Résumés
Résumé
La concentration en oxygène dissous en milieu fluvial varie selon un cycle diurne (24 h) qu’il est essentiel de considérer dans l’évaluation de l’état d’oxygénation d’un cours d’eau. En principe, seules des mesures en continu recueillies au cours de cycles de 24 h permettent d’évaluer correctement l’état d’oxygénation d’une rivière, ce que, en pratique, les contraintes logistiques et budgétaires ne permettent pas de réaliser. Le présent article vise à faire la synthèse des connaissances sur les facteurs de contrôle et la modélisation des variations diurnes de la concentration en oxygène dissous en rivière. Parmi les facteurs biologiques et physico-chimiques, les activités autotrophe et hétérotrophe sont les facteurs dominants responsables des variations diurnes de l’oxygène dissous. Certains modèles de qualité de l’eau permettent de modéliser la teneur en oxygène et, dans certains cas, la variation diurne. Toutefois, ces modèles sont souvent complexes, d’utilisation ardue et impliquent la mesure directe sur des cycles de 24 h des variables qui régissent la concentration en oxygène dans le milieu. Une démarche est proposée pour l’élaboration d’un modèle et a été appliquée dans une étude de cas réalisée dans la rivière Saint-Charles (Québec, Canada). Un modèle simple (une fonction sinusoïdale) dont les paramètres ont été corrélés à la température et à la concentration moyenne en nitrate a permis de générer des valeurs simulées d’oxygène dissous très proches des valeurs observées in situ. Un modèle alternatif utilisant des valeurs ponctuelles de température et de concentration de nitrate a donné des résultats équivalents. L’approche proposée constitue donc une alternative simple et pratique à la mesure en continu de l’oxygène et permet une évaluation plus réaliste de l’état réel d’oxygénation d’une rivière que la prise de mesures ponctuelles.
Mots clés:
- oxygène,
- dynamique,
- modélisation,
- cycle diurne,
- qualité de l’eau,
- rivière,
- photosynthèse,
- respiration
Summary
In rivers, dissolved oxygen concentrations typically show diel variations with maximum values during daytime and minimum values at night. The diel cycle must be taken into account when assessing the state of oxygenation of a watercourse. However, in water quality monitoring programs, dissolved oxygen concentrations are usually obtained from single measurements taken during daytime. The resulting data do not represent the real overall oxygen levels of a watercourse and thus can lead to erroneous conclusions regarding the oxygen status of a river. Continuous data collected over 24‑hour cycles are required for an accurate oxygen status assessment, but in practice, logistic and budget constraints do not allow such samplings. Modelling can be a convenient alternative to direct measurements. However, the water quality models that take into account the diel cycle of oxygen are generally complex to run. The purpose of this study was to review the information relating to the dynamics and the modelling of the diel variations in dissolved oxygen in rivers and to apply a simple model in a case study involving dissolved oxygen, nutrients, temperature and chlorophyll a data collected over 24‑hour cycles in the St. Charles River near Quebec City (Canada).
Photosynthesis by algae, both benthic and planktonic, as well as by macrophytes, is an important and sometimes dominant factor in the oxygen budget of a river. Sediments and heterotrophic activity by bacteria, particularly in rivers receiving important loads of wastewaters, can be important sinks for oxygen. Temperature determines the solubility of oxygen, thereby directly influencing oxygen concentrations. Diel variations in oxygen thus reflect diel variations in temperature. Temperature also has an effect on biological processes such as respiration and photosynthesis. Reaeration varies not only with temperature, but also with the type of flow (laminar vs. turbulent) and current velocity. In rivers with important slopes and current velocities, reaeration can be sufficient to make up for oxygen losses due to high heterotrophic activity. Nevertheless, light is the first causative factor for the diel variations in oxygen, determining both autotrophic activity and water temperature. However, suspended matter in the water column reduces light penetration. Higher levels of suspended matter result in lower levels of photosynthesis and oxygen production. The sudden or large influx of runoff waters after heavy rain or snowmelt can also have an important impact on the oxygen budget of a river. Finally, chemical factors can have an influence on the diel variations in oxygen: nutrient inputs, in particular, can stimulate the rate of photosynthesis and oxygen production. Overall, oxygen dynamics are determined by the relative importance of biological, physical and chemical factors, which vary in time and space. However, biological processes often dominate over the other causative factors affecting diel variations in oxygen. In temperate climates, biological processes have a controlling function only during warmer months, with temperature and flow being the dominant controlling factors during colder months.
Water quality models that take into account diel variations in oxygen are often designed to assess primary production and respiration. These models are based on either ODUM’s (1956) concept of oxygen curves or on direct and continuous measurements of dissolved oxygen. Periodic functions or Fourier series are also used to simulate diel variations in oxygen. The widely used USEPA QUAL2e model predicts diel variations in oxygen from different measurements including light intensity and from computation of the rates of photosynthesis and respiration. Several other modelling approaches use various combinations of indirect methods to predict the variations in oxygen, based on light intensity, algal biomass, primary production, and reaeration. Specific models are sometimes necessary due to particular regional characteristics or environmental issues.
In general, water quality models designed for water management purposes are complex and require the measurement of a large number of parameters, which necessitates elaborate and costly logistics. Estimating the parameters controlling the oxygen budget in a river thus ends up being more time and labour consuming than the direct measurement of diel variations in oxygen. A simpler model leading to the estimation of the concentrations and the amplitude of the variations of dissolved oxygen was developed and applied to the St. Charles River.
The St. Charles River flows from the Laurentians north of Quebec City to the St. Lawrence River. The vast upper watershed is mostly forested, but the lower part is heavily urbanized. Two stations were located in the upper watershed and one in the section of the river within Quebec City. Water quality was excellent at the upstream stations and poor at the downstream station, due to wastewater inputs and low flow rates. Sampling was carried out in the months of July and August of 1996 and 1997. Physico-chemical and light measurements were made every two hours for periods of 24 hours. Nutrient and chlorophyll a samples were collected every four hours. Small diel variations in oxygen (amplitude: 1.48 mg/L) were observed at the upstream station, while much larger ones (amplitude: 4.23 mg/L) were measured at the downstream station. Modelling of the diel variations in oxygen was carried out using a sine function. Oxygen concentrations over 24 hours were successfully predicted from average concentration of dissolved oxygen, amplitude, and phase of the cycle. Average oxygen concentrations and amplitude can be derived from physico-chemical and/or biological variables easily measured in standard water quality monitoring programs. Average oxygen concentrations showed a very strong correlation with temperature (r2 = 0.91) and amplitude of oxygen level variations was strongly correlated with average nitrate concentration (r2 = 0.58). These relations were used in the sine function and resulted in significantly correlated modelled and measured oxygen concentrations for six of the seven cycles. Overall correlation between modelled and observed values was high (r2 = 0.77). Modelled values obtained with single measurements of temperature (taken at 2:30 P.M.) and nitrate concentration (which shows no diel variation) were also highly correlated with observed data (r2 = 0.75). Absolute and relative bias as well as root-mean-square error also showed the validity and the equivalence of the two approaches.
This study shows that simple models based on available water quality data may generate realistic oxygen values over 24‑hour cycles. These models would be a valuable diagnostic and decision making tool for the management of water quality in rivers.
Key words:
- oxygen dynamics,
- modelling,
- diel cycle,
- water quality,
- river,
- photosynthesis,
- respiration
Corps de l’article
1. Introduction
La concentration en oxygène dissous en milieu fluvial suit une variation cyclique caractérisée par des valeurs maximales durant le jour et minimales durant la nuit, souvent avant l’aurore (O’CONNOR et DI TORO, 1970). Bien que la variation cyclique de la concentration en oxygène soit connue et acceptée, il n’en demeure pas moins que ce processus est peu abordé dans les manuels de référence portant sur l’écologie aquatique, la qualité de l’eau ou la chimie des eaux. Lorsqu’elle y est discutée, seuls les principaux facteurs de contrôle de la variation de l’oxygène en rivière y sont brièvement mentionnés, notamment l’influence de l’activité autotrophe et hétérotrophe en relation avec la productivité du milieu, de même que l’effet de la température sur les concentrations en oxygène (p. ex. WETZEL, 2001; HORNE et GOLDMAN, 1994). Ainsi, l’ensemble des facteurs de contrôle biologiques, physiques et chimiques des fluctuations diurnes de l’oxygène n’y sont pas traités de façon complète et détaillée, de même que de l’importance relative de ces facteurs selon différents contextes (p. ex. rivière en milieu agricole, présence de rejets d’origine anthropique).
La variabilité diurne de l’oxygène dissous est rarement prise en considération dans les protocoles de suivi de la qualité de l’eau en milieu fluvial. Dans les programmes de suivi de la qualité de l’eau, la concentration en oxygène dissous est la plupart du temps mesurée de façon ponctuelle pendant le jour, soit durant la période où la concentration en oxygène est la plus élevée. Souvent, ces valeurs ne sont donc pas représentatives des conditions d’oxygénation réelles des cours d’eau. Une comparaison des mesures ponctuelles prises durant le jour avec des données obtenues sur une période de 24 h montre que les plus faibles valeurs de l’échantillonnage en continu se situent bien en-dessous de la plupart des valeurs de l’échantillonnage ponctuel de jour (LIVINGSTONE, 1991). Les conditions d’oxygénation du cours d’eau sont de cette façon surestimées et, par le fait même, non représentatives des conditions réelles. Cette surévaluation des conditions d’oxygénation peut avoir des conséquences importantes dans le cas d’études sur la qualité de l’eau où les concentrations mesurées pourraient suggérer une bonne oxygénation de l’eau alors que des concentrations très faibles peuvent survenir durant la nuit, en particulier dans le cas de rivières soumises aux activités anthropiques.
La prise en considération des fluctuations diurnes de l’oxygène en rivière est très peu souvent recommandée dans les manuels de références sur l’écologie aquatique ou encore portant sur les méthodes d’échantillonnage pour l’évaluation de la qualité de l’eau. L’importance de bien considérer les variations de la concentration de l’oxygène dissous à court terme dans un plan d’échantillonnage est à tout le moins soulevée par HAMMER et MAC KICHAN (1980). WETZEL et LIKENS (2000) mentionnent aussi qu’il est possible d’avoir à prendre plusieurs mesures dans un plan d’eau afin d’avoir une estimation des conditions moyennes d’oxygénation pour une période de temps donnée.
Afin d’obtenir un portrait représentatif de la variabilité de la concentration en oxygène en rivière, des mesures devraient être prises en continu sur une période de 24 h. Les exigences et l’investissement que demande la prise de mesures en continu sont considérables : disponibilité sur une base régulière de personnel compétent pour la calibration et l’entretien des instruments de mesure; quantité suffisante d’instruments pour permettre le remplacement chaque semaine des équipements pour l’entretien et la calibration; et compétence et temps nécessaire pour la gestion des bases de données (NEAL et al., 2002). Pour des programmes d’échantillonnage couvrant un large territoire, ce type de mesure est pratiquement impossible à réaliser pour des raisons budgétaires et logistiques. La modélisation de la dynamique de l’oxygène en rivière peut être la solution afin de remédier aux contraintes relatives à la prise de mesures en continu de la concentration en oxygène dissous. La modélisation des fluctuations de l’oxygène est possible car la variabilité de l’oxygène est cyclique et peut se modéliser à l’aide d’une fonction périodique.
Certains modèles de qualité de l’eau en milieu fluvial prennent en considération la variabilité diurne de l’oxygène et découlent du concept de courbes journalières développé par ODUM (1956) qui permet d’estimer le métabolisme (photosynthèse et respiration) à partir des changements diurnes de l’oxygène le long d’un segment de rivière. Souvent ces modèles de qualité de l’eau ont été créés afin d’estimer les taux de production primaire et de respiration (p. ex. le modèle Delta, CHAPRA et DI TORO, 1991; le modèle DORM, GULLIVER et STEFAN, 1984a et 1984b), alors que d’autres estiment la variabilité diurne de la concentration en oxygène par le calcul du taux de photosynthèse et de respiration, tel que dans le modèle de qualité de l’eau QUAL2e de l’US EPA (BROWN et BARNWELL, 1987). Ces modèles sont souvent complexes et exigent l’obtention d’un nombre important de données pour plusieurs paramètres, alors que l’utilisation de mesures directes peut s’avérer plus fiable et plus facile. Dans la littérature, il ne semble pas exister de modèles simples utilisant les cycles diurnes de l’oxygène pour estimer l’amplitude des variations de l’oxygène en milieu fluvial.
La présente étude vise à approfondir les connaissances sur la dynamique et la modélisation de l’oxygène en rivière et à appliquer un modèle simple à une étude de cas basée sur des données d’oxygène dissous, d’éléments nutritifs, de température et de chlorophylle a (Chl a) recueillies au cours de cycles diurnes dans la rivière Saint-Charles, près de Québec (Canada).
2. Facteurs de contrôle de la variabilité diurne
2.1 Facteurs biologiques
2.1.1 Activité autotrophe
L’activité photosynthétique par les algues et les macrophytes peut être le facteur dominant des variations diurnes de l’oxygène (O’CONNOR et DI TORO, 1970), et ainsi faire augmenter la concentration en oxygène en rivière sur une période de 24 h (LIVINGSTONE, 1991; MÜLLER et WEISE, 1987). Une corrélation significative (r = ‑ 0,503; α = 0,01) entre les concentrations en Chl a phytoplanctonique et les taux de saturation d’oxygène mesurés le matin dans le fleuve Garonne (France) a été démontrée par BENGEN et al. (1992), ce qui suggère que le pourcentage matinal de saturation en oxygène est influencé principalement par l’activité biologique. Cette relation inverse reflète en fait une respiration algale plus élevée durant la nuit pour une concentration en Chl a élevée, sachant que les mesures ont été prises le matin, juste après le minimum de concentration en oxygène alors qu’il y avait encore peu d’apport d’oxygène par la photosynthèse.
Le lien entre la concentration en oxygène en rivière et celle en Chl a phytoplanctonique n’est pas toujours clair dans la littérature (CHESTERIKOFF et al., 1991; UCHRIN et al., 1988). L’activité des algues benthiques plutôt que planctoniques aurait parfois une influence plus forte sur les cycles diurnes. La contribution de la biomasse des algues benthiques aux variations diurnes de l’oxygène est évidente dans les rivières sous influences anthropiques (milieu agricole, BACHMANN et al., 1988 et WILCOCK et al., 1995; rejet d’eaux usées, QUINN et MCFARLANE, 1989). Sur plusieurs cycles diurnes, les concentrations en oxygène mesurées dans une rivière dont le bassin de drainage comprend principalement des fermes laitières se situaient sous les 50 % de saturation et variaient selon une amplitude maximale de plus de 25 % (WILCOCK et al., 1995). Une rivière recevant les eaux de ruissellement de cultures de maïs et de soja a montré une amplitude moyenne de 62 % de saturation et des valeurs minimales et maximales moyennes de 59 % et 121 % (BACHMANN et al., 1988). Une comparaison a été faite par SABATER et al. (2000) entre un site en milieu ouvert et sous influence anthropique (riche en éléments nutritifs) et dont la concentration en Chl a benthique était élevée (923 mg m‑2), avec un site en forêt, pauvre en éléments nutritifs et caractérisé par une faible biomasse d’algues périphytiques (0,007 mg m‑2). L’amplitude des variations journalières était beaucoup plus élevée au premier site (10 mg L‑1) qu’au second (1,5 mg L‑1), ce qui suggère que la biomasse d’algues benthiques contribue considérablement aux variations journalières de concentrations en oxygène. L’impact de l’activité benthique en milieu eutrophe a aussi été démontré par RAUCH et VANROLLEGHEM (1998) dont les conclusions sont basées sur une étude de cas virtuelle, dans laquelle les données simulent un rejet d’eaux usées municipales dans une rivière.
La production primaire élevée dans une rivière en zone agricole et en milieu ouvert, et où l’on observe de fortes fluctuations de l’oxygène, semble être aussi attribuable aux épiphytes et aux macrophytes (BACHMANN et al., 1988). WILLIAMS et al. (2000) expliquent d’ailleurs la forte amplitude des cycles diurnes de l’oxygène par le recouvrement élevé (80‑90 %) du lit de la rivière par les macrophytes. Dans le cas d’une rivière enrichie par les activités urbaines et agricoles, de fortes fluctuations diurnes de l’oxygène dissous (70‑140 % de saturation) reflètent la combinaison d’un bas niveau de l’eau et d’un recouvrement élevé en macrophytes dont l’activité photosynthétique est favorisée par un faible débit (NEAL et al., 2002). Cependant, les macrophytes semblent n’avoir qu’une influence partielle sur les cycles diurnes de l’oxygène, car le métabolisme d’une rivière caractérisée par une très forte biomasse en macrophytes n’est pas nécessairement dominé par l’activité de celles-ci (KAENEL et al., 2000). Les plantes ripariennes peuvent aussi causer indirectement une déplétion de la concentration en oxygène dissous par leur décomposition dans le lit de la rivière (WILCOCK et al., 1995).
2.1.2 Activité hétérotrophe
Les variations de la concentration en oxygène à court terme seraient aussi influencées par l’activité hétérotrophe, et ce, particulièrement dans les rivières très anthropisées. Dans la Seine, l’apport de bactéries allochtones provenant d’un débordement d’égouts serait responsable de la totalité de la consommation d’oxygène pour des événements à court terme (environ 24 h) (SEIDL et al., 1998). L’apport de matière organique d’origine naturelle terrestre ou amenée par les eaux d’infiltration souterraines peut aussi avoir une influence sur le taux de respiration (autotrophe et hetérotrophe) et par le fait même, sur la dynamique de l’oxygène (JACOBS et al., 1988; HOWARTH et al., 1992).
Le processus de nitrification peut s’avérer très important dans le bilan de la concentration en oxygène dissous en rivière à court terme (heures), particulièrement dans les cas où il y a présence de rejets d’eaux usées. Ainsi, des rejets d’ammoniaque d’une usine de traitement des eaux usées peuvent être la cause d’une chute de la concentration journalière moyenne en oxygène (CHAUDHURY et al., 1998). Ce déclin de la concentration en oxygène peut être provoqué par la croissance des bactéries nitrifiantes qui est favorisée par une charge ponctuelle d’ammoniaque d’origine industrielle (DUNNETTE et AVEDOVECH, 1983; RICKERT, 1984). La déplétion maximale en oxygène dissous est aussi attribuable en grande partie à la DBON (demande biochimique en oxygène associée à l’azote) pour une rivière qui reçoit les effluents d’une usine de traitement des boues, ceci basé sur un modèle construit à l’aide de données prises toutes les 4 h pendant 36 h et 64 h (GOWDA, 1983). La nitrification semble aussi être un facteur non négligeable dans le bilan de l’oxygène de la Seine en raison des rejets d’effluents riches en ammoniaque (GARNIER et al., 2001).
La demande en oxygène des sédiments (DOS) peut aussi constituer une perte prédominante en oxygène dans une rivière recevant les rejets d’une usine de traitement des eaux usées municipales (VAN ORDEN et UCHRIN, 1993) ou pour une rivière dans laquelle s’accumulent des sédiments en période d’étiage à cause de la présence de réservoirs le long de son cours (CHAUDHURY et al., 1998). Il est à noter que la DOS inclut l’activité benthique autotrophe en plus de l’activité bactérienne et des réactions chimiques d’oxydation, ce qui ne permet pas de discriminer l’influence relative de ces facteurs dans le bilan de l’oxygène.
2.2 Facteurs physiques
2.2.1 Température
Le cycle diurne de la concentration en oxygène d’une rivière est aussi influencé par la température de l’eau qui a un effet sur le taux de réaération et sur l’activité des microorganismes aquatiques (LIVINGSTONE, 1991). La solubilité de l’oxygène dans l’eau diminue lorsque la température augmente, ce qui induit une diminution de la concentration en oxygène à saturation. Ce phénomène peut être compensé par un apport en oxygène tel que par la photosynthèse. Or, la température influence le taux de consommation d’oxygène par les organismes durant le jour (effet de l’énergie solaire sur la température de l’eau). Cette influence de la température sur le taux de consommation en oxygène doit être prise en compte dans le calcul du bilan de l’oxygène. BUTCHER et COVINGTON (1995) mentionnent aussi que la prédiction de la variabilité de l’oxygène dans des rivières peu profondes et caractérisées par une forte variabilité diurne de la température est plus performante en considérant les taux de réaération, de respiration et de photosynthèse comme étant fonction de la température. La température de l’eau semble d’ailleurs être un facteur de contrôle important de la biomasse des algues benthiques (relation linéaire r = 0,50; p < 0,05) avec les éléments nutritifs, influençant ainsi la variabilité journalière en oxygène dissous (SABATER et al., 2000). Selon les résultats d’une expérience in situ effectuée dans une rivière en milieu agricole, la température de l’eau serait plus importante que les nutriments dans le contrôle de la croissance du périphyton (BUSHONG et BACHMANN, 1989).
2.2.2 Réaération
La réaération correspond à l’apport d’oxygène à travers l’interface air-eau et varie en fonction de la température de l’eau (voir ci-dessus) et du débit, du type d’écoulement (laminaire vs turbulent) ou de la vitesse du courant. Les échanges d’oxygène air-eau peuvent être suffisamment élevés pour compenser les pertes en oxygène dues à une forte activité hétérotrophe. Ceci peut s’observer dans le cas d’une rivière polluée et caractérisée par une forte pente et une vitesse de courant élevée favorisant par le fait même la réaération (WILCOCK et al., 1995). Selon cette même étude, la réaération pourrait ainsi permettre de réduire les risques d’anoxie durant la nuit. D’autre part, les échanges gazeux air-eau peuvent être insuffisants pour équilibrer les gains et les pertes en oxygène occasionnés par l’activité biologique dans une rivière enrichie en nutriments et où de fortes fluctuations diurnes sont observées (NEAL et al., 2002). Dans le cas de rivières non perturbées, la réaération peut avoir un impact plus important que l’activité biologique sur les variations diurnes de l’oxygène, plus spécifiquement pour une rivière sous couvert végétal avec un débit relativement plus élevé, soit de 80 à 150 L sec‑1 (0,08 à 0,15 m3 sec‑1), en comparaison avec une rivière en milieu ouvert et dont le débit est plus faible, soit de 3 à 12 L sec‑1 (0,018 à 0,003 m3 sec‑1), pour laquelle les variations diurnes sont influencées par la réaération et l’activité biologique (GUASCH et al., 1998). Le vent est un facteur qui a une influence sur les échanges gazeux et ainsi sur la réaération, et donc peut avoir un effet sur les variations diurnes de l’oxygène. Cette variable est d’ailleurs incluse dans l’estimation du coefficient de réaération dans le modèle DORM servant à l’analyse du métabolisme des rivières (GULLIVER et STEFAN, 1984a et 1984b).
2.2.3 Lumière
Le cycle diurne de l’oxygène est fortement influencé par la disponibilité de la lumière car elle conditionne d’une part l’activité autotrophe, et d’autre part, les variations de la température de l’eau (LIVINGSTONE, 1991). La température et la lumière sont d’ailleurs les facteurs environnementaux prédominants dans le contrôle de la production primaire (SABATER et al., 2000). Or, la photosynthèse et la température de l’eau sont deux facteurs déterminants dans les variations diurnes de l’oxygène dissous en rivière tels que décrits précédemment.
La concentration en matière en suspension influence aussi la production primaire de façon indirecte par la diminution de la quantité de lumière disponible pour la photosynthèse. L’effet de la luminosité sur les variations diurnes de l’oxygène peut s’observer lors de certains événements de courte durée (quelques heures), comme par exemple lors de l’arrivée des eaux d’évacuation d’un réservoir. Celle-ci engendre une augmentation de la turbidité de l’eau par la remise en suspension des sédiments et, par le fait même, une diminution de la pénétration de la lumière. Ceci a pour effet de faire chuter l’activité photosynthétique et de faire augmenter les taux de respiration algale et bactérienne, causant ainsi une diminution de la concentration en oxygène dans le milieu et une perturbation du cycle diurne de l’oxygène (BARILLIER et al., 1993). Ce genre d’événement s’apparente à une augmentation du débit après un orage qui fait augmenter la turbidité de l’eau et modifie le cycle diurne de l’oxygène. De plus, les eaux de ruissellement apportées à la rivière lors de pluies intenses sont susceptibles d’être chargées entre autres de matière organique dissoute. Une partie de cette matière organique dissoute peut être colorée (appelée CDOM, en anglais, coloured dissolved organic matter) et comprend essentiellement des substances humiques qui proviennent de la végétation et des sols du bassin versant. Ces substances affectent la pénétration de la lumière dans l’eau (voir WETZEL, 2001) et par le fait même, possiblement les fluctuations diurnes de l’oxygène.
2.2.4 Événements météorologiques ponctuels
Certains événements météorologiques ponctuels sont susceptibles d’intervenir dans le cycle diurne de l’oxygène en rivière. Les précipitations et les eaux de ruissellement sont chargées en oxygène et constituent ainsi un apport en oxygène pour le milieu. Les eaux de fonte des neiges apportées à la rivière viennent aussi modifier le bilan journalier en oxygène de différentes façons : elles font augmenter le taux de réaération par l’augmentation du débit; elles provoquent une augmentation de la concentration à saturation par l’abaissement de la température de l’eau; elles causent une diminution du taux de consommation de l’oxygène car elles contiennent moins de matériel organique; et finalement, elles favorisent une augmentation de la concentration en oxygène dissous par apport direct (LIVINGSTONE, 1991).
2.3 Facteurs chimiques
L’apport d’éléments nutritifs influe sur le cycle diurne de l’oxygène en favorisant l’activité photosynthétique. Les processus d’oxydation photochimique (photolyse) peuvent aussi avoir une influence sur les fluctuations diurnes de l’oxygène dissous. La photolyse des composés organiques dissous implique en effet la consommation d’oxygène et est ainsi susceptible d’engendrer une perte directe en oxygène dissous (BREZONIK, 1994 et LAANE et al., 1985). L’oxydation chimique comme celle du fer présent dans les sédiments est aussi une perte potentielle mais probablement mineure dans le bilan de l’oxygène. Ces types de réactions liées à l’oxydation chimique ne sont d’ailleurs pas soulevés comme facteur de contrôle dans les études abordant les variations diurnes de l’oxygène.
En somme, l’importance relative des facteurs biologiques, physiques et chimiques détermine la dynamique de l’oxygène en rivière. L’activité biologique demeure néanmoins le facteur dominant responsable des variations diurnes de l’oxygène dissous. En climat tempéré, l’effet de l’activité biologique sur les variations diurnes est toutefois moindre durant les périodes plus froides de l’année. L’activité photosynthétique contrôlerait ainsi les changements en oxygène dissous durant l’été, alors que durant les mois plus froids, les forts débits et les basses températures seraient les facteurs de contrôle dominants de la concentration en oxygène dissous en milieu fluvial (ANTOINE et BOWKER, 1996). Par ailleurs, l’amplitude des fluctuations diurnes de l’oxygène durant l’été est plus grande que celle mesurée durant les mois plus froids à cause de l’activité plus élevée du phytoplancton et des macrophytes durant les mois les plus chauds (NEAL et al., 2002).
3. Modélisation de la dynamique de l’oxygène en rivière
La modélisation dans le domaine de la qualité de l’eau en général connaît un essor important depuis les années 1970 avec le développement de modèles mathématiques dans le but d’aider à la planification et à la prise de décision en gestion de l’eau. La revue et l’évaluation des nombreux modèles de qualité de l’eau ont été faites dans plusieurs ouvrages de référence (CHAPRA, 1997; FALCONER, 1992; McCUTCHEON, 1989; PALMER, 2001; TCHOBANOGLOUS et SCHROEDER, 1985) et publications scientifiques (AMBROSE et al., 1988; MORIN et al., 1984). Le but de cette section est d’identifier les modèles qui prennent en considération ou simulent la variabilité diurne de l’oxygène dissous en rivière. Certains sont d’application générale, c’est-à-dire sans restriction géographique, alors que d’autres sont beaucoup plus spécifiques à une situation ou à une région donnée.
3.1 Modèles généraux
Les modèles de qualité de l’eau en milieu fluvial qui tiennent compte de la variabilité diurne de l’oxygène ont souvent été créés afin d’estimer les taux de production primaire et de respiration. Ces modèles découlent du concept de courbes journalières développé par ODUM (1956) qui permet d’estimer le métabolisme (photosynthèse et respiration) à partir des changements diurnes de l’oxygène le long d’un segment de rivière. Cette méthode est basée sur le principe que la photosynthèse, la respiration et la réaération sont responsables des fluctuations diurnes de l’oxygène en rivière. Plusieurs auteurs ont utilisé les courbes journalières de la concentration en oxygène dissous pour estimer ou simuler le métabolisme global d’une rivière donnée (p. ex. BACHMANN et al., 1988; ERDMANN, 1979a et 1979b; KELLY et al., 1974; SCHURR et RUCHTII, 1977; SIMONSEN et HARREMOËS, 1978; WILLIAMS et al., 2000). KELLY et al. (1974) et SCHURR et RUCHTII (1977) se sont basés sur la méthode d’Odum mais ont procédé à des mesures directes et en continu pour obtenir les variations de l’oxygène dissous. Cette méthode est plus précise et moins susceptible d’introduire un biais dans l’estimation des concentrations en comparaison avec l’utilisation de chambres d’incubation in situ selon la méthode d’Odum. La méthode de KELLY et al. (1974) et de SCHURR et RUCHTII (1977) implique une technique d’analyse basée sur une fonction continue (séries de Fourier) afin d’estimer les taux de production primaire et de respiration. La méthode Delta de CHAPRA et DITORO (1991) est basée sur les équations mathématiques de O’CONNOR et DITORO (1970) et inclut aussi l’utilisation de séries de Fourier, mais propose une approche plus facile de l’estimation des taux de photosynthèse et de respiration à l’aide de données en continu d’oxygène dissous. Cette estimation de la photosynthèse par les séries de Fourier est utilisée par exemple par STAHL et UCHRIN (1989) afin de simuler les variations diurnes de l’oxygène. MARZOLF et al. (1994) procèdent aussi à l’estimation du métabolisme global avec des données en continu d’oxygène, mais aussi à l’aide de mesures in situ du métabolisme autotrophe (chambre d’incubation) et du coefficient de réaération, alors que ce dernier est généralement estimé par modélisation. Cette utilisation des données diurnes de l’oxygène afin de déterminer les taux de photosynthèse et de respiration est aussi appliquée dans le modèle DORM (GULLIVER et STEFAN, 1984a et 1984b). Certaines des méthodes pour évaluer la production primaire citées ici sont présentées et décrites par BOWIE et al. (1985).
D’autres modèles d’application générale qui prennent en considération la variabilité diurne de l’oxygène ont été créés dans le but de prédire les fluctuations de l’oxygène dissous, en plus d’estimer le métabolisme global de la rivière. Le modèle de qualité de l’eau QUAL2e de l’USEPA permet de simuler le comportement de nombreuses variables telles que l’oxygène (BROWN et BARNWELL, 1987). Ce modèle estime la variabilité diurne de la concentration en oxygène à l’aide de différentes mesures (p. ex. la luminosité) et par le calcul du taux de photosynthèse et de respiration. Il a été utilisé entre autres afin de simuler la variabilité de l’oxygène dissous en fonction de la demande biochimique en oxygène et de la photosynthèse (New Jersey, VAN ORDEN et UCHRIN, 1993); d’établir les facteurs majeurs influençant la qualité de l’eau pour le développement des objectifs environnementaux de rejets (CHAUDHURY et al., 1998); et d’évaluer de nouvelles stratégies de gestion pour un fleuve à Taiwan (NING et al., 2001). Le modèle QUASAR est aussi un modèle de qualité de l’eau pouvant être d’application générale et permettant de modéliser diverses variables de qualité de l’eau le long d’une rivière, dont la concentration en oxygène dissous (WHITEHEAD et al., 1997). Dans ce modèle, l’estimation de la photosynthèse se fait avec la mesure de l’intensité lumineuse et de l’estimation de la biomasse algale. EL-SHAARAWI et al. (1988), pour leur part, proposent des méthodes indirectes pour prédire les fluctuations de la concentration en oxygène. Ces méthodes consistent en l’analyse de régression non linéaire des données d’oxygène prises à chaque heure et sert à estimer les paramètres (p. ex. la luminosité) nécessaires à la détermination des facteurs de gains et de pertes (p. ex. la production primaire) pour l’équation de base du bilan massique de l’oxygène. La modélisation peut aussi être utile dans l’interprétation des variations diurnes de l’oxygène, comme dans le cas du modèle de LIVINGSTONE (1991) qui est basé sur des approximations polynomiales des principaux facteurs dictant les variations diurnes, soit la réaération, la production primaire et la respiration, qui sont alors estimés à l’aide de mesures in situ de la température de l’eau. L’utilité de cette démarche est que tous les paramètres ne doivent pas nécessairement être modélisés contrairement à la plupart des modèles.
3.2 Modèles spécifiques
Les caractéristiques particulières d’une région ou d’une problématique donnée demandent parfois la création de modèles de qualité de l’eau plus spécifiques et souvent d’application plus limitée. Le modèle DOMOD3 par exemple a été créé afin d’avoir un aperçu des effets relatifs des pertes sur le déficit en oxygène et aussi pour prédire la répartition spatiale de l’oxygène selon différents paramètres tels que le débit et la température. Ce modèle est basé sur les formulations d’O’CONNOR et DITORO (1970) et a comme objectif spécifique d’évaluer l’effet de la nitrification sur l’état d’oxygénation de la rivière Speed localisée dans le sud de l’Ontario (Canada, GOWDA, 1983). Le modèle WIRQAS DO a plutôt été créé afin de prévoir l’effet de la charge de polluants et du débit sur les conditions d’oxygénation le long du fleuve Willamette (États-Unis, RICKERT, 1984). Il a été appliqué par DUNNETTE et AVEDOVECH (1983) pour évaluer l’impact d’un apport majeur en ammoniaque sur le régime de l’oxygène. Un autre exemple de modèle spécifique est celui de QUINN et MCFARLANE (1989) dont le champ d’application est limité à une rivière en Nouvelle-Zélande. Ce modèle sert à prédire les variations de l’oxygène dissous à court terme (heures) afin de déterminer les taux de respiration et les biomasses épilithiques permissibles pour maintenir des concentrations acceptables en oxygène. Finalement, le modèle Sam-1D/Rive sert à estimer les variations de l’activité microbienne et de l’oxygénation dans l’estuaire de la Seine. Il permet de rendre compte des variations à court terme de l’oxygénation telles qu’observées à l’aide de mesures en continu (BILLEN et al., 1999).
En somme, le but commun des modèles de qualité de l’eau qui tiennent compte des variations diurnes de l’oxygène est soit d’évaluer le métabolisme global d’une rivière en estimant la production primaire et la respiration par la modélisation des variations diurnes de l’oxygène, soit de prédire les variations de la concentration en oxygène dissous, le plus souvent dans un but de gestion de l’eau (p. ex. l’effet de réduction de rejets anthropiques, l’amélioration du traitement des eaux avant rejet en rivière, l’enlèvement des plantes aquatiques). Ces modèles sont toutefois complexes et demandent la mesure d’un nombre considérable de paramètres, ce qui exige une logistique élaborée et coûteuse. L’estimation des paramètres responsables du bilan en oxygène devient alors plus fastidieuse que de mesurer directement les fluctuations journalières de l’oxygène. L’élaboration d’un modèle simple permettant de déterminer la concentration moyenne en oxygène dissous et l’amplitude des variations est toutefois possible et sera présentée dans l’étude de cas de la rivière Saint-Charles.
3.3 Processus de modélisation : démarche suggérée
L’élaboration d’un modèle mathématique nécessite le respect d’une démarche spécifique afin d’assurer la validité du modèle. Voici un résumé des étapes proposées par McCUTCHEON (1989) et FALCONER (1992) :
Choix et délimitation de la zone d’étude et élaboration d’un plan d’échantillonnage;
Création du modèle à l’aide de séries de mesures en continu de l’oxygène dans un ensemble de rivières pour une région déterminée, ainsi que d’autres paramètres mesurés en simultané (p. ex. la concentration en Chl a et en phosphore);
Calibration du modèle afin de choisir les coefficients des termes de l’équation et les limites du modèle;
Validation du modèle avec un ensemble de données autre que celui utilisé pour la calibration et qui aura été obtenu lors d’une deuxième campagne d’échantillonnage;
Définition des limites du modèle : la saison (étiage, crue), la région et le type de rivière selon les caractéristiques physicochimiques et biologiques et les particularités du bassin versant (p. ex. les types d’activités anthropiques) doivent être clairement définis afin d’orienter l’utilisateur dans l’application du modèle.
4. Étude de cas : modélisation de l’oxygène dans la rivière Saint-Charles
4.1 Site d’étude et échantillonnage
La rivière Saint-Charles prend sa source dans les Laurentides et son embouchure donne sur le fleuve Saint-Laurent après avoir traversé la ville de Québec. Très anthropisé dans sa section aval, le bassin versant de la Saint-Charles est largement forestier dans sa partie amont.
L’échantillonnage a été effectué à l’embouchure de la rivière des Hurons et à deux stations sur la rivière Saint-Charles. Ces deux rivières sont respectivement l’affluent principal et l’effluent du lac Saint-Charles (Figure 1). La rivière Saint-Charles a été échantillonnée à la sortie du lac, juste en aval du barrage (RSC1), ainsi que dans la zone urbaine de la ville de Québec, à 7 km de son embouchure (RSC2). Le bassin versant de la rivière Saint-Charles couvre une superficie totale de 513 km2 tandis que celui de la rivière des Hurons draine un territoire principalement forestier de 136 km2. Le débit annuel moyen de la rivière Saint-Charles est de 8,29 m3/s (valeurs pour la période de 1969 à 1992 et mesurées à la station débitmétrique localisée entre les stations RSC1 et RSC2; (HÉBERT, 1995)). Les prélèvements dans la rivière Saint-Charles pour l’approvisionnement en eau potable sont en moyenne d’environ 150 000 m3 par jour (environ 1,7 m3 sec‑1), ce qui correspond approximativement à 20 % du débit annuel moyen (LAVALLÉE, 1983; FILLION et al., 1987). La pente moyenne aux stations RSC1 et RSC2 est respectivement de 0,3 % et 0,1 % (HÉBERT, 1995) et celle à la station Hurons est aussi relativement faible (0,1 %) (GERARDIN et LACHANCE, 1997). La pente de la station RSC1 a peu de signification ici compte tenu que cette station se situe juste en aval du barrage du lac Saint-Charles, d’une hauteur de 2,5 m.
Figure 1
Localisation des stations d’échantillonnage dans le bassin versant des rivières Saint-Charles et des Hurons (modifié de LÉGARÉ 1998).
Location of the sampling stations in the watershed of the St. Charles and des Hurons rivers (modified from LÉGARÉ 1998).

L’eau à la station RSC1 est qualifiée d’excellente alors que celle à la station en milieu urbain (RSC2) est considérée de mauvaise qualité à cause de la pollution urbaine et industrielle qui affecte la qualité de l’eau (LÉGARÉ, 1998). La mauvaise qualité de l’eau à cette station serait due à la combinaison du faible débit de la rivière et des apports de certains effluents municipaux et des nombreux débordements d’égouts qui surviennent lors des pluies. La qualité de l’eau à la rivière des Hurons est bonne malgré la présence d’habitations à son embouchure qui engendrent une légère contamination bactériologique due à l’absence de moyens adéquats d’évacuation et de traitement des eaux usées (HÉBERT, 1995).
L’échantillonnage a été réalisé en 1996 et 1997 aux mois de juillet et août, soit en conditions d’étiage et de forte activité biologique, afin d’obtenir les variations journalières maximales de la concentration en oxygène dissous. Un total de neuf séries de données physicochimiques et biologiques (température et concentrations en oxygène dissous, en éléments nutritifs et en Chl a) ont été obtenues sur 24 h pour les stations Hurons, RSC1 et RSC2 (Tableau 1).
Tableau 1
Cycles diurnes mesurés aux stations d’échantillonnage Hurons, RCS1 et RCS2.
Diurnal cycles measured at the Hurons, RSC1 and RSC2 stations.
Cycle |
Date |
Station |
Donnée et fréquence de mesure |
|---|---|---|---|
1 |
5 au 6 août 1996 |
RSC2 |
O2, T et luminosité mesurés à chaque 2 h Ntot, NO3 et Chl a mesurés à chaque 4 h |
2 |
7 au 8 août 1996 |
Hurons |
|
3 |
7 au 8 août 1996 |
RSC1 |
|
4 |
26 au 27 août 1996 |
Hurons |
|
5 |
26 au 27 août 1996 |
RSC1 |
|
6 |
17 au 18 juillet 1997 |
Hurons |
O2, T et luminosité mesurés à chaque 2 h Ptot, PTD, Ntot, NO3 et Chl a mesurés à chaque 4 h |
7 |
17 au 18 juillet 1997 |
RSC2 |
|
8 |
23 au 24 juillet 1997 |
Hurons |
|
9 |
23 au 24 juillet 1997 |
RSC2 |
O2 : oxygène dissous, T : température, Ptot : phosphore total, PTD : phosphore total dissous,
Ntot : azote total, NO3 : nitrate
La concentration en oxygène dissous et la température ont été mesurées à l’aide d’une multisonde (Reporter, Hydrolab Corp.) munie d’un mélangeur et d’une boite de contrôle (Surveyor 3, Hydrolab Corp.). Les mesures ont été prises à des intervalles d’environ 2 h pour des périodes de 24 h à partir des ponts qui surplombent les rivières. La luminosité a été mesurée à toutes les deux heures à l’aide d’un photomètre portatif (QSL-100, Biospherical Instruments). Des échantillons d’eau ont été prélevés à toutes les quatre heures pour l’analyse de la concentration en éléments nutritifs et en chlorophylle (Chl a). Les analyses pour le phosphore ont été réalisées au Laboratoire national des essais environnementaux (NLET) du Centre canadien des eaux intérieures à Burlington (Ontario, Environnement Canada). Celles pour les composés azotés l’ont été au Centre d’expertise en analyse environnementale du Québec (Sainte-Foy, ministère de l’Environnement du Québec). Le dosage de la Chl a a été effectué à l’aide d’un fluorimètre après extraction des pigments à l’éthanol.
4.2 Modélisation
4.2.1 Cycles de la concentration en oxygène
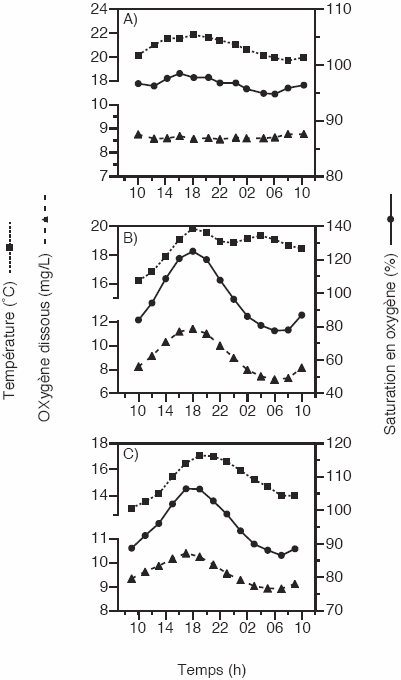
La variabilité cyclique de la concentration en oxygène dissous sur une période de 24 h est évidente à la station RSC2 de la rivière Saint-Charles (Figure 2). L’amplitude du cycle à cette station est de 4,23 mg/L, ce qui correspond à des variations du taux de saturation en oxygène de 77 % à 125 %. La concentration en oxygène à la station de la rivière des Hurons montre aussi une variation diurne mais de moindre amplitude (1,48 mg/L). Contrairement aux stations RSC2 et Hurons, la concentration en oxygène obtenue à la station RSC1 demeure pratiquement constante et près du point de saturation (moyenne : 97 % de saturation). L’eau évacuée à la sortie du barrage situé en amont de la station crée un brassage constant, ce qui assure une bonne réaération de l’eau. En conséquence, les données retenues pour analyse sont celles pour lesquelles une variation cyclique a été observée, soit aux stations RSC2 et Hurons.
Figure 2
Cycles de la concentration en oxygène dissous (mg/L), du taux de saturation en oxygène (%) et de la température (oC) dans les rivières Saint-Charles (RSC1 (a), RSC2 (b)) et des Hurons (c).
Cycles of dissolved oxygen concentration (mg/L), oxygen saturation (%) and temperature (oC) in the St. Charles river (RSC1 (a), RSC2 (b)) and des Hurons river (c).
4.2.2 Modélisation du cycle diurne de l’oxygène
L’estimation des variations diurnes de la concentration en oxygène dissous est possible aux stations RSC2 et Hurons à l’aide d’une fonction de type sinusoïdal (1).
où
O2 |
concentration en oxygène dissous à un temps (t) donné; |
Cm |
concentration moyenne en oxygène dissous; |
A |
amplitude du cycle; |
B |
phase de l’oscillation; |
t |
temps. |
Plusieurs tests ont démontré qu’il y a une bonne correspondance entre les données mesurées et celles obtenues avec le modèle lorsque B = ‑p. Les paramètres Cm et A doivent par la suite être estimés par régression avec les variables physicochimiques et/ou biologiques qu’il est facile d’obtenir à l’intérieur d’un programme de suivi.
Estimation des paramètres du modèle
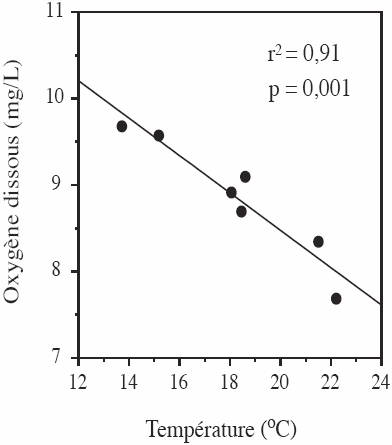
La relation significative entre la concentration moyenne en oxygène et la température pour les sept cycles obtenus aux stations Hurons et RSC2 (r2 = 0,91, p = 0,001; (Figure 3)) permet de substituer le paramètre Cm dans l’équation (1) :
Figure 3
Relation entre la concentration moyenne en oxygène (mg/L) et la température moyenne (oC) sur 24 heures des cycles diurnes mesurés aux stations Hurons et RSC2.
Relationship between the mean oxygen concentration (mg/L) and temperature (oC) over 24 hours of the diurnal cycles measured at the Hurons and RSC2 stations.
La concentration en oxygène suit d’ailleurs une oscillation semblable à celle de la température pour les cycles observés aux stations Hurons et RSC2 (Figure 2).
Les régressions effectuées entre l’amplitude (paramètre A) et les différentes variables mesurées indiquent que seule la concentration moyenne en nitrate montre une corrélation significative (r2 = 0,58) avec l’amplitude des cycles de concentration en oxygène obtenues aux stations Hurons et RSC2 (Figure 4). La relation entre l’amplitude et la concentration moyenne en nitrate est la suivante :
Figure 4
Relation entre l’amplitude de variation des cycles de concentration en oxygène (mg/L) et la concentration moyenne en nitrate (mg/L) aux stations Hurons et RSC2.
Relationship between the amplitude of the cycles in oxygen concentration (mg/L) and mean nitrate concentration (mg/L) at the Hurons and RSC2 stations.

Selon les corrélations obtenues, il est alors possible de remplacer les paramètres Cm et A dans l’équation (1) par les équations (2) et (3). L’estimation des fluctuations diurnes de la concentration en oxygène peut ainsi être faite selon le modèle suivant :
Les courbes diurnes obtenues à l’aide du modèle sont comparables aux données mesurées aux stations RSC2 et Hurons (Figure 5). La régression entre les données modélisées et les valeurs mesurées est significative pour six des sept cycles (r2 = 0,38 à 0,99, p < 0,001 à 0,06), alors qu’elle est non significative que pour le cycle 1 (r2 = 0,15, p = 0,18). La concentration moyenne en oxygène et l’amplitude des variations obtenues par modélisation pour le cycle 1 (concentration moyenne = 8,15 mg/L, amplitude = 1,11 mg/L) demeurent néanmoins semblables aux données mesurées (concentration moyenne = 8,35 mg/L, amplitude = 1,21 mg/L). Une forte relation est aussi observée entre les données modélisées et les observations réelles (r2 = 0,77, p < 0,0001 (Figure 6)) des sept cycles regroupés.
Figure 5
Comparaison des cycles de concentrations en oxygène mesurées et estimées (mg/L) à l’aide du modèle (Éq. 4) aux stations Hurons et RSC2.
Comparison of the cycles in oxygen concentration (mg/L) measured and estimated with the model (Eq. 4) at the Hurons and RSC2 stations.
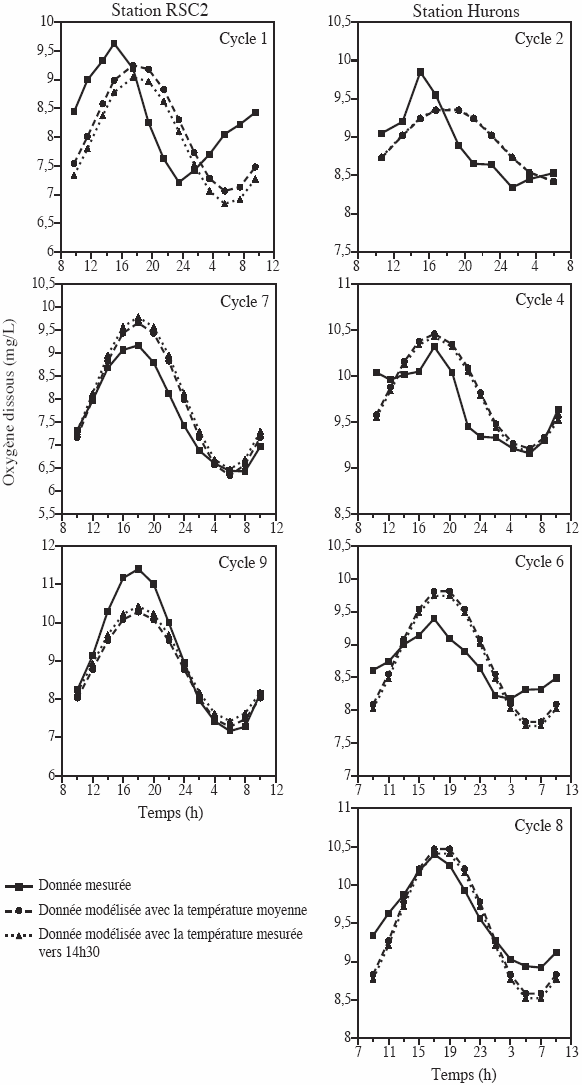
Figure 6
Relation entre la concentration en oxygène dissous estimée et mesurée (mg/L) pour les sept cycles diurnes aux stations Hurons et RSC2.
Relation between the measured and estimated dissolved oxygen concentration (mg/L) for the seven diurnal cycles at the Hurons and RSC2 stations.

L’application du modèle exige l’obtention de la température moyenne et de la concentration moyenne en nitrate sur 24 h. Toutefois, il est possible ici d’obtenir l’équivalent de la température moyenne par la mesure ponctuelle de la température entre 12 h et 16 h (en moyenne à 14 h 30). Les courbes modélisées avec la température mesurée vers 14 h 30 sont en effet presqu’identiques à celles obtenues avec la température moyenne (Figure 5). La régression entre les données modélisées avec la température mesurée vers 14 h 30 et les données mesurées (r2 = 0,75, p < 0,0001) est de plus similaire à la régression entre les données modélisées avec la température moyenne et les données mesurées (r2 = 0,77, p < 0,0001(Figure 6)). La concentration moyenne en nitrate peut aussi être remplacée par une mesure ponctuelle de la concentration en nitrate durant le jour car cette variable demeure stable durant 24 h.
L’adéquation entre les valeurs modélisées et les données mesurées a également été évaluée à l’aide du biais (absolu et relatif) et de la racine carrée de l’erreur quadratique moyenne (RCEQM). Ces critères de performance affichent des valeurs presque identiques pour les deux modèles (Tableau 2) tout en indiquant une qualité légèrement supérieure dans les résultats du modèle général basé sur la température moyenne. Les deux modèles donnent donc des performances pratiquement équivalentes pour estimer la variation diurne de l’oxygène. Le biais absolu et relatif s’avère très faible pour les deux modèles, ce qui indique que la fonction choisie induit un biais négligeable dans l’estimation de l’oxygène. Par ailleurs, l’erreur quadratique moyenne (˜ 0,5 mg/L), une fois comparée avec la moyenne des valeurs observées (˜ 9 mg/L), s’avère faible, ce qui indique une bonne performance pour les deux modèles utilisés.
Tableau 2
Biais (absolu et relatif) et racine carrée de l’erreur quadratique moyenne (RCEQM) entre les valeurs modélisées et les données mesurées.
Bias (absolute and relative) and root-mean-square error (RCEQM) between modelled values and measured data.
Critère de performance |
Modèle utilisant la température moyenne |
Modèle utilisant la température à 14 h 30 |
|---|---|---|
Biais absolu (mg/L) |
0,04 |
0,05 |
Biais relatif (%) |
0,22 |
0,33 |
RCEQM (mg/L) |
0,51 |
0,53 |
Les limites et la validation du modèle
Le modèle présenté constitue un exemple de la démarche à suivre afin de construire un modèle simple pour estimer la concentration moyenne en oxygène dissous en rivière et l’amplitude des variations. Le modèle a été développé pour la rivière Saint-Charles et des Hurons en période d’étiage estival. Toutefois, il ne pourrait être appliqué avant de compléter la validation à l’aide d’une banque de données autres que celles utilisées pour la calibration. Il demeure néanmoins que la présente étude démontre clairement que la méthode proposée est performante et facile à suivre afin de modéliser les cycles diurnes de l’oxygène en rivière.
4.3 Discussion
4.3.1 Température et concentration en nitrate
Parmi l’ensemble des facteurs physicochimiques et biologiques qui interviennent dans la variabilité diurne de l’oxygène aux rivières Saint-Charles et des Hurons, la température est un des facteurs dominants. La température influence en effet à la fois la solubilité de l’oxygène dans l’eau et l’activité biologique (photosynthèse et respiration), agissant ainsi sur le bilan de la concentration en oxygène à court terme (BUSHONG et BACHMANN, 1989; BUTCHER et COVINGTON, 1995; LIVINGSTONE, 1991; SABATER et al., 2000).
La relation entre l’amplitude des cycles d’oxygène et la concentration en nitrate reflète en partie la nitrification résultant de l’apport d’azote ammoniacal dans le milieu. L’absence de système de traitement des eaux usées adéquat à la rivière des Hurons et les débordements d’égouts qui peuvent survenir en aval de la rivière Saint-Charles (RSC2) lors des pluies sont une source potentielle d’ammoniaque. Le processus de nitrification engendre une perte en oxygène et peut en effet avoir un impact sur la concentration moyenne en oxygène en rivière (CHAUDHURY et al., 1998). Donc, l’intensité de la nitrification est susceptible d’avoir un impact direct sur l’amplitude des variations de l’oxygène dissous. Il est par ailleurs possible que la relation entre l’amplitude des variations de l’oxygène et le nitrate soit une relation indirecte et que la relation directe serait avec le phosphore qui est généralement considéré comme l’élément nutritif limitant la croissance algale en eau douce. En effet, le phosphore total et les nitrites-nitrates partagent un modèle de répartition très semblable dans les rivières du Québec (PAINCHAUD, 1997). Les rejets d’eaux usées municipales et les sources agricoles (fumiers, engrais minéraux) constituent des apports considérables à la fois d’azote et de phosphore aux écosystèmes aquatiques. Toutefois, les données en phosphore obtenues dans la présente étude sont insuffisantes pour faire une régression avec l’amplitude en raison de l’absence de données de phosphore pour la première année d’échantillonnage.
4.3.2 Autres variables
En plus de la température et de la concentration en éléments nutritifs, d’autres variables sont susceptibles d’avoir un impact sur la concentration moyenne en oxygène et l’amplitude des variations. Ces variables pourraient alors être utilisées afin de remplacer les termes C et A dans l’équation (1). Les activités autotrophe et hétérotrophe sont deux facteurs importants dans la variabilité de la concentration en oxygène en rivière sur une base diurne. Une façon facile d’obtenir une estimation de l’activité autotrophe est de mesurer la biomasse algale (concentration en Chl a). La relation entre la concentration en Chl a phytoplanctonique et l’amplitude des cycles pour la rivière Saint-Charles et des Hurons n’est toutefois pas significative. Cependant, l’activité benthique autotrophe n’est pas considérée avec cette mesure alors qu’elle peut être responsable de fortes fluctuations en oxygène en milieu fluvial (SABATER et al., 2000). Il est possible que l’activité benthique autotrophe ait une influence sur le cycle diurne de l’oxygène dans la rivière Saint-Charles, particulièrement à la station RSC2 alors que les roches sises aux abord de la rivière étaient colonisées par le périphyton.
L’absence d’événement de vent important durant la période d’échantillonnage et les faibles vitesses du courant (0,03 à 0,15 m/s) font en sorte que la réaération (autre que celle causée par la température) est un facteur secondaire intervenant dans les fluctuations diurnes observées aux rivières Saint-Charles et des Hurons. La réaération devrait tout de même être considérée dans l’élaboration d’un modèle semblable à celui présenté ici car elle peut fortement influencer les fluctuations diurnes de l’oxygène (GUASCH et al., 1998; WILCOCK et al., 1995), et ce, particulièrement en période de crue. La réaération pourrait donc être calculée afin d’estimer les paramètres C et A, et ce, à l’aide de la mesure de la vitesse du courant, de la profondeur et de la pente en utilisant par exemple l’équation de THYSSEN et ERLANSEN (1987).
5. Conclusion
La présente étude démontre qu’il est possible de créer un modèle simple en comparaison avec les modèles de qualité de l’eau existants qui sont souvent complexes et impliquent la mesures de nombreux paramètres sur des cycles de 24 h. Le modèle proposé permet d’estimer la variabilité diurne de l’oxygène en milieu fluvial et ainsi d’éliminer le biais introduit par la prise de mesures ponctuelles. Grâce à ce type de modélisation, les études qui ont trait à l’évaluation de la qualité de l’eau peuvent estimer les fluctuations diurnes de l’oxygène dans une rivière donnée et ainsi obtenir une meilleure évaluation du niveau d’oxygénation. L’oxygène étant une variable clé pour le fonctionnement des écosystèmes aquatiques, ceci peut avoir des implications majeures dans un cadre décisionnel. En effet, les interventions d’assainissement des eaux visent souvent à protéger ou à restaurer l’état d’oxygénation des plans d’eau. Les objectifs environnementaux de rejet, et les choix techniques pour les atteindre, doivent être basés sur une connaissance exacte et non biaisée de l’état réel d’oxygénation du milieu récepteur.
La généralisation du modèle utilisé dans la présente étude implique son application éventuelle à d’autres rivières, notamment celles soumises à de fortes influences anthropiques et susceptibles de présenter de grandes variations diurnes des teneurs en oxygène. L’application du modèle devra également être étendue à d’autres saisons. Le modèle pourra ainsi devenir un outil important pour le diagnostic et la gestion de la qualité de l’eau des rivières.
Parties annexes
Remerciements
Cette étude a été financée par une subvention du Programme d’aide à la recherche et au développement du ministère de l’Environnement du Québec (PARDE). Nous tenons à remercier David Booth de l’Université du Québec à Rimouski pour son aide précieuse dans l’élaboration du modèle sinusoïdal, ainsi que Richard Turcotte du Centre d’expertise hydrique du Québec (Ministère du Développement durable, de l’Environnement et des Parcs) dans le calcul des critères de performance du modèle. Nous remercions également la rédaction et les réviseurs de la Revue pour leurs suggestions constructives.
Références bibliographiques
- AMBROSE R.B.JR., WOOL T.A., CONNOLLY J.P. et R.W.SCHANZ, 1988. WASP4, A hydrodynamic and water quality model - model theory, user’s manual, and programmer’s guide. U.S. Environmental Protection Agency report. Athens, Georgia. EPA/600/3-87/039.
- ANTOINE Y.W.S. et D. BOWKER, 1996. Hydrological and environmental characteristics of the River Taff, South Wales, U.K. Limnologica, 26, 217-233.
- BACHMANN R.W., KORTGE K.J. et T.E. ROBERTSON, 1988. Primary production in a small agricultural stream. Verh. Int. Ver. Theor. Angew. Limnol., 23, 1179-1182.
- BARILLIER A., GARNIER J. et M. COSTE, 1993. Experimental reservoir water release: impact on the water quality on a river 60 km downstream (upper Seine River, France). Water Res., 27, 635-643.
- BENGEN D., LIM P. et A. BELAUD, 1992. Qualité des eaux de trois bras morts de la Garonne : variabilité spatio-temporelle. Rev. Sci. Eau, 5, 131-156.
- BILLEN G., GARNIER J., SERVAIS P., BRION N., FICHT A., EVEN S., BERTHE T. et M. POULIN, 1999. L’oxygène : un témoin du fonctionnement microbiologique. Programme scientifique Seine-Aval, vol. 5. Université de Rouen. Rouen.
- BOWIE G. L., MILLS W. B., PORCELLA D. B., CAMPBELL C. L., PAGENKOPF J.R., RUPP G.L., JOHNSON K.M., CHAN P.W.H. et S.A. GHERINI, 1985. Rates, constants, and kinetics formulations in surface water quality modeling. Environmental Research Laboratory, Office of Research and Development, U.S. Environmental Protection Agency. Athens, Georgia. EPA/600/3-85/040.
- BREZONIK P.L., 1994. Chemical kinetics and process dynamics in aquatic systems. Lewis Publishers, Boca Raton, 784 p.
- BROWN L.C. et J. BARNWELL, 1987. The enhanced stream water quality models QUAL2W and QUAL2E-UNCAS: documentation and user manual. Environmental Research Laboratory, Office of Research and Development, U.S. Environmental Protection Agency. Athens, Georgia. EPA/600/3-87/007.
- BUSHONG S.J. et R.W. BACHMANN, 1989. In situ nutrient enrichment experiments with periphyton in agricultural streams. Hydrobiol., 178, 1-10.
- BUTCHER J.B. et S. COVINGTON, 1995. Dissolved-oxygen analysis with temperature dependence. J. Environ. Eng., 121, 756-759.
- CHAPRA S.C., 1997. Surface water-quality modeling. McGraw-Hill, New York, 784 p.
- CHAPRA S.C. et D.M. DI TORO, 1991. Delta method for estimating primary production, respiration, and reaeration in streams. J. Envir. Eng., 117, 640-655.
- CHAUDHURY R.R., SOBRINHO J.A.H., WRIGHT R.M. et M. SREENIVAS, 1998. Dissolved oxygen modeling of the Blackstone River (northeastern United States). Water Res., 32, 2400-2412.
- CHESTERIKOFF A., GARBAN B. et D. OLLIVON, 1991. Daily rhythms in the River Seine: relative impacts of natural and anthropogenic factors. Water Res., 25, 1523-1528.
- DUNNETTE D.A. et R.M. AVEDOVECH, 1983. Effect of an industrial ammonia discharge on the dissolved oxygen regime of the Willamette River, Oregon. Water Res., 17, 997-1007.
- EL-SHAARAWI A.H., MAUL A. et B.G. BROWNLEE, 1988. Modélisation des fluctuations de l’oxygène dissous dans les cours d’eau par analyse de régression non linéaire. Direction générale des eaux intérieures, Institut National de Recherche sur les Eaux, Centre Canadien des Eaux Intérieures. Burlington, Ontario. Étude No 163, 9 p.
- ERDMANN J.B., 1979a. Systematic diurnal curve analysis. J. Water Pollut. Contr. Fed., 15, 78-86.
- ERDMANN J.B., 1979b. Simplified diurnal curve analysis. J. Envir. Eng., 105, 1063-1074.
- FALCONER R.A., 1992. Water Quality Modelling. Ashgate, Brookfield, 139 p.
- FILLION S., BLANCHET,D., BEAULIEU B., BERTHOD C., THÉRIAULT A. et C. TREMBLAY, 1987. Rivière Saint-Charles : contexte évolutif et potentiel de développement. Service de l’urbanisme, Ville de Québec, 43 p.
- GARNIER J., SERVAIS P., BILLEN G., AKOPIAN M. et N. BRION, 2001. Lower Seine River and estuary (France) carbon and oxygen budgets during low flow. Estuaries, 24, 964-976.
- GERARDIN V. et Y. LACHANCE, 1997. Vers une gestion intégrée des bassins versants. Atlas du cadre écologique de référence du bassin versant de la rivière Saint-Charles, Québec, Canada. Ministère de l’Environnement et de la Faune du Québec. Ministère de l’Environnement du Canada.
- GOWDA T.P.H., 1983. Modelling nitrification effects on the dissolved oxygen regime of the Speed River. Water Res., 17, 1917-1927.
- GUASCH H., ARMENGOL J., MART E. et S. SABATER, 1998. Diurnal variation in dissolved oxygen and carbon dioxide in two low-order streams. Water Res., 32, 1067‑1074.
- GULLIVER J.S. et H.G. STEFAN, 1984a. Stream productivity analysis with DORM-I, development of a computational model. Water Res., 18, 1569-1576.
- GULLIVER J.S. et H.G. STEFAN, 1984b. Stream productivity analysis with DORM-II, parameter estimation and sensitivity. Water Res., 18, 1577-1588.
- HAMMER M.J. et K.A. MacKICHAN, 1980. Hydrology and Quality of Water Resources. John Wiley & Sons, New York, 486 p.
- HÉBERT S., 1995. Qualité des eaux du bassin de la rivière Saint-Charles, 1979-1995. Ministère de l’Environnement et de la Faune, Québec. Rapport No QE-101, 41 p.
- HORNE A.J. et C.R. GOLDMAN, 1994. Limnology. 2e édition. McGraw-Hill, New York, 576 p.
- HOWARTH R. W., MARINO R., GARRITT R. et D. SHERMAN, 1992. Ecosystem respiration and organic carbon processing in a large, tidally influenced river: the Hudson River. Biogeochem., 16, 83-102.
- JACOBS L.A., VON GUNTEN H.R., KEIL R. et M. KUSLYS, 1988. Geochemical changes along a river-groundwater infiltration flow path: Glattfelden, Switzerland. Geochim. Cosmochim. A., 52, 2693-2706.
- KAENEL B.R., BUEHRER H. et U. UEHLINGER, 2000. Effects of aquatic plant management on stream metabolism and oxygen balance in streams. Freshwater Biol., 45, 85‑95.
- KELLY M.G., HORNBERGER G.M. et B.J. COSBY, 1974. Continuous automated measurement of rates of photosynthesis and respiration in an undisturbed river community. Limnol. Oceanogr., 19, 305-312.
- LAANE R.W.P.M., GIESKES W.W.C., KRAAY G.W. et A. EVERSDIJK, 1985. Oxygen consumption from natural waters by photo-oxidizing processes. Netherlands J. Sea Res., 19, 125-128.
- LAVALLÉE P., 1983. Rapport technique, rivière Saint-Charles. Ministère de l’Environnement du Québec, Direction de l’assainissement des eaux, Québec, 24 p.
- LÉGARÉ S., 1998. Dynamique de l’oxygène en lac et en rivière dans le bassin versant de la rivière Saint-Charles. Mémoire Maîtrise, Université Laval, Sainte-Foy, 146 p.
- LIVINGSTONE D.M., 1991. The diel oxygen cycle in three subalpine Swiss streams. Arch. Hydrobiol., 120, 457-479.
- MARZOLF E.R., MULHOLLAND P.J. et A.D. STEINMAN, 1994. Improvements to the diurnal upstream-downstream dissolved oxygen change technique of determining whole-stream metabolism in small streams. Can. J. Fish. Aquat. Sci., 51, 1591-1599.
- McCUTCHEON S.C., 1989. Volume I. Transport and Surface Exchange in Rivers. CRC Press, Las Vegas, NV, 344 p.
- MORIN G., CLUIS D., COUILLARD D., JONES H.G. et J.M. GAUTHIER, 1984. Modélisation de l’oxygène dissous et de la demande biochimique en oxygène à l’aide du modèle quantité-qualité CEQUEAU. Université du Québec, Institut national de la recherche scientifique, INRS-Eau. Sainte-Foy. Rapport No 155, 107 p.
- MÜLLER J. et G. WEISE, 1987. Oxygen budget of a river rich in submerged macrophytes (River Zschopau in the south of the GDR). Int. Rev. Gesamten Hydrobiol., 72, 653-667.
- NEAL C., WATTS C., WILLIAMS R.J., NEAL M., HILL L. et T.H. WICKMAN, 2002. Diurnal and longer term patterns in carbon dioxide and calcite saturation for the River Kennet, south-eastern England. Sci. Total Environ., 282-283, 205-231.
- NING S.K., CHANG N.-B., YANG L., CHEN H.W. et H.Y. HSU, 2001. Assessing pollution prevention program by QUAL2E simulation analysis for the Kao-Ping River Basin, Taiwan. J. Envir. Manage., 61, 61-76.
- O’CONNOR D.J. et D.M. DI TORO, 1970. Photosynthesis and oxygen balance in streams. J. Sanit. Eng. Div. ASCE, 96, 547-571.
- ODUM H.T., 1956. Primary production in flowing waters. Limnol. Oceanogr., 1, 102-117.
- PAINCHAUD J., 1997. La qualité de l’eau des rivières du Québec. Nat. Can., 121, 38-43.
- PALMER, M.D., 2001. Water quality modeling. A guide to effective practice. The World Bank, Washington, D.C., 157 p.
- QUINN J.M. et P.N. McFARLANE, 1989. Epilithon and dissolved oxygen depletion in the Manawatu River, New Zealand: simple models and management implications. Water Res., 23, 825-832.
- RAUCH W. et P.A. VANROLLEGHEM, 1998. Modelling benthic activity in shallow eutrophic rivers. Wat. Sci. Tech., 37, 129-137.
- RICKERT D.A., 1984. Use of dissolved oxygen modeling results in the management of river quality. J. Water Pollut. Contr. Fed., 56, 94-101.
- SABATER S., ARMENGOL J., COMAS E., SABATER F., URRIZALQUI I. et I. URRUTIA, 2000. Algal biomass in a disturbed Altantic river: water quality relationships and environmental implications. Sci. Total Environ., 263, 185‑195.
- SCHURR J.M. et J. RUCHTII, 1977. Dynamics of O2 and CO2 exchange, photosynthesis, and respiration in rivers from time-delayed correlations with ideal sunlight. Limnol. Oceanogr., 22, 208-225.
- SEIDL M., SERVAIS P. et J.M. MOUCHEL, 1998. Organic matter transport and degradation in the River Seine (France) after a combined sewer overflow. Water Res., 32, 3569-3580.
- SIMONSEN J.F. et P. HARREMOËS, 1978. Oxygen and pH fluctuations in rivers. Water Res., 12, 477-489.
- STAHL B. et C.G UCHRIN, 1989. Dissolved oxygen dynamics in the mid-Passaic River (New Jersey, USA): Mathematical model. J. Environ. Sci. Health Part A Environ. Sci. Eng., A24, 175-193.
- TCHOBANOGLOUS G. et E.D. SCHROEDER, 1985. Water quality, characteristics, modeling, modification. Addison-Wesley Publising Company, Reading, UK, 768 p.
- THYSSEN N. et M. ERLANDSEN, 1987. Reaeration of oxygen in shallow, macrophyte rich streams: II. Relationship between the reaeration rate coefficent and hydraulic properties. Int. Rev. Gesamten Hydrobiol., 72, 575-597.
- UCHRIN C.G., AHLERT W.K., CRYAN A.P., GIGA J.V., HSUEH S.-F., JENQ T.-R., LEWIS T., PARK S.S. et B. STAHL, 1988. Dissolved oxygen dynamics in the mid-Passaic River (New Jersey, USA): late summer/early fall 1983-4 field studies. J. Environ. Sci. Health Part A Environ. Sci. and Eng., A23, 525-542.
- VAN ORDEN G.N. et C.G. UCHRIN, 1993. The study of dissolved oxygen dynamics in the Whippany River, New Jersey using the QUAL2E model. Ecol. Model., 70, 1-17.
- WETZEL R. G., 2001. Limnology, lakes and river ecosystems. 3e édition. Academic Press, San Diego, 1006 p.
- WETZEL R.G. et G.E. LIKENS, 2000. Limnological analysis. 2e édition. Springer-Verlag, New York, 429 p.
- WHITEHEAD P.G., WILLIAMS R.J. et D.R. LEWIS, 1997. Quality simulation along river systems (QUASAR): model theory and development. Sci. Total Environ., 194/195, 447-456.
- WILCOCK R.J., McBRIDE G.B., NAGELS J.W. et G.L. NORTHCOTT, 1995. Water quality in a polluted lowland stream with chronically depressed dissolved oxygen: Causes and effects. New Zealand J. Mar. Freshwater Res., 29, 277‑288.
- WILLIAMS R.J., WHITE C., HARROW M.L. et C. NEAL, 2000. Temporal and small-scale spatial variations of dissolved oxygen in the Rivers Thames, Pang and Kennet, UK. Sci. Total Environ., 251/252, 497-510.
Liste des figures
Figure 1
Localisation des stations d’échantillonnage dans le bassin versant des rivières Saint-Charles et des Hurons (modifié de LÉGARÉ 1998).
Location of the sampling stations in the watershed of the St. Charles and des Hurons rivers (modified from LÉGARÉ 1998).
Figure 2
Cycles de la concentration en oxygène dissous (mg/L), du taux de saturation en oxygène (%) et de la température (oC) dans les rivières Saint-Charles (RSC1 (a), RSC2 (b)) et des Hurons (c).
Cycles of dissolved oxygen concentration (mg/L), oxygen saturation (%) and temperature (oC) in the St. Charles river (RSC1 (a), RSC2 (b)) and des Hurons river (c).
Figure 3
Relation entre la concentration moyenne en oxygène (mg/L) et la température moyenne (oC) sur 24 heures des cycles diurnes mesurés aux stations Hurons et RSC2.
Relationship between the mean oxygen concentration (mg/L) and temperature (oC) over 24 hours of the diurnal cycles measured at the Hurons and RSC2 stations.
Figure 4
Relation entre l’amplitude de variation des cycles de concentration en oxygène (mg/L) et la concentration moyenne en nitrate (mg/L) aux stations Hurons et RSC2.
Relationship between the amplitude of the cycles in oxygen concentration (mg/L) and mean nitrate concentration (mg/L) at the Hurons and RSC2 stations.
Figure 5
Comparaison des cycles de concentrations en oxygène mesurées et estimées (mg/L) à l’aide du modèle (Éq. 4) aux stations Hurons et RSC2.
Comparison of the cycles in oxygen concentration (mg/L) measured and estimated with the model (Eq. 4) at the Hurons and RSC2 stations.
Figure 6
Relation entre la concentration en oxygène dissous estimée et mesurée (mg/L) pour les sept cycles diurnes aux stations Hurons et RSC2.
Relation between the measured and estimated dissolved oxygen concentration (mg/L) for the seven diurnal cycles at the Hurons and RSC2 stations.
Liste des tableaux
Tableau 1
Cycles diurnes mesurés aux stations d’échantillonnage Hurons, RCS1 et RCS2.
Diurnal cycles measured at the Hurons, RSC1 and RSC2 stations.
Cycle |
Date |
Station |
Donnée et fréquence de mesure |
|---|---|---|---|
1 |
5 au 6 août 1996 |
RSC2 |
O2, T et luminosité mesurés à chaque 2 h Ntot, NO3 et Chl a mesurés à chaque 4 h |
2 |
7 au 8 août 1996 |
Hurons |
|
3 |
7 au 8 août 1996 |
RSC1 |
|
4 |
26 au 27 août 1996 |
Hurons |
|
5 |
26 au 27 août 1996 |
RSC1 |
|
6 |
17 au 18 juillet 1997 |
Hurons |
O2, T et luminosité mesurés à chaque 2 h Ptot, PTD, Ntot, NO3 et Chl a mesurés à chaque 4 h |
7 |
17 au 18 juillet 1997 |
RSC2 |
|
8 |
23 au 24 juillet 1997 |
Hurons |
|
9 |
23 au 24 juillet 1997 |
RSC2 |
O2 : oxygène dissous, T : température, Ptot : phosphore total, PTD : phosphore total dissous,
Ntot : azote total, NO3 : nitrate
Tableau 2
Biais (absolu et relatif) et racine carrée de l’erreur quadratique moyenne (RCEQM) entre les valeurs modélisées et les données mesurées.
Bias (absolute and relative) and root-mean-square error (RCEQM) between modelled values and measured data.
Critère de performance |
Modèle utilisant la température moyenne |
Modèle utilisant la température à 14 h 30 |
|---|---|---|
Biais absolu (mg/L) |
0,04 |
0,05 |
Biais relatif (%) |
0,22 |
0,33 |
RCEQM (mg/L) |
0,51 |
0,53 |