Résumés
Abstract
Wheat (Triticumaestivum) relatives are sources of useful genes for disease resistance. Chromosomally segregating populations of intergeneric hybrids between wheat and its distantly related species provide opportunity to study and introgress multiple disease resistance. While introgressing resistance to barley yellow dwarf virus (BYDV) from Thinopyrum into wheat, which is susceptible to BYDV, we scored powdery mildew (Erysiphaegraminis) and leaf rust (Pucciniatriticina) resistance, and chromosome numbers in second and third backcrosses (BC2 and BC3) of intergeneric hybrids of wheat with Thinopyrum ponticum and Thinopyrumintermedium. The frequency of multiple resistance to all the three diseases was low or became low when selection was applied for BYDV resistance and low chromosome numbers. Selection for fewer alien chromosomes while maintaining BYDV resistance was more effective in wheat x T. intermedium than in the wheat x T. ponticum cross. Mean chromosome numbers were significantly different in BC3 generation between BYDV resistant and susceptible plants in both crosses. Significant negative correlations between chromosome numbers and enzyme-linked immunosorbent assay (ELISA) values showed that as the critical Thinopyrum chromosome(s) were eliminated, susceptibility to BYDV increased. Results indicated that it is unlikely that genes for full resistance to all three diseases can be transferred simultaneously from Thinopyrum to wheat.
Résumé
Les espèces proches du blé (Triticumaestivum) sont à l’origine de gènes utiles pour la résistance aux maladies. Les populations à ségrégation chromosomique d’hybrides intergénériques entre le blé et des espèces peu apparentées nous offrent la chance de procéder à l’étude et à l’introgression de la multirésistance aux maladies. Alors que nous introgressions la résistance au virus de la jaunisse nanisante de l’orge (BYDV) de Thinopyrum dans le blé, qui est sensible au BYDV, nous avons coté la résistance à l’oïdium (Erysiphaegraminis) et à la rouille des feuilles (Pucciniatriticina), et compté le nombre de chromosomes dans les deuxième et troisième rétrocroisements (BC2 et BC3) d’hybrides intergénériques entre le blé et le Thinopyrum ponticum ou le Thinopyrumintermedium. Le taux de multirésistance aux trois maladies était bas ou est devenu bas lorsqu’une sélection a été faite pour la résistance au BYDV et pour un faible nombre de chromosomes. La sélection pour un plus faible nombre de chromosomes étrangers tout en maintenant la résistance au BYDV a été plus efficace pour le croisement de blé x T. intermedium que pour celui de blé x T. ponticum. À la génération du BC3, pour les deux croisements, le nombre moyen de chromosomes était significativement différent entre les plantes résistantes et celles sensibles au BYDV. Des corrélations négatives significatives entre le nombre de chromosomes et les valeurs du test immunoenzymatique ELISA ont démontré qu’à mesure que le ou les chromosomes déterminants du Thinopyrum étaient éliminés, la sensibilité au BYDV augmentait. Les résultats montrent qu’il est peu probable que les gènes conférant une résistance complète aux trois maladies puissent être transférés simultanément du Thinopyrum au blé.
Corps de l’article
Introduction
Wheat (Triticumaestivum L.) relatives are useful for wheat improvement for disease resistance and multiple resistance is not uncommon in them (Damania 1993; Fedak 1999; Friebe et al. 1996; Hegde et al. 2002; Sharma et al. 1984; Singh 1993; Wienhues 1963). Transfer of gene(s) for resistance from distantly related (alien) species to wheat through wide crosses may be associated with other desirable genes located in the alien donor species chromosomes. Transfer of multiple resistance in intergeneric crosses would be desirable. Backcrosses of intergeneric hybrids to wheat, in which alien chromosomes are eliminated at random and have wide variation for disease reaction and alien chromosome segregation, provide opportunity to investigate whether resistances to multiple diseases, present in the donor parent, are associated.
Barley yellow dwarf virus (BYDV), transmitted by Rhopalosiphum species of aphid [Homoptera: Aphididae], is one of the most serious viral diseases of wheat and no resistance genes have been found in Triticum species (Comeau and Plourde 1987; Sharma et al. 1997a). Thinopyrumponticum (Podp.) Barkworth & Dewey (syn. Agropyronelongatum (Host) Beauv.) and T. intermedium (Host) Barkworth & Dewey (syn. A. intermedium (Host) Beauv.) are resistant to BYDV (Brettell et al. 1988; Comeau and Plourde 1987; Sharma et al. 1995), leaf rust (Puccinia triticina Erick.) (Cauderon et al. 1973; Franke et al. 1992; Knott 1978; Kochumadhavan et al. 1988; Wienhues 1963), and powdery mildew (Erysiphe (=Blumeria) graminis (DC.) E.O. Speer) (Franke et al. 1992; Sinigovets 1976). These diseases are important in soft red winter wheat production in Indiana (Patterson et al. 1990).
We produced backcross populations from the intergeneric hybrids of wheat (somatic chromosome number = 2n = 42) with T. ponticum (2n = 70) and T. intermedium (2n = 42) primarily to transfer resistance to BYDV from Thinopyrum into wheat (Sharma et al. 1989, 1995). During this introgression for resistance to BYDV, we also compiled data on the reaction of backcross populations to powdery mildew and leaf rust along with somatic chromosome numbers of individual plants. This paper reports the results of analysis of these four traits in the second and third backcross (BC2, BC3) populations.
Materials and methods
The backcross populations were developed from wheat x T. ponticum F1 hybrids, and wheat x T. intermedium F1 hybrids. We used soft red winter wheats in these hybrids and backcrosses (Sharma et al. 1989, 1995). The reaction of the F1 hybrids to the prevalent and virulent strain PAV of BYDV has been described earlier (Sharma et al. 1989). The backcross seedlings at two-three leaf stage were first inoculated with BYDV-PAV by feeding viruliferous aphids on them. Aphid survival was monitored and any plant without live aphids was reinfected (Sharma et al. 1989, 1995). The plants were inoculated with Pucciniatriticina culture No. 901 collected in Indiana. For leaf rust, the seedlings were inoculated in a moisture chamber by applying spore suspension with an automizer and keeping the seedlings in the moisture chamber overnight (Shaner et al. 1997). Subsequently, natural infection by Erysiphegraminis was allowed to occur in a greenhouse where no fungicide was applied. The plants were grown under 20-25oC, 14 h light and natural humidity. Scoring was done 12-14 d after inoculation. We analysed 190 BC2 plants and 209 BC3 plants from the wheat x T. ponticum cross, and were able to score all four traits (BYDV, powdery mildew, leaf rust and chromosome number) on 28 BC2 plants and 49 BC3 plants, and all the three diseases on 63 BC2 plants and 146 BC3 plants. There were 92 BC2 plants and 60 BC3 plants from the wheat x T. intermedium cross, of which 29 and 26, respectively, had all the four traits scored, and 54 and 28, respectively, had all the three diseases scored. Checks for the ELISA were oat cultivar Clintland 64, wheat cultivars Abe, Caldwell and Cardinal, T. ponticum and T. intermedium. Additionally, P-PAV and healthy (uninfected) Clintland 64 were included to rule out variation due to the method. In the powdery mildew and leaf rust tests, the checks were wheat cultivar Morocco, T. ponticum and T. intermedium. The BC2 populations were random while the BC3 were the result of selection in BC2 for fewer alien chromosomes and lower enzyme-linked immunosorbent assay (ELISA) values (measure of level of BYDV resistance). Wheat was the recurrent parent. The Thinopyrum chromosomes do not pair meiotically with wheat chromosomes due to lack of homology and are thus randomly eliminated during backcrossing (Dvorak 1979; Johnson and Kimber 1967; Sharma and Gill 1983). The resistance to the diseases studied thus reflects the effect of the Thinopyrum chromosomes.
The procedure for testing BYDV resistance by ELISA is given in Sharma et al. (1989). No scoring was done on the basis of visual symptoms. Unlike oat, the concentration of virus in infected plants was not consistently correlated with symptom severity in wheat (Baltenberger et al. 1987, Sharma et al. 1989). The chromosome numbers were determined from root-tip cells of the germinated seeds by the Feulgen method (Metz et al. 1988). Plants for powdery mildew and leaf rust were scored as 1 for resistant, 2 for moderately resistant, 3 for moderately susceptible and 4 for susceptible (Hu et al. 1997; Shaner et al. 1997). Actual ELISA values were used for quantitative data analysis. For qualitative analysis, plants with ELISA values 0.3 or less were classified as resistant and those with higher than 0.3 were classified as susceptible (Sharma et al. 1995). Likewise, for qualitative analysis of powdery mildew and leaf rust, scores 1 and 2 were classified as resistant, and scores 3 and 4 as susceptible.
Data analyses were performed using SAS statistical software (SAS Inst. 1999).
Results and discussion
Results of our study showed that the possibility of having multiple resistance to all three diseases was low. Of the plants scored for all the four traits (three diseases and chromosome numbers), 53.6% and 6.9% were resistant to all the three diseases (powdery mildew, leaf rust and BYDV) in BC2 in wheat x T. ponticum cross and wheat x T. intermedium cross, respectively (Table 1). The proportion of plants in BC3 resistant to all the three diseases after selection for resistance to BYDV and fewer alien chromosomes in BC2 declined to 4.1% and 3.8%, respectively. The trend remained the same when larger population, scored for all the three diseases including those without chromosome counts, was analysed (Table 1). Taking two diseases at a time, the proportion of BYDV resistant plants with resistance to powdery mildew or leaf rust also decreased in the BC3 generation when compared to the BC2 generation in both crosses (Table 2). Only 9.8% and 6.8% of the BC2 plants resistant to powdery mildew and leaf rust, respectively, were resistant to BYDV in wheat x T. intermedium cross (data not shown). These percentages were higher (72.6% and 69.5%, respectively) in wheat x T. ponticum cross but were reduced drastically in BC3 to 8.3% and 7.7%, respectively, once selection was applied in the BC2 generation in favor of BYDV resistance and lower chromosome numbers (data not shown). The proportion of leaf rust resistant plants among powdery mildew resistant plants also decreased in going from BC2 to BC3 generation in both crosses (from 97.7% to 76.2% in wheat x T. ponticum cross and from 81.4% to 62.5% in wheat x T. intermedium cross) (data not shown). No selection was applied to leaf rust or powdery mildew and the decrease was probably due to random elimination of Thinopyrum chromosomes that carried resistance to these diseases.
Table 1
Percentage of plants resistant to all three diseases (BYDV, powdery mildew and leaf rust) in BC2 and BC3 populations (values in parentheses are for the plants scored for all the three diseases but not all of them scored for chromosome numbers)
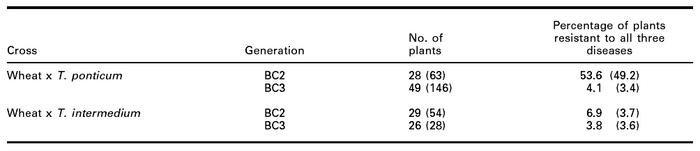
Table 2
Percentage of barley yellow dwarf virus (BYDV) resistant plants (BYDV resistant plant numbers are given in parantheses) that were resistant to powdery mildew (PM) or leaf rust (LR) in BC2 and BC3 generations (data include whole populations with two or all three diseases scored)

In both crosses, mean values increased significantly (increased susceptibility) for powdery mildew, leaf rust and BYDV, and mean chromosome numbers decreased significantly, in the BC3 populations compared to the BC2 populations, except for BYDV and leaf rust in the wheat x T. intermedium cross, indicating increase in susceptibility with reduction in chromosome numbers (Table 3). Percentage of resistant plants decreased in both crosses for powdery mildew, leaf rust and BYDV, except for BYDV in wheat x T. intermedium cross. The resistance in BC2 generation of wheat x T. ponticum cross was relatively high but decreased in BC3. Barring sample size effect, comparison of percent resistant plants showed that selection for fewer chromosomes while maintaining BYDV resistance was more effective in the wheat x T. intermedium cross than in wheat x T. ponticum cross (Table 3). It appears that the genetic control for resistance to the three diseases studied is not on one chromosome.
Table 3
Ranges, means and standard deviations (SD) for powdery mildew (PM) scores, leaf rust (LR) scores, BYDV (ELISA) values and chromosome number (No. of plants and percent resistant plants are given in parantheses in each case; e = wheat x T. ponticum cross; i = wheat x T. intermedium cross)
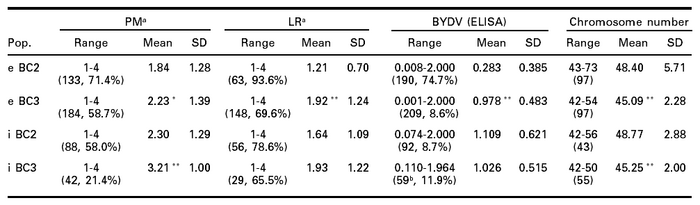
a For PM and LR, 1 = resistant, 2 = moderately resistant, 3 = moderately susceptible and 4 = susceptible, (1,2) = resistant, (3,4) = susceptible.
b Of the total 60 plants, one was not analyzed by ELISA.
*,** BC3 mean signifiantly different from BC2 mean at P = 0.05 and P = 0.01, respectively, for each of the four traits (PM, LR, ELISA and chromosome number) within each of the two crosses (e, i.).
T-test showed that the mean chromosome numbers were significantly different between BYDV resistant and susceptible plants in BC3 in both crosses, and in BC2 of wheat x T. ponticum cross (Table 4). Mean chromosome numbers between leaf rust resistant and susceptible plants in BC3 of wheat x T. ponticum cross, and between powdery mildew resistant and susceptible plants in BC2 of wheat x T. intermedium cross were also significantly different. In other cases, mean chromosome numbers were not statistically different. In all cases of significant differences, the chromosome numbers were significantly lower in susceptible plants, except for leaf rust in BC3 of wheat x T. ponticum cross (Table 4). T-test indicated that mean ELISA values were significantly different in both generations in wheat x T. ponticum cross between powdery mildew resistant and susceptible plants as well as between leaf rust resistant and susceptible plants (Table 5). The mean ELISA values were significantly higher in BC2 but significantly lower in BC3 for the plants susceptible to leaf rust or powdery mildew. The differences were not significant in the wheat x T. intermedium cross.
Table 4
T-test for chromosome numbers on BC2 and BC3 between resistant and susceptible plants for powdery mildew (PM), leaf rust (LR), or for BYDV

*, ** Significant at P = 0.05 and P = 0.01, respectively.
a All plants resistant (no susceptible plants with chromosome numbers).
b Significant at P = 0.10.
Table 5
T-test for ELISA on BC2 and BC3 between resistant and susceptible plants for powdery mildew (PM), or leaf rust (LR)

*, ** Significant at P = 0.05 and P = 0.01, respectively.
Correlation coefficients between chromosome numbers and ELISA values were negative for all the four populations (data not shown). They were highly significant (P < 0.01) for both BC3 polulations (r = -0.296 for wheat x T. ponticum cross and r = -0.371 for wheat x T. intermedium cross), and significant for BC2 of wheat x T. ponticum cross (r = -0.172, P < 0.10) and wheat x T. intermedium cross (r = -0.330, P < 0.05). These results showed that as the critical or major Thinopyrum chromosome(s) were eliminated, susceptibility increased.
Chi-square test by 2x2 contingency analysis of the three diseases showed that the patterns of resistance were significantly different for powdery mildew and leaf rust in BC2 and BC3, and for BYDV and leaf rust in BC3 in wheat x T. ponticum cross (Table 6). At 10% level of confidence, chi-square for BYDV and powdery mildew was significant in BC3 in the wheat x T. intermedium cross. Other chi-squares were non-significant i.e. the patterns of resistance or susceptibility were similar for any two diseases compared.
Table 6
Frequency distribution by 2x2 contingency table for BC2 and BC3 plants for the three diseases, PM, LR and BYDV (resistant = R, susceptible = S)
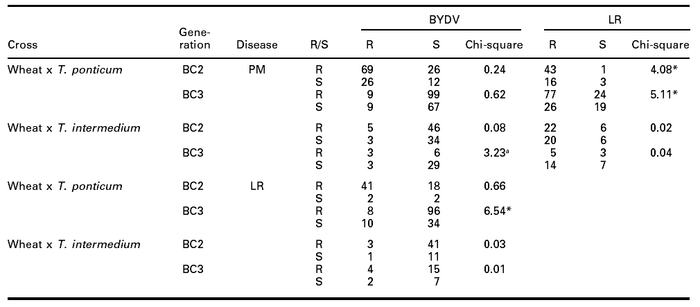
* Significant at P = 0.05.
a Significant at P = 0.10.
From progeny of a cross between wheat x T. junceiforme Love & Love (Love), Ellneskog (2002) obtained lines resistant to powdery mildew and leaf rust. Li et al. (1998) obtained wheat x A. cristatum (L.) Gaertner derivatives which were more resistant to both powdery mildew and BYDV compared to wheat. Such an association was not evident in the present intergeneric crosses. Larkin et al. (1995) reported that out of the disomic alien addition lines isolated from a partial amphiploid (2n = 56) of wheat x T. intermedium, one was resistant to BYDV, and one to leaf, stem and stripe rusts. In the study of Comeau et al. (1994), Agrotricum (Agropyron x Triticum) line OK211542 (2n = 56) was immune to BYDV. Through a selection for BYDV and threshable phenotype with wheat-like seed, F7-derived lines were obtained which were immune to BYDV with 44 somatic chromosomes. The low frequency of expression of immunity suggested that more than one gene was required for full expression of resistance (Comeau et al. 1994). Anderson et al. (1998) have shown that the full BYDV resistance of T. intermedium was not realized in the wheat-T. intermedium chromosome substitution line P29 for group 7 chromosome developed by Sharma et al. (1997b).
Sharma et al. (1997b) reported that substitution line P29 was resistant to BYDV but susceptible to four cultures of powdery mildew. Genes for resistance to Hessian fly (Mayetioladestructor Say) [Diptera: Ceci-domyiidae] biotype L and BYDV were also found to be on different chromosomes of T. intermedium (Sharma et al. 1992, 1997b). This is consistent with the present finding that these resistances are not carried in a linkage block in T. intermedium. P29 showed resistance to leaf rust. It is not known whether this resistance was due to gene(s) in wheat or T. intermedium, or to BYDV reducing the resistance to leaf rust in the present study. BYDV infection increased the susceptibility of wheat to Gibberellazeae (Schwein.) Petch, Alternaria and Cladosporium (Smith 1962), Cochliobolussativus (Ito & Kuribayashi) Drechs. ex Dastur (Scott 1968), and Gaeumannomycesgraminis (Sacc.) Arx & D. Olivier (Price 1970). The present work did not include experiments to study synergetic effects. The results of our study indicated that whereas some gene(s) for resistance to leaf rust may be introgressed along with genes for resistance to BYDV, major genes for resistance to BYDV, powdery mildew and leaf rust cannot be bred simultaneously from Thinopyrum into wheat. Since we know little about the genetic control of resistance to these diseases, it is difficult to speculate the physical distribution and possible linkage.
Parties annexes
Acknowledgement
Purdue Agric. Res. Programs Journal No. 17245. Judy Santini’s professional help in the statistical analysis is gratefully praised.
References
- Anderson, J., D. Bucholtz, A. Greene, M. Francki, S. Gray, H. Sharma, H. Ohm, and K. Perry. 1998. Characterization of wheatgrass-derived barley yellow dwarf virus resistance in wheat alien chromosome substitution line. Phytopathology 88 : 851-855.
- Baltenberger, D., H. Ohm, and J. Foster. 1987. Reactions of oats, barley and wheat to infection with barley yellow dwarf virus isolates. Crop Sci. 27 : 195-198.
- Brettell, R., P. Banks, Y. Cauderon, X. Chen, Z. Cheng, P. Larkin, and P. Waterhouse. 1988. A single wheatgrass chromosome reduces the concentration of barley yellow dwarf virus in wheat. Ann. Appl. Biol. 113 : 599-603.
- Cauderon, Y., B. Saigne, and M. Dauge. 1973. The resistance to wheat rusts of Agropyronintermedium and its use in wheat improvement. Int. Wheat Genet. Symp. 4 : 401-407.
- Comeau, A., and A. Plourde. 1987. Cell, tissue culture and intergeneric hybridization for barley yellow dwarf virus resistance in wheat. Can. J. Plant Pathol. 9 : 188-192.
- Comeau, A., K. Makkouk, F. Ahmad, and C. St-Pierre. 1994. Bread wheat x agrotricum crosses as a source of immunity and resistance to the PAV strain of BYDV luteovirus. Agronomie 14 : 153-160.
- Damania, A. 1993. Biodiversity and wheat improvement. John Wiley & Sons, West Sussex, UK. 434 pp.
- Dvorak, J. 1979. Metaphase I pairing frequencies of indivi-dual Agropyronelongatum chromosome arms with Triticum chromosomes. Can. J. Genet. Cytol. 21 : 243-254.
- Ellneskog, S. 2002. Relationship in the Triticeae genomes and wide hybridizations. Acta Univ. Agric. Sueciae Agraria 357 : 1-85.
- Fedak, G. 1999. Molecular aids for integration of alien chromatin through wide crosses. Genome 42 : 584-591.
- Franke, R., R. Nestrowicz, A. Senula, and B. Staat. 1992. Intergeneric hybrids between Triticumaestivum L. and wild Triticeae. Hereditas 116 : 225-231.
- Friebe, B., J. Jiang, W. Raupp, R. McIntosh, and B. Gill. 1996. Characterization of wheat-alien translocations with resistance to diseases and pests: current status. Euphytica 91 : 59-87.
- Hegde, S., J. Valkoun, and J. Waines. 2002. Genetic diversity in wild and weedy Aegilops, Amblyopyrum and Secale species – a preliminary survey. Crop Sci. 42 : 608-614.
- Hu, X., H. Ohm, and I. Dweikat. 1997. Identification of RAPD markers linked to the gene PM1 for resistance to powdery mildew in wheat. Theor. Appl. Genet. 94 : 832-840.
- Johnson, R., and G. Kimber. 1967. Homoeologous pairing of chromosomes from Agropyronelongatum with those of Triticumaestivum and Aegilopsspeltoides. Genet. Res. 10 : 63-71.
- Knott, D. 1978. The transfer of genes for rust resistance to wheat from related species. Int. Wheat Genet. Symp. 5 : 354-357.
- Kochumadhavan, M., S. Tomar, and P. Nambisan. 1988.Agropyron-derived specific genes in common wheat and their adult plant response to wheat pathogenes. Indian J. Genet. Plant Breed. 48 : 383-387.
- Larkin, P., P. Banks, E. Lagudah, R. Appels, X. Chen, Z. Xin, H. Ohm, and R. McIntosh. 1995. Disomic Thinopyrumintermedium addition lines in wheat with barley yellow dwarf virus (BYDV) resistance and with rust resistances. Genome 38 : 385-394.
- Li, L., X. Yang, X. Li, Y. Dong, and X. Chen. 1998. Intro-duction of desirable genes from Agropyroncristatum to wheat by intergeneric hybridization. Sci. Agric. Sin. 31 : 1-5.
- Metz, S., H. Sharma, T. Armstrong, and P. Mascia. 1988. Chromosome doubling and aneuploidy in anther-derived plants from two winter wheat lines. Genome 30 : 177-181.
- Patterson, F., G. Shaner, J. Foster, and H. Ohm. 1990. A historical perspective for the establishment of research goals for wheat improvement. J. Prod. Agric. 3 : 30-38.
- Price, R. 1970. Stunted patches and dead heads in Victorian cereal crops. Page 165 in Technical Pub. No. 23. Dept. Agric. Victoria, Australia.
- SAS Institute. 1999. SAS/STAT user’s guide. Version 8. SAS Inst., Cary, NC.
- Scott, D. 1968. Effect of barley yellow dwarf virus infection on the development of root rot caused by Colchiobolussativa in Avenasativa and Triticum durum. Ph.D. Thesis, University of Illinois, USA. 106 pp.
- Shaner, G., G. Buechley, and W. Niquist. 1997. Inheritance of latent period of Pucciniarecondita in wheat. Crop Sci. 37 : 748-756.
- Sharma, H., and B. Gill. 1983. New hybrids between wheat and Agropyron. 2. Production, morphology and cytogenetic analysis of F1 hybrids and backcross derivatives. Theor. Appl. Genet. 66 : 111-121.
- Sharma, H., B. Gill, and J. Uyemoto. 1984. High levels of resistance in Agropyron species to barley yellow dwarf and wheat streak mosaic viruses. Phytopathol. Z. 110 : 143-147.
- Sharma, H., H. Ohm, R. Lister, J. Foster, and R. Shukle. 1989. Response of wheatgrasses and wheat x wheatgrass hybrids to barley yellow dwarf virus. Theor. Appl. Genet. 77 : 369-374.
- Sharma, H., J. Foster, H. Ohm, and F. Patterson. 1992. A note on resistance to Hessian fly (Mayetioladestructor Say) [Diptera: Cecidomyiidae] biotype L in tribe Triticeae. Phytoprotection 73 : 79-82.
- Sharma, H., H. Ohm, L. Goulart, R. Lister, R. Appels, and O. Benlhabib. 1995. Introgression and characterization of barley yellow dwarf virus resistance from Thinopyrumintermedium into wheat. Genome 38 : 406-413.
- Sharma, H., H. Ohm, and R. Lister. 1997a. Reaction of near relatives of wheat to P-PAV isolate of BYDV. Barley Yellow Dwarf Newsl. 6 : 7-8.
- Sharma, H., H. Ohm, and K. Perry. 1997b. Registration of barley yellow dwarf virus resistant wheat germplasm line P29. Crop Sci. 37 : 1031-1032.
- Singh, R. 1993. Genetic association of gene Bdv1 for tolerance to barley yellow dwarf virus with genes Lr34 and Yr18 for adult plant resistance to rust in bread wheat. Plant Dis. 77 : 1103-1106.
- Sinigovets, M. 1976. Effect of single Agropyron chromosome on wheat. Genetika 12 : 15-21.
- Smith, H. 1962. Is barley yellow dwarf virus a predisposing factor in the common root rot disease of wheat in Cana-da? Can. Plant Dis. Survey 42 : 143-148.
- Wienhues, A. 1963. Transfer of rust resistance of Agropyron to wheat by addition, substitution and translocation. Int. Wheat Genet. Symp. 2 : 328-341.
 10.7202/706023ar
10.7202/706023arListe des tableaux
Table 1
Percentage of plants resistant to all three diseases (BYDV, powdery mildew and leaf rust) in BC2 and BC3 populations (values in parentheses are for the plants scored for all the three diseases but not all of them scored for chromosome numbers)
Table 2
Percentage of barley yellow dwarf virus (BYDV) resistant plants (BYDV resistant plant numbers are given in parantheses) that were resistant to powdery mildew (PM) or leaf rust (LR) in BC2 and BC3 generations (data include whole populations with two or all three diseases scored)
Table 3
Ranges, means and standard deviations (SD) for powdery mildew (PM) scores, leaf rust (LR) scores, BYDV (ELISA) values and chromosome number (No. of plants and percent resistant plants are given in parantheses in each case; e = wheat x T. ponticum cross; i = wheat x T. intermedium cross)
a For PM and LR, 1 = resistant, 2 = moderately resistant, 3 = moderately susceptible and 4 = susceptible, (1,2) = resistant, (3,4) = susceptible.
b Of the total 60 plants, one was not analyzed by ELISA.
*,** BC3 mean signifiantly different from BC2 mean at P = 0.05 and P = 0.01, respectively, for each of the four traits (PM, LR, ELISA and chromosome number) within each of the two crosses (e, i.).
Table 4
T-test for chromosome numbers on BC2 and BC3 between resistant and susceptible plants for powdery mildew (PM), leaf rust (LR), or for BYDV
*, ** Significant at P = 0.05 and P = 0.01, respectively.
a All plants resistant (no susceptible plants with chromosome numbers).
b Significant at P = 0.10.
Table 5
T-test for ELISA on BC2 and BC3 between resistant and susceptible plants for powdery mildew (PM), or leaf rust (LR)
*, ** Significant at P = 0.05 and P = 0.01, respectively.
Table 6
Frequency distribution by 2x2 contingency table for BC2 and BC3 plants for the three diseases, PM, LR and BYDV (resistant = R, susceptible = S)
* Significant at P = 0.05.
a Significant at P = 0.10.