Résumés
Summary
The aims of this work were to study, for the first time, the succession of microbial communities (from viruses to ciliates) in the largest occidental European lake (Lake Geneva) and to perform two one-week in situ experiments in March-April (Exp1) and May (Exp2) 2004 in order to assess both small flagellate protozoan and virus-induced mortality of heterotrophic bacteria. Both nanoflagellates and viruses could be responsible for 31 to 42% of the total daily mortality of heterotrophic bacteria. In May (Exp2), viruses could explain up to 10% of the bacterial mortality whereas flagellates were responsible for 32% of the bacterioplankton removal. These results provide new evidence for the critical role played by viruses in the functioning of the microbial food webs and highlight the importance of further considering this biological compartment for a better understanding of the plankton ecology of Lake Geneva.
Keywords:
- lake,
- protists,
- viruses,
- bacteria,
- bacterial mortality,
- community dynamics,
- dilution method
Résumé
Il est aujourd’hui bien établi que l’on ne peut prétendre comprendre le rôle du vivant dans le fonctionnement des écosystèmes aquatiques sans répondre à un certain nombre de questions fondamentales telles que celles portant sur l’identité et l’abondance des organismes présents, leurs taux métaboliques et reproductifs, ou encore leurs fonctions précises dans l’écosystème. Dans les systèmes lacustres, l’omniprésence et le rôle-clé des processus microbiens ont été largement démontrés au cours des deux dernières décennies. Toutefois, les mécanismes de régulation ainsi que la diversité fonctionnelle des communautés microbiennes demeurent un sujet central de la recherche actuelle. Si nous avons commencé à accumuler de grandes quantités d’informations concernant le compartiment bactérien, il n’en est pas de même pour le compartiment contenant les plus petites et les plus abondantes entités biologiques de la colonne d’eau : les virus. L’importance qualitative, quantitative et fonctionnelle des virus bactériophages et leur impact dans le contrôle et le déclin des communautés bactériennes dans les écosystèmes lacustres sont en effet encore mal connus. On sait aujourd’hui que les virus interviennent dans les processus de perte (mortalité) qui affectent les communautés microbiennes, mais aussi dans la structure en taille, la composition et la régulation de la diversité des peuplements microbiens, dans le recyclage des nutriments inorganiques et de la redistribution de la matière organique.
Dans ce travail, nous avons étudié, pour la première fois, la dynamique des communautés microbiennes dans le plus grand lac naturel d’Europe occidental (le lac Léman) entre février et juin 2004. Le dénombrement des différents micro-organismes (effectué à huit profondeurs comprises entre 0 et 50 m) a été obtenu via la cytométyrie en flux et la microscopie à épifluorescence. Cette approche « écosystémique » nous a permis d’acquérir une image aussi précise que possible de la structure et de la dynamique de la communauté microbienne, composée des picocyanobactéries, des bactéries hétérotrophes, de petits eucaryotes flagellés et ciliés hétérotrophes et mixotrophes et enfin des virus. Typiquement, il a pu être mis en évidence des liens étroits entre certaines communautés, comme par exemple les bactéries et les virus du groupe VLP1 (r = 0,51; p < 0,05; n = 72), suggérés être, pour l’essentiel, représentatifs de la communauté des bactériophages.
Parallèlement au suivi limnologique des communautés, une approche expérimentale in situ, à différentes périodes de l’année, a aussi été utilisée, afin d’estimer la part relative de la prédation par les protozoaires flagellés et de l’impact des virus sur la mortalité bactérienne. La méthode mise en application ici est dérivée de la technique de dilution initialement proposée pour évaluer l’impact du broutage des organismes zooplanctoniques sur le phytoplancton. Un échantillon intégré entre 0 et 10 m de la colonne d’eau a été pré-filtré sur 100 µm puis 11 µm pour ne conserver que les communautés virales, bactériennes et protistes flagellés, soumises alors à des dilutions successives en présence (eau filtrée sur 0,2 µm) ou absence (eau ultrafiltrée sur 100 kDa) de la communauté des virus.
Ces expériences, réalisées en microcosmes de 0,5 litre, ont été conduites en mars - avril (Exp1) et en mai (Exp2) 2004. Nos résultats ont révélé que les nanoflagellés et les virus étaient responsables ensemble de 31 à 42 % de la mortalité bactérienne journalière. En mai, les virus expliquaient à eux seuls 10 % de la mortalité bactérienne, et les nanoflagellés 32 %. Ces données suggèrent que la lyse virale peut induire des changements dans l’importance relative des groupes fonctionnels de la chaîne alimentaire et qu’elle est d’une importance significative sur la mortalité du bactérioplancton.
Les expériences en microcosmes ont également permis de détecter une population virale VLP4, discriminable par cytométrie en flux des autres communautés de par ses caractéristiques de taille et de fluorescence. L’analyse précise des abondances de ce groupe et des autres communautés nous a alors permis de montrer une relation étroite entre les abondances des VLP4 et de petits eucaryotes pigmentés (r = 0,38; p < 0,05; n = 58), suggérant que ce groupe de virus pouvait être spécifique de ces cellules autotrophes.
L’ensemble des résultats acquis met en évidence l’importance des virus dans le fonctionnement du réseau trophique microbien et la nécessité de considérer désormais ce compartiment biologique pour mieux comprendre l’écologie planctonique du Léman.
Mots clefs:
- lacs,
- protozoaires,
- virus,
- bactéries,
- mortalité bactérienne,
- dynamique des communautés,
- méthode de dilution
Corps de l’article
1. Introduction
It is only in the past 15 years that it has been realized how viruses are an important biological compartment in aquatic microbial food webs. In the 1970’s, TORELLA and MORITA (1979) demonstrated that viruses present in seawater were more abundant than previously reported (> 104 viral particles per millilitre). In later studies, other researchers corroborated this observation measuring viral abundances in a typical range of 109 to 1010 viral particles per litre in seawater or lake water (BORSHEIM et al., 1990; PAUL et al., 1991). Henceforth, high viral abundances have been observed worldwide, in seawaters (BERGH et al., 1989; PROCTOR and FUHRMAN, 1990), coastal waters (SUTTLE et al., 1990; PAUL et al., 1991) and freshwaters (KLUT and STOCKNER, 1990; BERGH et al., 1989); in marine (PAUL et al., 1993) and freshwater sediments (MARANGER and BIRD, 1995); and in the annual ice of the Polar Sea as well (MARANGER et al., 1994).
Because they are very abundant and have a wide taxonomic and functional diversity, microorganisms play a predominant role in matter and energy fluxes within aquatic ecosystems. Viruses have been shown to be important controlling agents of planktonic community composition, diversity and succession, and to play a key role in cell mortality and nutrient cycles (BERGH et al., 1989; SUTTLE, 1994; MARANGER and BIRD, 1995; FUHRMAN, 1999; WOMMACK and COLWELL, 2000; WEINBAUER and RASSOULZADEGAN, 2004). Of additional ecological significance, the ability of aquatic viruses to transfer genetic material has been demonstrated (CHIURA, 1997; CLOKIE et al., 2003). It is noteworthy that the large majority of aquatic viral ecological studies have been carried out in seawater (see FUHRMAN, 1999; WOMMACK and COLWELL, 2000; WEINBAUER, 2004; WEINBAUER and RASSOULZADEGAN, 2004) and there is only one review dealing with freshwater viral ecology (SIME-NGANDO et al., 2003).
It is important to understand the mechanisms responsible for the mortality of aquatic bacteria and phytoplankton. Indeed, different ways of microbial mortality yield different pathways of organic matter flow and oxidation in the ecosystem. In marine ecosystems, for example, SUTTLE and CHEN (1992) estimated that 4‑13% of the bacterial community would have to be infected daily and that 8‑26% of the bacterial mortality could be explained by viral lysis in marine ecosystems. FUHRMAN and NOBLE (1995) demonstrated that viruses can have a significant impact on the cycling of organic matter in the pelagic microbial food web since they are capable of contributing to as much as half of the heterotrophic bacterial mortality in coastal waters. In fresh water ecosystems, viruses have also been shown to play a crucial role in bacterial mortality, with daily bacterial removal reaching on occasion up to 97% (WEINBAUER and HOFLE 1998). More generally, the impact of viruses is very similar to what has been recorded in marine research with bacterioplankton mortality varying between 10 and 60% (BETTAREL et al., 2003a,b; 2004; FISHER and VELMIROV, 2002; SIMEK et al., 2001; JACQUET et al., 2005).
The aim of this study was to assess the relative impact of viruses and flagellate protozoa on the fate of heterotrophic bacterial communities at two different periods of the year in Lake Geneva. Moreover, the dynamics of the different biological compartments of the microbial loop, including viruses, heterotrophic bacteria, picocyanobacteria, pigmented and heterotrophic flagellates and ciliates was also monitored between February and June 2004. This survey allowed us to select, two different periods of the year between February and June to perform our “mortality experiments”, on the basis of clearly different population concentrations and dynamics.
2. Materials and Methods
2.1 Study site
Lake Geneva (46°27’N, 06°32’E) is the largest natural occidental European lake, and is located in the eastern part of France. It is an elongated (72.3 and 13.8 km in length and width respectively) and west-east oriented lake, with an area of 580 km2, a total volume of 89 x 109 m3, maximum and average depths of 310 m and 153 m respectively, and a water residence time of approximately 11.4 years. The catchment area is approximately 7,395 km2, with an average altitude of 372 m. It is a mesotrophic lake according to OECD (Organisation for Economic Co-operation and Development) criteria.
2.2 Community dynamics
To assess the dynamics of the microbial community (i.e., viruses, heterotrophic bacteria, picocyanobacteria, pigmented picoeukaryotes, nanoflagellates and ciliates) we used flow cytometry (FCM) and epifluorescence microscopy (EFM). Samples were taken at the reference station of the lake named SHL2, which corresponds to the maximum depth of the ecosystem. Eight depths (2.5; 7.5; 10; 15; 20; 25; 30 and 50 m) were monitored once or twice a month between February and June 2004.
2.3 Experimental set-up to estimate the impact of viruses and flagellate protozoa on the bacterial community
A dilution approach initially introduced by LANDRY and HASSET (1982) and refined by LANDRY et al. (1995) has been routinely used as a field technique to quantify grazing of phytoplankton by micro-zooplankton and to estimate phytoplankton growth rates (CAMPBELL and CARPENTER, 1986; WEISSE and SCHEFFEL-MOSER, 1990; LANDRY et al., 1993; CALBET and LANDRY, 2004). We used a modification of this dilution technique to assess the virus-induced mortality of heterotrophic bacteria in Lake Geneva, which was successfully tested on Lake Bourget the year before (JACQUET et al., 2005). Briefly, the technique consists of incubations of water samples after dilution to different levels of the original sample to reduce the abundance of algal and/or bacterial predators and thus making contact and interactions between predator and prey species less likely to occur with dilution. The net growth rate of the prey can therefore be expected to be highest in the most diluted fractions, provided that there is no nutrient limitation. The method allows an estimate of the grazing impact of zooplankton on phyto- or bacterioplankton. By using 0.02‑µm filtered water to dilute the sample, it is thus possible to focus on the viral effect. In this study, the impact of flagellates using the dilution method was also compared with the assessment of flagellate grazing using 0.5‑µm tracer particles (details can be found in JACQUET et al., 2005).
In the present study, two similar experiments were conducted in March-April (30/03 – 09/04: Exp1) and in May (11/05 – 18/05: Exp2) 2004, which corresponded to distinct periods in terms of water column stratification and of the microbial planktonic dynamics (see below). For each period, an integrated > 60-L sample was taken in the 0‑10 m surface layer of the reference station. The water sample was first filtered through a series of 100- and 11‑µm mesh filters (NYCOM, Buisine, France), and then sequentially filtrated through a 11‑µm filter, a 2‑µm filter and a 0.2‑µm filter (Nuclepore, Whatman) twice for the latter in order to eliminate all the bacterivores (i.e., nanoflagellates and ciliates) and bacteria. A fraction of the 0.2 µm-filtered water was subjected to tangential ultra-filtration using a mini-Ultrasette with a 100 kDa cut-off membrane (Vivaflow, Vivasciences) in order to eliminate all biological entities (including viruses). The purity of the water was checked using both TEM and FCM (data not shown). The 11‑µm fraction was mixed in order to obtain fractions of the initial 0.2 µm or 100 kDa-filtered whole water of about 20, 40, 70 and 100% (also referred to as the control for the latter hereafter). A control bottle containing 0% of the original 0.2 µm‑ or 100 kDa‑filtered water was also prepared for each experiment in order to confirm the absence of plankton contamination from the surrounding environment throughout the experiments (data not shown). For each level of dilution, triplicates were prepared in acid-washed, water‑rinsed and autoclaved 500 mL polycarbonate bottles (Nalgene, Bioblock). Immediately after being prepared, the bottles were attached in the harbour at the Thonon hydrobiological station and then incubated in situ at a depth of 1 m. Population dynamics were monitored during one week, assuming that there were no nutrient limitation and/or confinement effects during incubation. All bottles were sampled daily in the morning and samples were subsequently prepared for FCM analysis.
2.4 FCM and EFM counts
We used a FACSCalibur flow cytometer (Becton Dickinson) equipped with an air-cooled laser providing 15 mW at 488 nm with the standard filter set-up. One‑mL samples were filtered through 60 µm. Autotrophs were analyzed without adding any fixative or dye. Fluorescent microbeads (Molecular Probes Inc.) of 1 µm in diameter were added to each sample as an internal standard. Viruses and bacteria were fixed with 0.25% glutaraldehyde (final concentration, Sigma). Briefly, for heterotrophic bacteria, samples were diluted with 0.02 µm-filtered water from the lake while for viruses, samples were diluted with 0.02 µm-filtered TE (Tris-EDTA, pH = 8) buffer, and then heated for 10 minutes at 75°C. Samples were stained with SYBR Green I (1/10,000 final dilution of stock supplied by manufacturer) for 15 minutes in the dark (more details can be found in MARIE et al., 1999).
Glutaraldehyde (1% final concentration) was used to fix the flagellates. Samples were filtered (pressure < 100 mm Hg) onto polycarbonate membranes (diameter: 25 mm, pore size: 0.8 µm), then stained with primulin (modified from CARON, 1983) and stored for no more than a few days at ‑20°C until analysis. Slides were examined under UV light to count the heterotrophic nanoflagellates, and under blue light to count the autotrophic flagellates at a 1,250 x magnification. Green light (545 nm) was useful to display phycoerythrin-containing flagellates (Figure 1).
Figure 1
Typical flow cytometric dot plots obtained during the study of the in situ microbial population dynamics (A, B) and during the experiments examining the impact of viruses and flagellated protozoa on bacteria (C, D). The flow cytometric signatures of the designated Virus-Like Populations (VLP) referred to as VLP1, VLP2, VLP3 and VLP4, of the picocyanobacteria, the small eukaryotes and the heterotrophic bacteria are indicated. The VLP1 group likely corresponds to bacteriophages. The VLP2 and VLP3 groups might be specific to either cyanobacteria of bacteria (Jacquet, personal observations). The VLP4 is likely to be specific to an eukaryotic alga.
Résultats typiques obtenus par cytométrie en flux pendant l’étude de la dynamique des populations microbiennes in situ (A, B), ainsi que pour les expériences où nous avons examiné l’impact des virus et des protozoaires flagellés sur les bactéries (C, D). Les signatures en cytométrie des populations ressemblant aux virus (VLP1, VLP2, VLP3 et VLP 4) et des populations de picocyanobactéries, de petits eucaryotes et de bactéries hétérotrophes sont indiquées. Le groupement VLP1 correspond probablement aux bactériophages. Les groupements VLP2 et VLP3 seraient peut-être spécifiques aux cyanobactéries et/ou aux bactéries (Jacquet, communication personnelle). Le groupement VLP4 est probablement spécifique à une algue eucaryotique.

Ciliates were preserved with mercuric chloride (2.5%), identified and counted according to SIME-NGANDO et al. (1991). The sedimentation of samples was done during 48 h in an UTHERMOL column, and their observation was carried out with an inverted microscope at a 1,250 x magnification.
Figure 1 was performed using WinMDI (J. TROTTER, SCRIPPS, USA). Figures 2 to 4 were created in SIGMAPLOT (SYSTAT Software Inc.). The same software was used to carry out simple model 1 statistical analysis.
Figure 2
In situ population dynamics between February and June 2004. Flow cytometric data are shown in panels A to D. Epifluorescence microscopic data are shown in panels E to H.
Dynamique in situ des populations entre février et juin 2004. Les données issues de la cytométrie en flux figurent dans les panneaux A – D. Les données obtenues en épifluorescence sont présentées dans les panneaux E – H.
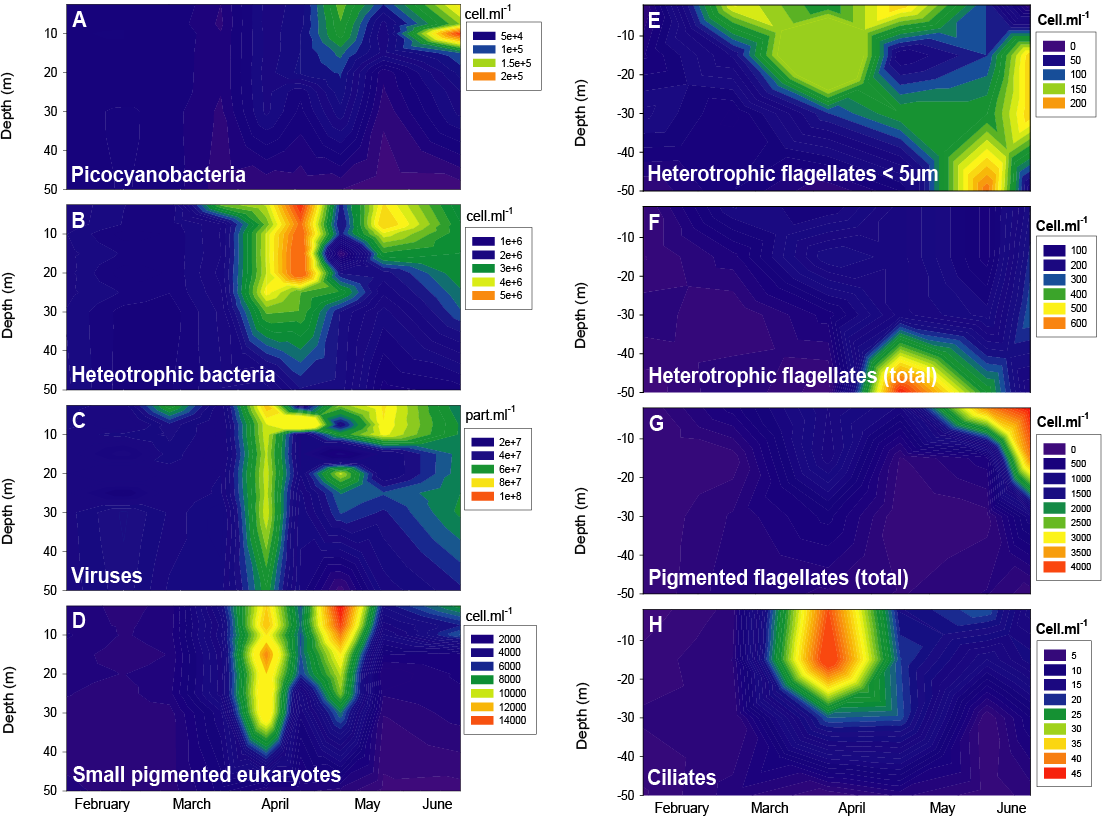
Figure 3
Dilution plots and analysis when < 100 KDa and < 0.2 µm filtered lake water were used to dilute the microbial samples (i.e. < 11 µm) during experiment 1 (A) and 2 (B).
Graphiques de dilution et analyse lorsque de l’eau de lac filtrée (< 100 kDa ou < 0,2 µm) est utilisée pour diluer les échantillons microbiens (i.e., < 11 µm) pendant l’expérience 1 (A) et 2 (B).

Figure 4
VLP4 population dynamics during the first experiment (A) and VLP4 concentration plotted against the pigmented eukaryotic community concentration for a dilution series set up with 0.2 µm and 100 KDa diluents (B).
Dynamique de la population VLP4 pendant la première expérience (A), et relation entre la concentration du groupement VPL4 et celle de la communauté eucaryotique pigmentée pour une série de dilutions obtenue avec l’eau filtrée (< 0.2 µm ou < 100 kDa).

3. Results
3.1 In situ microbial community dynamics
There was a clear succession of the different communities sampled between February and June 2004 (Figure 2). The first groups for which the concentrations increased significantly were those of the small pigmented and heterotrophic eukaryotes for which both EFM and FCM revealed different groups (see below). For the small pigmented eukaryotes detected with FCM, two peaks were recorded on April 15 and May 10, reaching up to 14 x 103 cells•mL‑1 between 0 and 15 m (Figure 2D, E). The heterotrophic bacteria dominated in April with a peak at the end of the month (Figure 2B), and were followed by picocyanobacteria (Figure 2A), whose biomass culminated between mid-April and the end of May with a peak at the beginning of May. Whereas the high concentrations of the bacterial communities (4‑6 x 106 cells•mL‑1) stretched over the first 25 meters, those of the picocyanobacteria (10‑14 x 103 cells•mL‑1) stretched over only the 15‑20 first meters. The highest concentration found for picocyanobacteria was recorded at the end of June at 10 m depth with 2.2 x 105 cells•mL‑1. Within the bacterial community, we could detect, at times, two distinct groups that had been observed previously and referred to as low‑ and high DNA content bacteria (ROBERTSON and BUTTON, 1989; LI et al., 1995; MARIE et al., 1996; LEBARON et al., 2001; SERVAIS et al., 2003). However, this distinction was not always evident and we did not find a clear relationship between each of these two groups when detected with the other biological communities. The viruses, counted as a whole community, presented higher concentrations (4‑5 x 107 particles•mL‑1) in the water column in mid-April and the viral titre increased from the beginning to the end of May (4‑7 x 107 particles•mL‑1), reaching a maximum concentration of 8 x 107 particles•mL‑1 near the surface (Figure 2C). At least three distinct viral groups were observed during the field survey but their discrimination was not always possible. The heterotrophic flagellate abundance as determined with EFM (Figure 2E, F) fluctuated from 0.5 x 101 cells•mL‑1 (April, 50 m) to 6.9 x 102 cells•mL‑1 (May, 50 m). Heterotrophic flagellates smaller than 5 µm (Figure 2E), which are known to be the main bacterial grazers, increased in the epilimnion from March and their maximal abundance was recorded in April at 15 m (2.4 x 102 cells•mL‑1). These small flagellates could also present rather high abundances in the hypolimnion during June. Peaks for total heterotrophic flagellate abundance in the upper layers (0‑15 m) were observed during April (2.4 x 102 cells•mL‑1 at 15 m) and at the beginning of May (2.1 x 102 cells•mL‑1 at 2 and 15 m) while in the bottom layers (from 30 to 50 m), very low abundances were recorded until the middle of May. At that time, a peak of abundance was observed due to the presence of epiphytic flagellates (mainly choanoflagellates) fixed on phytoplanktonic cells, probably corresponding to the sedimenting cells. Pigmented flagellates had moderate concentrations during the first two months (Figure 2G), with the first increase in numbers occurring in the euphotic zone during April (1.1 x 103 cells•mL‑1 at 15 m depth), but a significant peak in abundance appeared only in June (4.3 x 103 cells•mL‑1 at 2 m depth). From February to June, the abundance of ciliates (Figure 2H) ranged from 3 cells•mL‑1 (February, 2 m) to 46 cells•mL‑1 (April, 15 m). The highest values were observed during April in the epilimnetic and metalimnetic layers concomitantly with the development of bacteria, picocyanobacteria, and small flagellates. During this spring increase, we observed mainly Oligotrichs, Haptorids, Colpods and some Scuticociliates while Prostomatids constituted a dominant group during the winter period. Oligotrichs were dominant and were mainly represented by Halteria and Strombidium, which are known to be important bacterial consumers in meso‑ to eutrophic freshwater systems, with a strong grazing impact on a wide prey size spectrum (JURGENS and SIMEK, 2000; KISAND and ZINGEL, 2000; SIMEK et al., 2000).
3.2 Relationships among the different microbial groups
There was a positive relationship between viral and heterotrophic bacterial communities (r = 0.51; p < 0.05; n = 72). The higher densities of ciliates in the epilimnion were consistent with the higher densities of resources in these layers (flagellates, small picoeukaryotes and bacteria), and significant correlations were found between the abundance of ciliates and the abundance of heterotrophic bacteria (r = 0.7; p < 0.05; n = 24), and with the abundance of pigmented eukaryotes (r = 0.81; p < 0.05; n = 24). No significant correlations were found among the other communities.
3.3 Estimates of the impact of viruses and flagellate protozoa on bacterial mortality
For the first in situ experiment estimating the impact of viruses and flagellated protozoa on bacteria, the experimental samples were characterized by relatively low abundances of the four microbial communities counted by FCM at t0: 2 x 103, 3 x 103, 3 x 106, and 3 x 107 cells•mL‑1 for picocyanobacteria, small pigmented eukaryotes, heterotrophic bacteria and viruses respectively. For the second experiment, higher concentrations of picocyanobacteria (12‑14 x 103 cells•mL‑1), eukaryotes (12‑14 x 103 cells•mL‑1), viruses (6‑7 x 107 particles•mL‑1) and heterotrophic bacteria (4 x 106 cells•mL‑1) were recorded. For the two experiments, we specifically determined the abundance of protozoan grazers in the water filtered through 11 µm. On March 30, the total abundance of flagellates (pigmented and heterotrophic) in the fraction < 11 µm was 248 cells•mL‑1. Heterotrophic flagellates (HF) largely dominated with 230 cells•mL‑1. At the beginning of the second experiment (May 11) the abundance of flagellates was significantly higher, with a total number of 708 cells•mL‑1 (heterotrophic + pigmented cells). Once again, heterotrophic flagellates were dominant with an abundance reaching 574 cells•mL‑1, representing 81% of the total flagellate community. The differences in abundance and structure of the flagellated grazers at the beginning of the two experiments suggested that their impact in the epilimnion was different in March and May. From one experiment to the other, we switched from a moderate abundance of flagellates to a rather high abundance. During the first experiment, small flagellates (size 2‑4 µm) were dominant, accounting for 84% of the total abundance of heterotrophic flagellates, whereas in May heterotrophic flagellates were more diversified. Typically, small uniflagellates were well represented (34% of total HF), but cryptomonad forms such as Katablepharis dominated (45% of total HF) and Chrysomonad forms as Spumella represented 12% of the total HF abundance.
The results which follow relate to lake water initially collected and filtered through 11 µm and thus containing only viruses, bacteria and flagellate protozoa. Initially, we checked that the dilution with ultra pure water (< 100 KDa) yielded a clear gradient in viral concentrations (data not shown). In the two dilution experiments conducted in March-April (Exp1) and in May (Exp2), significant relationships were found between the level of dilution and the apparent growth rate of the heterotrophic bacterial community. We estimated total mortality rates (attributable to both viruses and flagellate protozoa) to be 31% d‑1 for Exp1 (Figure 3A) and 42% d‑1 for Exp2 (Figure 3B). During the second experiment, by using virus-free diluent (< 100 kDa) in the dilution experiments in parallel with the conventional 0.2 µm dilution series, the impact of flagellates and viruses could be separated and was calculated to be equivalent to daily bacterial removal rates of 32% d‑1 and 10% d‑1 respectively.
3.4 VLP4: a particular population of viruses?
The FCM analysis allowed us to observe four different populations of viruses (Figure 1C, D), with the onset of a particular viral population named VLP4 for «Virus-Like-Particles group 4» (Figure 1D) distinguished by its typical fluorescence and size characteristics. The VLP4 population emerged between the 4th and the 6th day of the first experiment with a concentration increasing from about 3 x 104 particles•mL‑1 up to 3 x 106 particles•mL‑1. Subsequently, the VLP4 concentration remained quasi-constant until the end of the experiment (at the 9th day). This suggests that these viruses infected their hosts between the 1st and 4th day of the experiment. During the second in situ experiment, we also detected the presence of VLP4. This population was present in considerable concentrations (> 1 x 106 particles•mL‑1 in the undiluted < 11 µm raw samples) from the very beginning of Exp2. The concentrations of these viral particles were multiplied by 2.2 between the 5th and the last day of the experiment, ranging from 7.5 x 105 to 1.8 x 106 particles•mL‑1 in the undiluted water fraction, and from 1 x 105 to 1.1 x 106 particles•mL‑1 in the dilution containing 20% of the raw samples (i.e. < 11 µm). Although this population was present from the beginning of the second experiment, its concentrations did not reach those observed in the first experiment (1.9 x 106 particles•mL‑1 in Exp2 vs 3 x 106 in Exp1). The concentrations of VLP4 were not correlated to those of the prokaryotic community (i.e. heterotrophic bacteria or picocyanobacteria) but to those of the small pigmented eukaryotes. The correlations obtained in experimental samples diluted with < 100 kDa and < 0.2 filtered water were both significant (rα/2 = 0.38, n = 58, p < 0.05). We counted approximately 50 VLP4 per eukaryote.
4. Discussion
No data concerning the dynamics of the entire main microbial community, characteristic of the microbial loop in Lake Geneva, have been published to date. In addition, this is the first study dealing with natural aquatic viruses of Lake Geneva, both in terms of their dynamics at different periods of the year and their potential functional role in bacterial mortality. To date, there have been few investigations of seasonal fluctuations in the abundance of these particles in large natural freshwater ecosystems (HENNES and SIMON, 1995; TAPPER and HICKS, 1998; HOFFER and SOMMARUGA, 2001; FISHER and VELMIROV, 2002; BETTAREL et al., 2003b; VREDE et al., 2003; BETTAREL et al., 2004). Moreover, predation by protozoa and parasitism by viruses were investigated simultaneously, a combination that has not often been accomplished in the past (FUHRMAN and NOBLE, 1995; GUIXA-BOIXAREU et al., 1996, 1999; WEINBAUER and HOFLE, 1998; PEDROS-ALIO et al., 2000; ALMEIDA et al., 2001; SIMEK et al., 2001; BETTAREL et al., 2003a, 2004; HORNAK et al., 2005; JACQUET et al., 2005).
The dynamics of the microbial communities were comparable to those found in many other freshwater ecosystems in which, after a winter period characterized by relatively low concentrations of the different groups, there is a traditional spring proliferation between 0 and 20 meters, supported by increases in temperature and luminosity, and by the presence of nutrients in surface waters (WEISSE, 1993; CALLIERI and STOCKNER, 2002; BETTAREL et al., 2003a). By examining all the data together, we found a clear and positive relationship between bacterial and viral numbers (especially VLP1, cf. Fig. 1C), suggesting that these viruses were mainly bacteriophages (SIME-NGANDO et al., 2003; Weinbauer, 2004) and that a tight coupling existed between the two communities in Lake Geneva. However, the succession of the different populations, especially the HNF and the ciliates after the bacterial groups, also strongly suggested that these protists were an important factor in bacterial biomass regulation (JURGENS and SIMEK, 2000; KISAND and ZINGEL, 2000; SIMEK et al., 2000). The importance of flagellate predation was definitely confirmed with our two in situ experiments, which revealed that these small grazers could be responsible for ~30% of the daily bacterial production removal. The evaluation of clearance rates for each flagellate taxon (using ingestion of fluorescent microbeads), allowed us to estimate that the potential flagellate bacterivory in the < 11 µm water sample represented 10% and 43% of daily bacterial removal, respectively in the first and the second experiments. Grazing rates of flagellates were estimated as the product of bacterial concentration, flagellate concentration and assessed as clearance rates for each taxon. Although we are aware that short-term measurements done with fluorescent particles (a few minutes) are quite difficult to extrapolate to daily impact, it appeared clearly that the impact of flagellate grazing on bacteria was much lower during the first experiment compared to the second one. During the experiment carried out in May, the estimation of grazing rates using fluorescent microbead methods could be compared to the result obtained with the dilution method. It appeared that the fluorescent microsphere technique led to the estimation of higher values in terms of flagellate impact on bacteria (43% vs. 32% with the dilution method), but still the range was comparable. This also means that the viral impact could be higher in April than May.
By comparison with the flagellate impact on bacteria, we found that viral-induced bacterial mortality was lower, as generally reported in the literature on pelagic systems. However, the impact of lysis in Lake Geneva seemed to be in the same range as the oligotrophic Lake Pavin (6‑16%) or the mesotrophic lake Bourget (9‑19%) but definitely lower than in the anoxic layer of the eutrophic Lake Plussee or the oligotrophic Lake Pavin (up to 97%) as reported by BETTAREL et al. (2003, 2004), WEINBAUER and HOFLE (1998) and SIME-NGANDO (personal communication). Thus, viruses were at least responsible for 10% of the daily bacterial removal and we can expect that this impact could be higher at other periods of the year, for example in summer when temperature increases (BETTAREL et al., 2003, 2004). We also estimated viral impact in surface waters only and it is likely that viral-induced bacterial mortality may change with increasing depth (BETTAREL et al., 2003; WEINBAUER and HOFLE 1998).
The existence of different viral groups observed with FCM has been observed in the past but to the best of our knowledge this is one of the rare reports where four different groups could be detected. This again emphasizes how powerful a technique like flow cytometry is for the detection, discrimination and precise quantification of different microbial communities. This is particularly true for the VLP4 population. Although we did not identify this population using transmission electron microscopy and molecular tools, we speculate that this viral group was probably linked to the pigmented eukaryote community. Indeed, during a field experiment in Norwegian coastal waters, JACQUET et al. (2002) revealed, on the basis of cytofluorimetric signature, the presence of a viral population that infects the photosynthetic eukaryote Emiliana huxleyi. Similarly, BRUSSAARD et al. (1999) described the characteristics of the cytofluorimetric signature of a viral population that infected the Prymnesiophyceae Phaeocystis pouchetii. Because the viral population VLP4 had exactly the same signature as that observed in these previous studies relating to phytoplanktonic eukaryotes, we decided to investigate the eukaryotic population in Exp2. According to the significant correlation we found between these two groups, and according to what is described above, it is likely that VLP4 population comprised viruses infecting one or several of the small, pigmented eukaryotes in our samples. Finally, all VLP4 particles passed through 0.45 µm polycarbonate filters and only a negligible fraction through 0.2 µm polycarbonate filters, suggesting that these particles were indeed viruses and not small bacteria. In the future, we will try to isolate this population of viruses for molecular characterization and for studying their ecological significance in Lake Geneva.
5. Conclusion
We are just beginning to study the natural viral dynamics and diversity of Lake Geneva and our first results are very promising. Using a modified approach of the dilution approach, we could address the question of the viral vs. protozoan impact on bacterial mortality. This is particularly critical given that bacterioplankton are one of the most important biological compartments at the base of the food web, responsible for the recycling of nutrients and the decomposition of organic matter in the pelagic zone of freshwater ecosystems, and the main food source for pelagic protists. To our knowledge, this dilution technique has only been used a few times (WILHELM et al., 2002; EVANS et al., 2003; JACQUET et al., 2005). Although this technique has some drawbacks (discussed in JACQUET et al., 2005), we strongly believe that it will be used more and more in the field of aquatic viral ecology to assess viral impacts on various planktonic populations or communities. Finally, we have shown that flow cytometry allows for rapid observation and discrimination of several viral groups of interest.
Parties annexes
Acknowledgements
We are grateful to J.C. HUSTACHE who helped in water sampling and to A. MILLERY and J.N. AVRILLIER who helped in protist counting. R. NOBLE is acknowledged for her critical reading and English correction of an initial version of the manuscript.
References
- Almeida M.A, Cunha M.A., Alcantara F., 2001. Loss of estuarine bacteria by viral infection and predation in microcosm conditions, Microb. Ecol., 42, 562-571.
- Bergh O., Borsheim K.Y., Bratbak G., Heldal M., 1989. High abundances of viruses found in aquatic environments, Nature, 340, 467-468.
- Bettarel Y., Amblard C., Sime-NGando T., Carrias J.F., Sargos D., Garabetian F., Lavandier P., 2003a. Viral lysis, flagellate grazing potential and bacterial production in Lake Pavin, Microb. Ecol., 45, 119-127.
- Bettarel Y., Sime-Ngando T., Amblard C., Carrias J.F., Portelli C., 2003b. Virioplankton and microbial communities in aquatic systems: a seasonal study in two lakes of differing trophy, Freshw. Biol., 48, 810‑ 822.
- Bettarel Y., Sime-NGando T., Amblard C., Dolan J., 2004. Viral activity in two contrasting lake ecosystems, Appl. Environ. Microb., 70, 2941-2951.
- Børsheim G., Bratbak G., Heldal M., 1990. Enumeration and biomass estimation of planktonic bacteria and viruses by transmission electron microscopy, Appl. Environ. Microb., 56, 352-356.
- Brussaard C.P.D., Thyrhaug R., Marie D., Bratbak G., 1999. Flow cytometric analyses of viral infection in two marine phytoplankton species, Micromonaspusilla (Prasinophyceae) and Phaeocystis pouchetii (Prymnesiophyceae), J. Phycol., 35, 941-948.
- Calbet A., Landry M.R., 2004. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems, Limnol. Oceanogr., 49, 51-57.
- Callieri C., Stockner J.G., 2002. Freshwater autotrophic picoplankton: a review, Limnology, 61, 1-14.
- Campbell L., Carpenter E.J., 1986. Estimating the grazing pressure of heterotrophic nanoplankton on Synechococcus spp. using the sea water dilution and selective inhibitor techniques, Mar. Ecol. Progr. Ser., 33, 121-129.
- Caron D.A., 1983. Technique for enumeration of heterotrophic and phototrophic nanoplankton, using epifluorescence microscopy, and comparison with other procedures, Appl. Environ. Microb., 46, 491-498.
- Chiura H.X., 1997. Generalized gene transfer by virus-like particles from marine bacteria, Aquat. Microb. Ecol., 13, 75‑83.
- Clokie M.R.J., Millard A.D., Wilson W.H., Mann N.H., 2003. Encapsidation of host DNA by bacteriophages infecting marine Synechococus strains, FEMS Microb. Ecol., 46, 349-352.
- Demuth J., Neve H., Witzel K., 1993. Direct electron microscopy study on the morphological diversity of bacteriophage populations in Lake Plusssee, Appl. Environ. Microb., 59, 3378-3384.
- Evans C., Archer S.D., Jacquet S., Wilson W.H., 2003. Direct estimates of the contribution of viral lysis and microzooplankton grazing to the decline of a Micromonas spp. population, Aquat. Microb. Ecol., 30, 207-219.
- Fischer U.R., Velimirov B., 2002. High control of bacterial production by viruses in a eutrophic oxbow lake, Aquat. Microb. Ecol., 27, 1-12.
- Fuhrman J.A., 1999. Marine viruses and their biogeochemical and ecological effects, Nature, 399, 541‑548.
- Fuhrman J.A., Noble R.T., 1995. Viruses and protists cause similar bacterial mortality in coastal seawater, Limnol. Oceanogr., 40, 1236-1242.
- Guixa-Boixereu N., Vaqué D., Gasol J.M., Sanchez-Camara J.M., Pedros-Alio C., 2002. Viral distribution and activity in Antarctic waters, Deep Sea Res. Pt. II, 49, 827-845.
- Hennes K.P., Simon M., 1995. Significance of bacteriophages for controlling bacterioplankton growth in a mesotrophic lake, Appl. Environ. Microb., 61, 333-340.
- Hofer J.S., Sommaruga R., 2001. Seasonal dynamics of viruses in an alpine lake: importance of filamentous forms, Aquat. Microb. Ecol., 26, 1-11.
- Hornak K., Masin M., Jezbera J., Bettarel Y., Nedoma J., Sime-Ngando T., Simeck K., 2005. Effects of decreased resource availability, protozaon grazing and viral impact on a structure of bacterioplankton assemblage in a canyon-shaped reservoir. FEMS Microb. Ecol., 52, 315-327.
- Jacquet S., Domaison I., Personnic S., Duhamel S., Pradeep Ram A.S., Heldal M., Sime-Ngando T., 2005. Estimates of viral-induced vs. protozoan-induced bacterial mortality in Lake Bourget, France, Freshw. Biol., 50, 627-645.
- Jacquet S., Heldal M., Iglesias-Rodriguez D., Larsen A., Wilson W., Bratbak G., 2002. Flow cytometric analysis of an Emiliana huxleyi bloom terminated by viral infection, Aquat. Microb. Ecol., 27, 111-124.
- Jiang S.C., Paul J.H., 1994. Seasonal and diel abundance of viruses and occurrence of lysogeny/bacteriocinogeny in the marine environnement, Mar. Ecol. Progr. Ser., 104, 163-172.
- Jürgens K., Simek K., 2000. Functional response and particle size selection of Halteria cf. grandinella, a common freshwater oligotrichous ciliate, Aquat. Microb. Ecol., 22, 57‑68.
- Kisand V., Zingel P., 2000. Dominance of ciliate grazing on bacteria during spring in a shallow eutrophic lake, Aquat. Microb. Ecol., 22, 135-142.
- Klut M.E., Stockner J.G., 1990. Virus-like particles in an ultraoligotrophic lake on Vancouver Island, British Columbia, Can. J. Fish. Aquat. Sci., 47, 725-732.
- Landry M.R., Monger B.C., Selph K.E., 1993. Time-dependency of microzooplankton grazing and phytoplankton growth in the subarctic Pacific, Prog. Oceanogr., 32, 205-222.
- Landry M.R., Hasset R.P.,1982. Estimating the grazing impact of marine microzooplankton, Mar. Biol., 67, 283‑288.
- Landry M.R., Kirshtein J., Constantinou J., 1995. A refined dilution technique for measuring the community grazing impact of microzooplankton, with experimental tests in the central equatorial Pacific, Mar. Ecol. Progr. Ser., 120, 53‑63.
- Lebaron P., Servais P., Agogue H., Courties C., Joux F., 2001. Does the high nucleic acid content of individual bacterial cells allow us to discriminate between active cells and inactive cells in aquatic systems?, Appl. Environ. Microb., 67, 1775-1782.
- Li W.K.W., Jellet J. F., Dickie P. M., 1995. DNA distributions in marine bacteria stained with TOTO or TO-PRO, Limnol. Oceanogr., 40, 1485-1495.
- Li W.K.W., 1998. Annual average abundance of heterotrophic bacteria and Synechococcus in surface ocean waters, Limnol. Oceanogr., 43, 1746-1753.
- Maranger R., Bird D.F., 1995. Viral abundance in aquatic systems: a comparison between marine and fresh waters, Mar. Ecol. Progr. Ser., 121, 217-226.
- Maranger R., Bird D.F., Juniper S.K., 1994. Viral and bacterial dynamics in Arctic sea ice during the spring algal bloom, near Resolute, NWT, Canada, Mar. Ecol. Progr. Ser., 111, 121-127.
- Marie D., Vaulot D., Partensky F., 1996. Application of the novel nucleic-acid strains YOYO-1, YO-PRO-1 and PicoGreen for flow cytometic analyses of marine prokaryotes, Appl. Environ. Microb., 62, 1649-1655.
- Marie D., Brussaard C.P.D., Thyrhaug R., Bratbak G., Vaulot D., 1999. Enumeration of marine viruses in culture and natural samples by flow cytometry, Appl. Environ. Microb., 65, 45-52.
- Paul J.H., Rose J.B., Jiang S.C., Kellogg C.A., Dickson L., 1993. Distribution of viral abundance in the reef environment of Key Largo, Florida, Appl. Environ. Microb., 59, 718-724.
- Paul J.H., Jiang S.C., Rose J.B., 1991. Concentration of viruses and dissolved DNA from aquatic environments by vortex flow filtration. Appl. Environ. Microb., 57, 2197‑2204.
- Pédros-Alio C., Caldéron-Paz J. I., Gasol J. M., 2000. Comparative analysis show that bacterivory, not viral lysis, controls the abundance of heterotrophic prokaryotic plankton, FEMS Microb. Ecol., 32, 157-165.
- Proctor L.M., Fuhrman J.A., 1990. Viral mortality of marine bacteria and cyanobacteria, Nature, 343, 60-62.
- Robertson B.R., Button D.K., 1989. Characterizing aquatic bacteria according to population cell size, and apparent DNA content by flow cytometry, Cytometry, 10, 70-76.
- Servais P., Casamayor E.O., Courties C., Catala P., Parthuisot N., Lebaron P., 2003. Activity and diversity of bacterial cells with high and low nucleic acid content. Aquat. Microb. Ecol., 33, 41-51.
- Sime-Ngando T., Bettarel Y., Chartogne C., Sean K., 2003. The imprint of wild viruses on freshwater microbial ecology, Recent Res. Dev. Microbiol, 7, 481-497.
- Sime-Ngando T., Bourdier G., Amblard C., Pinel-Alloul B., 1991. Short-term variations in specific biovolumes of different bacterial forms in aquatic ecosystems, Microb. Ecol., 21, 211-226.
- Simek K., Jürgens K., Nedoma J., Comerma M., Armengol J., 2000. Ecological role and bacterial grazing of Halteria sp: small freshwater oligotrichs as dominant pelagic ciliate bactérivores, Aquat. Microb. Ecol., 22, 43-56.
- Simek K., Pernthaler J., Weinbauer M.G., Hornák K., Dolan J.R., Nedoma J., Masín M., and Amann R., 2001. Changes in bacterial community composition and dynamics and viral mortality rates associated with enhanced flagellate grazing in a mesoeutrophic reservoir, Appl. Environ. Microb., 67, 2723-2733.
- Suttle C.A., 1994. The significance of viruses to mortality in aquatic microbial communities, Microb. Ecol., 28, 237‑243.
- Suttle C.A., Chan A.M., Cottrell M.T., 1990. Infection of phytoplankton by viruses and reduction of primary productivity, Nature, 347, 467-469.
- Suttle C.A., Chen F., 1992. Mechanisms and rates of decay of marine viruses in seawater, Appl. Environ. Microb., 58, 3721‑3729.
- Tapper M.A., Hicks R.E., 1998. Temperate viruses and lysogeny in Lake Superior bacterioplankton, Limnol. & Oceanogr., 43, 95-103.
- Torella F., Morita R., 1979. Evidence by electron micrographs for an incidence of bacteriophage particles in the waters of Yaquina Bay, Appl. Environ. Microb., 37, 774-778.
- Vrede K., Stensdotter U., Lindström E.S., 2003. Viral and bacterioplankton dynamics in two lakes with different humic contents, Microb. Ecol., 46, 406-415.
- Weinbauer M.G., 2004. Ecology of prokaryotic viruses, FEMS Microbiol. Rev., 28, 127-260.
- Weinbauer M.G., Höfle M.G., 1998. Significance of viral lysis and flagellate grazing as factors controlling bacterioplankton production in a eutrophic lake, Appl. Environ. Microb., 64, 431-438.
- Weinbaueur M.G., Rassoulzadegan F., 2004. Are viruses driving microbial diversification and diversity?, Environ. Microbiol., 6, 1-11.
- Weisse T., Scheffel-Möser U., 1990. Growth and grazing loss rates in single-celled Phaeocystis sp. (Prymnesiophyceae), Mar. Biol., 106, 153-158.
- Weisse T., 1993. Dynamics of autotrophic picoplankton in marine and freshwater ecosystems, Adv. Microb. Ecol., 13, 327-370.
- White P.A., Kalff J., Rasmussen J.B., Gasol J.M., 1991. The effect of temperature and algal biomass on bacterial production and specific growth rate in freshwater and marine habitats, Microb. Ecol., 21, 99-118.
- Wilhelm S.W., Brigden S.M., Suttle C.A., 2002. A dilution technique for the direct measurement of viral production: a comparison in stratified and tidally mixed coastal waters, Microb. Ecol., 43, 168-173.
- Wommack K.E., Colwell R.R., 2000. Virioplankton: viruses in aquatic ecosystems, Microbiol. Mol. Biol. Rev., 64, 69‑114.
Liste des figures
Figure 1
Typical flow cytometric dot plots obtained during the study of the in situ microbial population dynamics (A, B) and during the experiments examining the impact of viruses and flagellated protozoa on bacteria (C, D). The flow cytometric signatures of the designated Virus-Like Populations (VLP) referred to as VLP1, VLP2, VLP3 and VLP4, of the picocyanobacteria, the small eukaryotes and the heterotrophic bacteria are indicated. The VLP1 group likely corresponds to bacteriophages. The VLP2 and VLP3 groups might be specific to either cyanobacteria of bacteria (Jacquet, personal observations). The VLP4 is likely to be specific to an eukaryotic alga.
Résultats typiques obtenus par cytométrie en flux pendant l’étude de la dynamique des populations microbiennes in situ (A, B), ainsi que pour les expériences où nous avons examiné l’impact des virus et des protozoaires flagellés sur les bactéries (C, D). Les signatures en cytométrie des populations ressemblant aux virus (VLP1, VLP2, VLP3 et VLP 4) et des populations de picocyanobactéries, de petits eucaryotes et de bactéries hétérotrophes sont indiquées. Le groupement VLP1 correspond probablement aux bactériophages. Les groupements VLP2 et VLP3 seraient peut-être spécifiques aux cyanobactéries et/ou aux bactéries (Jacquet, communication personnelle). Le groupement VLP4 est probablement spécifique à une algue eucaryotique.
Figure 2
In situ population dynamics between February and June 2004. Flow cytometric data are shown in panels A to D. Epifluorescence microscopic data are shown in panels E to H.
Dynamique in situ des populations entre février et juin 2004. Les données issues de la cytométrie en flux figurent dans les panneaux A – D. Les données obtenues en épifluorescence sont présentées dans les panneaux E – H.
Figure 3
Dilution plots and analysis when < 100 KDa and < 0.2 µm filtered lake water were used to dilute the microbial samples (i.e. < 11 µm) during experiment 1 (A) and 2 (B).
Graphiques de dilution et analyse lorsque de l’eau de lac filtrée (< 100 kDa ou < 0,2 µm) est utilisée pour diluer les échantillons microbiens (i.e., < 11 µm) pendant l’expérience 1 (A) et 2 (B).
Figure 4
VLP4 population dynamics during the first experiment (A) and VLP4 concentration plotted against the pigmented eukaryotic community concentration for a dilution series set up with 0.2 µm and 100 KDa diluents (B).
Dynamique de la population VLP4 pendant la première expérience (A), et relation entre la concentration du groupement VPL4 et celle de la communauté eucaryotique pigmentée pour une série de dilutions obtenue avec l’eau filtrée (< 0.2 µm ou < 100 kDa).