neurohormonaux de la récompense permet au nourrisson, qui commence à se déplacer, d’explorer et de découvrir son environnement, ce qui lui apporte du plaisir, d’où l’apparition d’une troisième émotion, la joie. Il fait plus d’activités par et pour lui-même. Le jeu prend une grande importance, lui permettant de développer de nouvelles habiletés tout en s’amusant.
Selon Welker (2012), vers 12 mois, les régions myélinisées s’étendent dans les fibres en U sous-corticales des lobes frontaux et temporaux de même que tout au long du cervelet. Vers quatorze mois, ce sont les lobes occipitaux qui deviennent myélinisés tandis que les lobes frontaux le seront vers seize mois.
Ainsi, les connexions du thalamus s’étendent maintenant jusqu’au cortex préfrontal qui est un centre de planification et de motivation de l’action de même que de stimulation de l’attention, comme illustré à la Figure 4. Selon Tassin (1998), ce niveau plus élevé d’action et d’attention entraîne une augmentation de la synthèse de dopamine dans l’aire tegmentale ventrale (ATV). Cette dopamine est envoyée dans le septum, ce qui déclenche la sensation de plaisir. De la dopamine est aussi envoyée dans les noyaux accumbens qui en informent l’amygdale et l’hypothalamus. L’augmentation de la dopamine dans les noyaux accumbens a un effet de renforcement sur les comportements qui permettent de satisfaire les besoins primaires. « Tous ces centres sont interconnectés et innervent l’hypothalamus, l’informant de la présence d’une récompense. L’hypothalamus agit alors en retour non seulement sur l’aire tegmentale ventrale, mais aussi sur les fonctions autonomes et endocrines de tout le corps par l’entremise de l’hypophyse » (Le plaisir et la douleur, McGill University, 2012). C’est le système de récompense qui favorise la satisfaction d’un besoin, ce qui amène la joie. Ce sont les voies dopaminergiques mésocorticolimbiques qui prédominent dans ces circuits neurohormonaux. Parallèlement, l’hypothalamus fait produire de la bêta-endorphine dans l’adénohypophyse qui va se déverser dans les noyaux accumbens, permettant l’établissement d’un renforcement positif selon Trezza (2010).
Figure 4
Principaux circuits neurohormonaux de la récompense (joie) selon Tassin (1998) adaptés et schématisés par Poisson
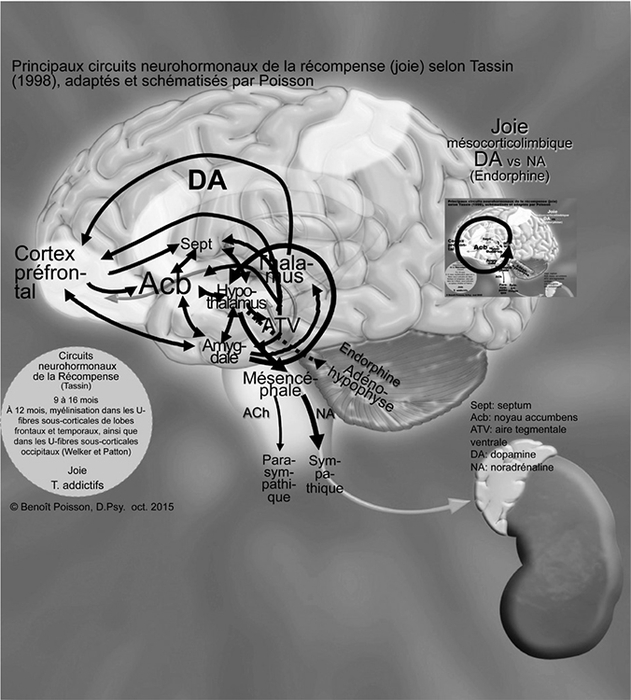
Les flèches  représentent les voies dopaminergiques et les flèches pointillées
représentent les voies dopaminergiques et les flèches pointillées
 représentent l’axe hypothalamo-hypophysaire.
représentent l’axe hypothalamo-hypophysaire.
Le nourrisson profite pleinement de cette phase lorsque les deux premiers circuits neurohormonaux sont en équilibre. Sinon, la dopamine sera transformée en noradrénaline dans le mésencéphale et si c’est nécessaire, en adrénaline dans les médullosurrénales pour retrouver l’équilibre au niveau des deux premiers circuits.
Les principaux circuits neurohormonaux de l’empathie (tristesse)
Le développement des circuits neurohormonaux de l’empathie permet au nourrisson la reconnaissance de soi (stade du miroir) et aussi la capacité de ressentir une nouvelle sensation : le mal-être, c’est-à-dire un profond malaise. Le nourrisson est aussi capable de ressentir ce mal-être chez les autres, grâce aux cellules miroirs (Decety, 2010). C’est ce qui le rend capable d’empathie. Cela lui permet de comprendre que l’autre a aussi des émotions. Il apprend qu’il n’est plus le centre du monde et qu’il doit tenir compte des besoins de l’autre, d’où l’apparition d’une nouvelle émotion : la tristesse. Il devient plus sensible à son environnement social puisqu’il n’est plus seulement centré sur ses besoins. Cela favorise la socialisation par le développement d’un réseau relationnel restreint à la famille immédiate dans un premier temps.
Figure 5
Principaux circuits neurohormonaux de l’empathie (tristesse) selon Decety (2010) adaptés et schématisés par Poisson
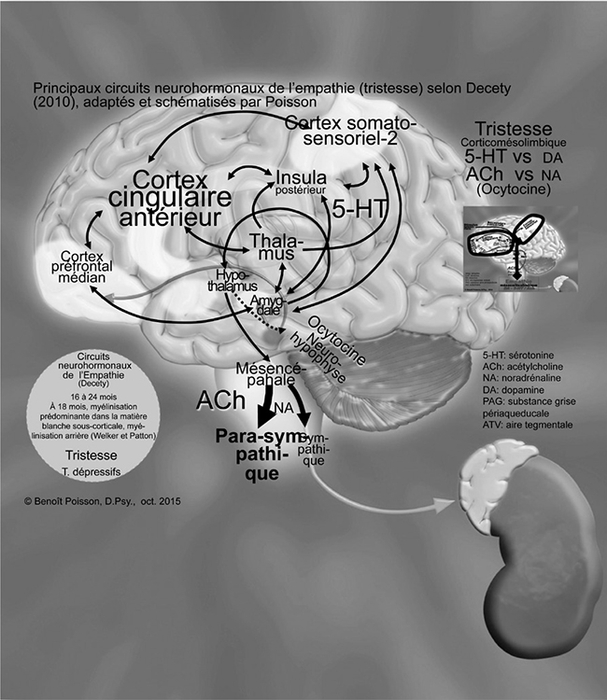
Les flèches  représentent les voies sérotoninergiques et les flèches pointillées
représentent les voies sérotoninergiques et les flèches pointillées
 représentent l’axe hypothalamo-hypophysaire.
représentent l’axe hypothalamo-hypophysaire.
À 18 mois, selon Welker (2012), la myélinisation est prédominante dans la matière blanche sous-corticale, surtout en arrière et beaucoup moins dans les lobes frontaux. La myélinisation de la matière blanche sous-corticale des lobes occipitaux est terminée vers l’âge de 15 mois. La face antérieure des lobes frontaux et temporaux va être complétée vers 24 mois.
Ainsi, les connexions continuent de s’établir au niveau du néocortex, comme illustré à la Figure 5. Selon Decety (2010), dans un premier temps, ce sont les connexions qui se font avec l’insula postérieure qui apporte l’information des états viscéraux et les cortex somatosensoriels secondaires, centres d’évaluation de la douleur physique. Ces connexions vont par la suite aux cortex cingulaires antérieurs qui apportent le côté émotif de la douleur, c’est-à-dire le mal-être, et aux cortex préfrontaux médians qui, selon l’intensité du mal-être, vont informer l’amygdale. Alors, l’amygdale est en mesure de commander à l’hypothalamus de ralentir l’activation, ce qu’il fait en activant le système parasympathique qui utilise de l’acétylcholine (ACh) comme neurotransmetteur et favorise de cette façon la récupération. Cette fois-ci, le principal neurotransmetteur est la sérotonine (5-HT), un neurotransmetteur qui, selon Roberge (1976), est en opposition avec la dopamine, car il a comme effet de ralentir l’activation. Parallèlement, l’hypothalamus produit de l’ocytocine dans la neurohypophyse qui va se déverser dans le cerveau, ce qui favorise l’attachement selon Ditzen (2009). Ce sont les voies sérotoninergiques qui prédominent dans ces circuits, mais qui sont contrebalancées par les voies dopaminergiques, ce qui crée le troisième niveau d’équilibre.
Les principaux circuits neurohormonaux de la conscience (bonheur)
Les cinquièmes circuits neurohormonaux à être fonctionnels sont ceux de la conscience. Selon Damasio (2010), ces connexions apportent une nouvelle capacité chez l’enfant, celle de la réflexion, ce qui favorise le développement du discernement et du jugement. Il apprend qu’il est un garçon ou une fille. Il commence à comprendre qu’il est un individu à part entière et un début de ce qui est bien ou mal va s’installer progressivement. C’est le début du développement de la morale et de l’acquisition de valeurs. L’enfant, lorsqu’il est en équilibre, a une bonne estime de soi, d’où l’apparition d’une cinquième émotion, le bonheur. Ce n’est qu’à long terme qu’il deviendra capable de se connaître véritablement et d’évaluer sa satisfaction personnelle.
Figure 6
Principaux circuits neurohormonaux de la conscience (bonheur) selon Damasio (2010) adaptés et schématisés par Poisson
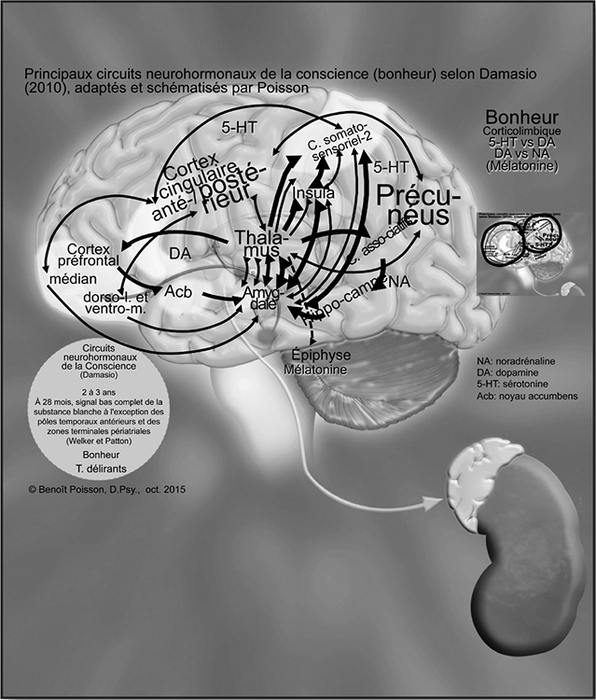
Les flèches  représentent les voies sérotoninergiques, celles-ci
représentent les voies sérotoninergiques, celles-ci  les voies dopaminergiques, celles-ci
les voies dopaminergiques, celles-ci  les voies noradrénergiques et les flèches pointillées
les voies noradrénergiques et les flèches pointillées  représentent l’axe hypothalamo-épiphysaire.
représentent l’axe hypothalamo-épiphysaire.
À 28 mois, selon Welker (2012), la substance blanche corticale est myélinisée, à l’exception des pôles temporaux antérieurs et des zones terminales périatriales. La myélinisation est complète vers 36-40 mois.
Selon Damasio (2010), les connexions se rendent jusqu’au précunéus, centre de détection des erreurs, comme illustré à la Figure 6. De là, elles vont aux cortex cingulaires postérieurs et aux cortex préfrontaux dorsolatéraux et ventromédians. Ce système évalue constamment l’homéostasie entre les systèmes sympathique (NA) et parasympathique (ACh). Il évalue parallèlement si le niveau de cortisol est stable (inhibiteur et activateur) et si la dopamine est contrebalancée par la sérotonine dans le cerveau. Lorsque l’homéostasie est atteinte, c’est à ce moment que les besoins de l’enfant sont satisfaits et qu’il se sent bien dans sa peau, d’où l’état de bonheur qui va perdurer jusqu’au prochain déséquilibre. Parallèlement, l’épiphyse produit de la mélatonine sous la supervision de l’hypothalamus, ce qui favoriserait un sommeil réparateur selon Christophe de Jaeger (2012) et ainsi une meilleure capacité d’apprentissage permettant à l’enfant d’évoluer sainement.
Vers trois ans, l’enfant apprend à utiliser de façon appropriée ces différents circuits neurohormonaux lui permettant de s’adapter à son environnement. Tous ces circuits neurohormonaux sont interdépendants et en interrelations permanentes afin de maintenir l’homéostasie. Chaque circuit a son utilité et prédomine selon la nécessité de la situation. C’est cette capacité de flexibilité dans l’utilisation appropriée de chaque émotion selon le contexte qui varie d’une personne à l’autre et qui détermine la capacité d’adaptation de chacun.
Le processus de myélinisation est rapide pendant les trois premières années, suivi d’une progression lente jusqu’à 20-30 ans selon Hayakawa (1991). La myélinisation se poursuit tout au long de la vie. C’est la plasticité du cerveau qui permet à la personne de faire de nouveaux apprentissages tout au long de sa vie afin de maintenir son équilibre et de permettre une saine adaptation à son environnement physique et social.
Fonctionnement des circuits neurohormonaux
En plus de la sortie d’information du thalamus vers le mésencéphale afin de mettre en branle le système nerveux somatique, selon Roberge (1998), il existe deux autres sorties d’information du cerveau qui sont gérées par l’hypothalamus (système limbique), tel qu’illustré à la Figure 7. Ces deux sorties d’information permettent de peaufiner les réponses comportementales favorisant l’homéostasie de l’organisme. La première sortie d’information part de l’hypothalamus vers le mésencéphale afin d’activer le système nerveux périphérique. Ce dernier se subdivise en deux : le sympathique (noradrénaline) qui accélère la réponse motrice et le parasympathique (acétylcholine), qui la ralentit. Ces deux systèmes fonctionnent en complémentarité. Quant à la deuxième sortie d’information, elle est endocrinienne, c’est-à-dire qu’elle emprunte l’axe hypothalamus-hypophyso-surrénalien pour le cortisol, l’axe hypothalamus-hypophyse pour les endorphines et l’ocytocine, et l’axe hypothalamus-épiphyse pour la mélatonine. Les différents comportements émotifs résultent d’une de ces deux sorties d’information ou de leur combinaison gérées par le système limbique qui est en lien continu avec le néocortex.
Figure 7
Synthèse des principaux circuits neurohormonaux des cinq émotions de base schématisés par Poisson

Les flèches  représentent les voies sérotoninergiques, celles-ci
représentent les voies sérotoninergiques, celles-ci  les voies dopaminergiques et celles-ci
les voies dopaminergiques et celles-ci  les voies noradrénergiques.
les voies noradrénergiques.
Par exemple, selon l’origine de l’information, le thalamus va envoyer cette dernière au système limbique (amygdale) qui, à son tour, par l’entremise du mésencéphale, si c’est une question de survie dans l’immédiat, va activer le système sympathique afin de déclencher la production d’adrénaline dans les médullosurrénales. Cela fera produire très rapidement un surplus d’énergie pour faire face à cette perturbation. Si c’est une question de protection face à un danger, l’information va passer par le système limbique (hippocampe) et c’est le système sympathique qui va être encore activé. Cependant, l’axe hypothalamo-hypophyso-surrénalien va être enclenché afin de produire du cortisol dans les corticosurrénales, permettant ainsi de maintenir l’énergie pour éviter la perte de contrôle due à cette perturbation. Par ailleurs, si la situation ne présente pas un enjeu de survie ou de protection, l’information sera orientée du thalamus vers les différents cortex (préfrontal, cingulaire et autres) pour en obtenir une analyse plus détaillée et permettre un comportement adapté à la situation, tel que l’exploration, le maintien d’une relation ou la réflexion.
L’équilibre s’exerce à trois niveaux. Le premier est au sein du système nerveux autonome, sympathique (noradrénaline) ou parasympathique (acétylcholine), le deuxième au sein des corticosurrénales, cortisol activateur ou inhibiteur, et le troisième au sein du cerveau, dopamine ou sérotonine.
En somme, aucun centre spécifique ne contrôle la totalité du comportement humain. Ce contrôle est exercé par un ensemble de structures cérébrales et de relais qui permettent, par des échanges permanents, une réponse adaptée et ainsi le maintien de l’équilibre. La colère, par exemple, est une émotion de survie qui permet de protéger son intégrité physique. Elle est très utile en situation d’urgence pour une réponse immédiate. Mais elle peut aussi être néfaste si elle est utilisée intensément et sans égard aux situations, car elle pourrait alors entraîner des troubles des conduites ou du comportement. Ce sont donc les autres circuits neurohormonaux qui vont intervenir pour réguler la colère.
La Figure 8 illustre un schéma de la biopsychologie systémique des émotions de base et de certains troubles cliniques qui arrivent lorsqu’il y a une perte importante d’équilibre. L’ordre d’apparition des différents circuits neurohormonaux est présenté de même que les différents circuits neurohormonaux en lien avec la myélinisation.
Figure 8
Schéma de la biopsychologie systémique des émotions de base et de certains troubles cliniques

Conclusion
En s’appuyant sur les récents constats de la recherche en neurosciences, le modèle heuristique présenté dans cet article permet de mieux comprendre comment se sont développés les circuits neurohormonaux soutenant les émotions de base et comment celles-ci fonctionnent en complémentarité. Les émotions font partie intégrante du comportement humain. Elles permettent à l’individu de s’adapter constamment à son environnement physique et social. Cette approche apporte une nouvelle perspective pour comprendre comment chaque personne maintient son équilibre afin d’éviter l’apparition de troubles cliniques. Elle ouvre la porte à des applications cliniques grâce à la compréhension des mécanismes neurochimiques qui sous-tendent les émotions de base.
Parties annexes
Note
-
[1]
L’auteur remercie sincèrement Dre Andrée G. Roberge, Ph. D., pour son soutien tout au long de la réalisation de cet article.
Bibliographie
- Anacker, C., O’Donnell, K. J. et Meaney, M. J. (2014). Early life adversity and the epigenetic programming of hypothalamic-pituitary-adrenal function. Dialogues Clinical Neuroscience, 16, 321-333.
- Damasio, A. (2010). L’autre moi-même, les nouvelles cartes du cerveau, de la conscience et des émotions. Paris, France : Éditions Odile Jacob.
- Decety, J. (2010). La force de l’empathie. Cerveau & Psycho. 38, 43-49.
- Deoni, S. C. L., Dean, D. C., O’Muircheartaigh, J., Dirks, H. et Beth A. Jerskey, B. A. (2012). Investigating white matter development in infancy and early childhood using myelin water faction and relaxation time mapping. Neuroimage, 63(3), 1038–1053.
- Dietrich, R. B., Bradley, W. G., Zaragoza, E. J., Ottd, R. J., Taira, R. K., Wilson, G. H. et Kangarloo, H. (1988). MR Evaluation of Early Myelination Patterns in Normal and Developmentally Delayed Infants. AJNR 9,69-76.
- Ditzen, B., Schaer, M., Gabriel, B., Bodenmann, G., Ehlert, U. et Heinrichs, M. (2009).Intranasal oxytocin increases positive communication and reduces cortisol levels during couple conflict. Biology Psychiatry, 1 ; 65(9), 728-731.
- Ekman, P. (2007). Emotions Revealed, Second Edition : Recognizing Faces and Feelings to Improve Communication and Emotional Life. New York : Holt Paperbacks.
- Giedd, J. N. et al. (1999). Brain development during childhood and adolescence : a longitudinal MRI study. Nature neuroscience, 2(10), 861-863.
- Ginger, S. (2010). Cerveau féminin, cerveau masculin. Récupéré le 26 mars 2012 sur le site de l’auteur : http://sergeginger.net/resources/Cerveau+f$C3$A9minin$3Acerveau+masculin.pdf
- Hayakawa, K., Konishi, Y., Kuriyama, M., Konishi. K. et Matsuda, T. (1991). Normal brain maturation in MRI. European Journal of Radiolology, 12(3), 208-215.
- Jack, R. E., Garrod, O. G. B. et Schyns, P. G. (2014). Dynamic Facial Expressions of Emotion Transmit an Evolving Hierarchy of Signals over Time. Current Biology, 24(2), 187-192.
- Jaeger, C. (2012). La mélatonine. Dans Les techniques de lutte contre le vieillissement, 95-110, Que Sais-je, no 3463, Paris, France : PUF.
- Kinney, H. C., Brody, B. A., Kloman, A. S. et Gilles, F. H. (1988). Sequence of central nervous system myelination in human infancy. II. Patterns of myelination in autopsied infants. J. Neuropathology Exp Neurology, 47(3), 217-234.
- Laborit, H. (1970). L’agressivité détournée : Introduction à une biologie du comportement social. Éd. Union Générale d’Edition, coll. « 10/18 ».
- Lacombe, M. (2007). Le Lacombe : précis d’anatomie et de physiologie humaine, volume 1. France, Éditions Lamarre.
- LeDoux, J. (1998). The Emotional Brain. London, U.K. : Weidenfeld & Nicolson.
- Le plaisir et la douleur. Récupéré le 20 mars 2012 du site : Le cerveau à tous les niveaux : http://lecerveau.mcgill.ca/flash/index_a.html
- Lupien, S. (2010). Par amour du stress. Montréal, Canada : Éditions au carré.
- Miller, D. J., Duka, T., Stimpson, C. D., Steven J., Schapiro, S., Baze, W. B., McArthur, M. J., Fobbs, A. J., /Sousa, A. M., Šestan, N., Wildman, D. E., Lipovich, L., Kuzawa, C. W., Patrick R., Hof, P., Chet C. et Sherwood, C. C. (2012). Prolonged myelination in human neocortical evolution. PNAS, 109(41), 16480-16485.
- Panksepp, J. (1998). Affective neuroscience. New York, N.Y. : Oxford University Press.
- Panksepp, J. (2011) Cross-Species Affective Neuroscience Decoding of the Primal Affective Experiences of Humans and Related Animals. PLoS ONE, 6(8), e21236.
- Parazzini, C, Baldoli, C, Scotti, G et Triulzi F. (2002). Terminal zones of myelination : MR evaluation of children aged 20-40 months. American Journal of Neuroradiology. 23(10), 1669-1673.
- Roberge, A. G., Parent, A. et Boulay, M. (1976). Démonstration d’une relation inversement proportionnelle entre la dopamine et la sérotonine dans certaines structures cérébrales : aspects neurochimique et morphologique. Journal of Neurochemistry, 26, 591-595.
- Roberge, A. G. (1998). Le système neuro-endocrino-immunitaire : un trio indissociable. Le clinicien, septembre, pp. 185-194.
- Schore, A. N. (2008). La régulation affective et la réparation du soi. Montréal. Les éditions du CIG.
- Selye, H. (1955). Selye explique les mécanismes du stress. www.youtube.com/watch?v=ZH37hdGiwVw, 2012.
- Staudt, M., Schropp, M., Staudt, C., Obletter, N., Bise, K. et Breit, A. (1993). Myelination of the brain in MRI : A staging system. Pediatric Radiology, 23(3), 169-176.
- Tassin, J.-P. (1998). Drogues, plaisir et douleur. La recherche, 306.
- Trezza, V., Damsteegt, R., Achterberg E. J. et Vanderschuren, L. J. (2011). Nucleus accumbens μ-opioid receptors mediate social reward. The Journal of Neuroscience, 31(17), 6362-6370.
- Welker, K. M. et Patton, A. (2012). Assessment of normal myelination with magnetic resonance imaging. Seminar in Neurology, 32, 15-28.
- Wright, N. D., Bahrami, B., Johnson, E., Di Malta, G., Rees, G., Frith, C. D., Dolan, R. J. (2012). Testosterone disrupts human collaboration by increasing egocentric choices. Proceedings Biological Sciences, 279(1736) :2275-2280.
- Zuckerman, M. (2003). La IIIe révolution du cerveau : Psychobiologie de la personnalité. Paris, France : Payot.
Liste des figures
Figure 1
Principaux circuits neuronaux du système nerveux somatique schématisés par Poisson
Figure 2
Principaux circuits neurohormonaux de l’agressivité (colère) selon Panksepp (1998) adaptés et schématisés par Poisson
Les flèches représentent les voies noradrénergiques. Les flèches pointillées
représentent l’axe hypothalamo-hypophyso-surrénalien.
Figure 3
Principaux circuits neurohormonaux du stress (surprise) selon LeDoux (1998) schématisés par Poisson
Les flèches représentent les voies noradrénergiques et les flèches pointillées
représentent l’axe hypothalamo-hypophyso-surrénalien.
Figure 4
Principaux circuits neurohormonaux de la récompense (joie) selon Tassin (1998) adaptés et schématisés par Poisson
Les flèches représentent les voies dopaminergiques et les flèches pointillées
représentent l’axe hypothalamo-hypophysaire.
Figure 5
Principaux circuits neurohormonaux de l’empathie (tristesse) selon Decety (2010) adaptés et schématisés par Poisson
Les flèches représentent les voies sérotoninergiques et les flèches pointillées
représentent l’axe hypothalamo-hypophysaire.
Figure 6
Principaux circuits neurohormonaux de la conscience (bonheur) selon Damasio (2010) adaptés et schématisés par Poisson
Les flèches représentent les voies sérotoninergiques, celles-ci les voies dopaminergiques, celles-ci les voies noradrénergiques et les flèches pointillées représentent l’axe hypothalamo-épiphysaire.
Figure 7
Synthèse des principaux circuits neurohormonaux des cinq émotions de base schématisés par Poisson
Les flèches représentent les voies sérotoninergiques, celles-ci les voies dopaminergiques et celles-ci les voies noradrénergiques.
Figure 8
Schéma de la biopsychologie systémique des émotions de base et de certains troubles cliniques
Liste des tableaux
Tableau 1
Différents modèles des émotions et de la conscience relativement à la myélinisation et au modèle biopsychologique systémique
Panksepp et Damasio ont développé un modèle des émotions selon trois niveaux. Une proposition de classement des émotions pour chacun des niveaux y est présentée.