Résumés
Résumé
Les endothélines, ET-1, ET-2 et ET-3 sont trois peptides de 21 acides aminés, qui ont été découverts au travers de leurs puissants effets vasoconstricteurs. Les précurseurs des endothélines sont clivés en peptides actifs par l’enzyme de conversion de l’endothéline. Deux récepteurs, ET-A et ET-B, constitués de sept domaines transmembranaires ont été clonés chez les mammifères. Le développement d’agonistes et d’antagonistes sélectifs des deux types de récepteurs et l’élaboration de modèles animaux génétiquement modifiés ont permis de montrer que le système endothéline n’était pas seulement important dans le domaine cardiovasculaire. Chez les mammifères, ce système est essentiel pour le développement embryonnaire et néonatal, et joue un rôle dans l’homéostasie rénale, le maintien du tonus vasculaire, la réparation du tissu cardiaque, l’angiogenèse et la régulation de la respiration. Mais le système endothéline est aussi impliqué de manière délétère dans l’athérosclérose, le remodelage cardiaque et l’hypertension pulmonaire.
Summary
Endothelins are a family of three peptides of 21 amino acids with strong vasoconstrictor effects. The three peptides are encoded by three different genes and derived from precursors (« big endothelins ») which are cleaved by metalloproteases, named endothelin-converting enzyme. Two receptors have been cloned, ET-A and ET-B which bind the three endothelins with various affinities. The diverse expression pattern of the endothelin system (ET) components is associated with a complex pharmacology and its counteracting physiological actions. New modulators of the ET system have been described : retinoic acid, leptin, prostaglandins, hypoxia. Endothelins can be considered as regulators working in paracrine and autocrine fashion in a variety of organs in different cellular types. The ET system has beneficial and detrimental roles in mammals. The different components have been shown to be essential for a normal embryonic and neonatal development, for renal homeostasis and maintenance of basal vascular tone. They are involved in physiological and tumoral angiogenesis. They affect the physiology and pathophysiology of the liver, muscle, skin, adipose tissue and reproductive tract. The endothelin system participates in the development of atherosclerosis as well as pulmonary hypertension, and mediates cardiac remodeling in heart failure. Elaboration of new animal models (knock-out, pathophysiological models…) will allow the clear genetic dissection of physiological and pathophysiological roles of the endothelin system.
Corps de l’article
Dans les années 1980, le rôle des cellules endothéliales en tant qu’élément actif du système vasculaire a permis de cloner un facteur vasoconstricteur, l’endothéline-1 (ET-1) à partir de cellules endothéliales dérivées d’aorte de porc [1].
Le système endothéline (ET) est constitué de deux récepteurs couplés aux protéines G, ET-A et ET-B, et de deux enzymes de maturation, une protéase de type furine et une enzyme de conversion de l’endothéline.
Une analyse fonctionnelle du système ET a été effectuée grâce aux études pharmacologiques utilisant des antagonistes des récepteurs de l’endothéline spécifiques de chaque type, et au développement de modèles murins génétiquement modifiés.
Biosynthèse des constituants du système endothéline
L’ET-1 est un peptide de 21 acides aminés comportant une extrémité carboxy-terminale hydrophobe et 2 ponts disulfures en position amino-terminale. En 1989, deux peptides ont été identifiés différant de ET-1 par trois et six acides aminés et ont été appelés respectivement, endothéline-2 (ET-2) et endothéline-3 (ET-3) [2]. Les précurseurs des endothélines sont clivés par deux protéases pour produire les endothélines matures et actives. Les préproendothélines (PPET) sont clivées au niveau de sites dibasiques par des endopeptidases de type furine (Figure 1) pour former des intermédiaires biologiquement inactifs, appelés big endothélines (big ET).
Figure 1
Genèse des endothélines.
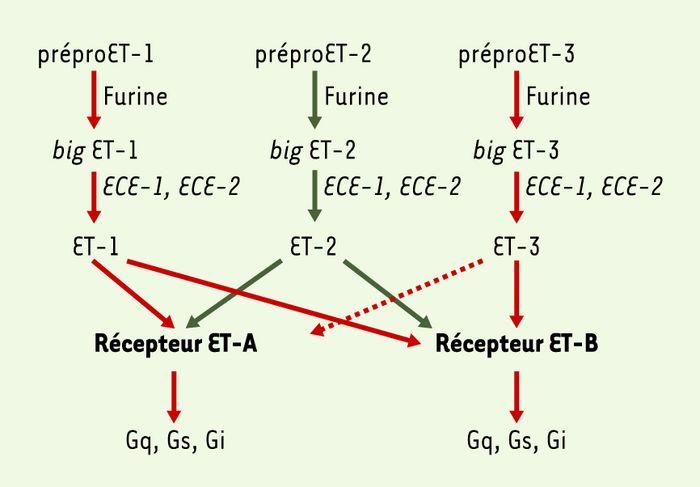
Les précurseurs des endothélines sont clivés par deux groupes de protéases pour produire les endothélines matures et actives. Les préproendothélines (prépro ET-1, -2, -3) constituées d’environ 200 résidus sont clivés au niveau de sites dibasiques par des endopeptidases de type furine pour former des intermédiaires biologiquement inactifs appelés big endothélines (big ET-1, 2, 3) qui sont des peptides de 37 à 41 acides aminés. Les big endothélines sont ensuite clivées au niveau de la liaison Trp-Val de la big ET-1 et de la big ET-2 ou au niveau de la liaison Trp-Ile de la big ET-3 pour former le peptide final de 21 acides aminés (ET-1, 2). Cette dernière étape est effectuée par des métalloprotéases membranaires à zinc de la superfamille des néprilysines [3], appelées enzymes de conversion de l’endothéline (ECE-1, 2). Les peptides endothélines (ET-1, 2, 3) produits interagissent de manière sélective avec le récepteur ET-A et de manière non sélective avec le récepteur ET-B. Ces deux récepteurs sont couplés à des protéines G variées (Gq, Gs, Gi) permettant différentes réponses physiologiques. L’affinité du récepteur ET-A pour les peptides ET-1 et ET-2 est de l’ordre de la nanomole (flèches en trait plein) et celle pour ET-3 est plus faible de deux ordres de grandeur (flèche en trait pointillé). En revanche, le récepteur ET-B a la même affinité pour les trois peptides (de l’ordre de la nanomole).
Les big ET sont ensuite clivées au niveau de la liaison Trp-Val de la big ET-1 et de la big ET-2 ou au niveau de la liaison Trp-Ile de la big ET-3 (Figure 1). Cette dernière étape est effectuée par des métalloprotéases membranaires à zinc de la superfamille des néprilysines [3], appelées enzymes de conversion de l’endothéline (ECE).
Actuellement, deux membres sont connus, l’ECE-1 [4] et l’ECE-2 [5]. L’ECE-1 est exprimée dans un grand nombre de cellules [6], son activité est maximale à pH neutre et elle clive les big ET aussi bien à l’intérieur qu’à la surface des cellules [4, 7]. L’ECE-2 est localisée dans divers types cellulaires et son activité, maximale à pH acide (pH 5,8), suggère son implication comme enzyme de maturation intracellulaire [5]. L’ECE-1 et l’ECE-2 clivent de manière préférentielle la big ET-1 in vivo et de manière équivalente la big ET-2 et la big ET-3 in vitro. Le fait que les souris doublement invalidées pour les gènes de l’ECE-1 et de l’ECE-2 produisent toujours des concentrations élevées de peptides ET matures suggère qu’une autre enzyme non identifiée puisse effectuer cette étape finale de maturation [8].
Pour l’ECE-1 d’origine humaine, quatre isoformes produites par épissage alternatif à partir d’un gène unique ont été mises en évidence (Figure 2) [9]. Ces isoformes diffèrent uniquement par leur séquence amino-terminale (Figure 2) et leur distribution subcellulaire. L’expression stable de ces quatre isoformes dans des cellules CHO a permis de montrer que les isoformes a, c et d sont présentes principalement à la surface des cellules tandis que l’isoforme b est essentiellement intracellulaire [10].
Figure 2
Isoformes de l’enzyme de conversion humaine de l’endothéline (ECE-1).
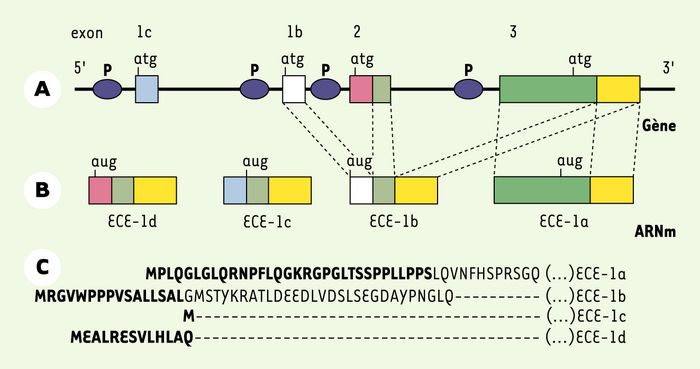
ECE-1 est codée par un seul gène qui subit un épissage alternatif permettant la création de quatre isoformes, appelées a, b, c et d. A. Schéma de la structure partielle du gène ECE-1. Seuls quatre premiers exons du gène sont représentés (1c, 1b, 2, 3). La position du site d’initiation de la traduction est indiquée (atg). Les ellipses bleues représentent les isoformes du promoteur (P) présomptif du gène. B. Production des ARN messagers ECE-1a, 1b, 1c, 1d par épissage alternatif. Pour la clarté du schéma, seuls les traits en pointillé indiquant l’épissage des formes ECE-1a et 1b sont figurés. C. Séquence amino-terminale des quatre isoformes. Pour ECE-1b, 1c et 1d, les traits en pointillé indiquent l’identité des résidus avec la ligne précédente. Les résidus en gras correspondent à la séquence codée par l’exon 3 pour ECE-1a, par l’exon 1b pour ECE-1b, par l’exon 1c pour ECE-1c, et par l’exon 2 pour ECE-1d.
D’autres substrats ont été recherchés pour l’ECE-1. Si ce peptide big ET-1 est le substrat clivé le plus efficacement par ECE-1, la neurotensine, la substance P et la bradykinine le sont également, bien que les constantes catalytiques (kcat ) soient beaucoup plus élevées [11].
Chez les mammifères, deux récepteurs de l’endothéline (ET-A et ET-B) ont été identifiés et clonés [2]. ET-A et ET-B sont des récepteurs à sept domaines transmembranaires hydrophobes de 427 et 416 acides aminés respectivement, dont la séquence n’a que 59% de similitude. Chaque récepteur active les mêmes protéines G, donnant cependant des réponses différentes suivant le type cellulaire, comme l’activation des phospholipases C, l’augmentation de calcium intracellulaire et l’induction de gènes précoces.
Le récepteur ET-A a des affinités de l’ordre de la nanomole pour les peptides ET-1 et ET-2 et une affinité plus faible pour ET-3. En revanche, le récepteur ET-B a la même affinité, de l’ordre de la nanomole, pour les trois peptides (Figure 1).
Expression et modulation du système endothéline
Expression du système endothéline
La physiologie complexe du système ET-1 est en rapport avec un profil diversifié d’expression de ses constituants (Tableau I) [12].
Tableau I
Profil d’expression des principaux constituants du système endothéline.
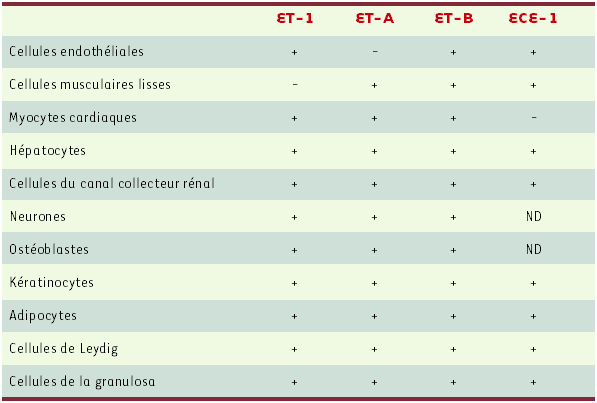
Le signe + indique la présence du composant, le signe - son absence. ND : non déterminé. La distribution d’ET-2, d’ET-3 et d’ECE-2 n’est pas indiquée car ces constituants n’ont pas été étudiés de manière exhaustive.
ET-1 est produit par les cellules endothéliales, les cellules épithéliales, les macrophages, les fibroblastes, les myocytes cardiaques, les neurones. In vivo, les cellules endothéliales sont la source majeure d’ET-1. ET-2 est exprimé par les cellules épithéliales de l’intestin et ET-3 par les neurones, les cellules épithéliales tubulaires du rein et les cellules de l’intestin. Dans plusieurs tissus, les récepteurs ET-A et ET-B sont présents dans les mêmes types cellulaires, excepté les cellules endothéliales où ET-B est majoritairement présent. Dans le foie, ET-A est présent dans les cellules de Kupffer et ET-B dans les cellules de Ito.
ECE-1 est exprimée dans la plupart des tissus, suggérant un rôle majeur de cette enzyme. Une très forte expression a été observée dans la surrénale [6]. La présence d’ET-1 dans les structures vasculaires suggère une action autocrine et/ou paracrine sur le contrôle du flux sanguin dans la surrénale [13]. Les concentrations extrêmement élevées d’ECE-1 font penser que d’autres substrats que la big ET-1 sont clivés.
Cette description montre le rôle majeur d’ET-1 via le clivage par ECE-1 de la big ET-1. La distribution pratiquement ubiquitaire de ses deux récepteurs (cibles d’ET-1) illustre l’importance du site de production d’ET-1 pour son action physiologique.
Modulation du système endothéline
Durant ces dernières années, de nouveaux modulateurs des différents constituants du système ET ont été décrits. Certains interviennent dans la croissance cellulaire, d’autres dans les maladies cardiovasculaires.
Les acides rétinoïques inhibent la croissance cellulaire, et ont donc une action anti-athérogène potentielle dans les vaisseaux. Les acides rétinoïques répriment également l’expression de l’ARNm codant pour la PPET-1 et l’activité transcriptionnelle de son promoteur [14]. Ces résultats suggèrent que les rétinoïdes pourraient être des modulateurs de la fonction endothéliale.
Les prostaglandines, notamment la prostaglandine F2α, contrôlent l’expression de différents composés du système ET (ECE-1, ET-A et ET-1) dans les phases de lutéinisation [15].
Il a été montré in vitro que la leptine induisait une stimulation de la synthèse et de la sécrétion d’ET-1 à partir de cultures primaires de cellules endothéliales [16].
Une inhibition de la production d’ET-1 par les astrocytes soumis à une hypoxie, associée à une augmentation d’activité ECE-1 à été mise en évidence [17]; cette inhibition n’est pas retrouvée dans les cultures d’astrocytes issus d’animaux invalidés pour le gène ET-B.
Dans les cellules endothéliales cultivées en hypoxie, les facteurs AP-1 (activating protein-1), GATA-2 et NF-1 (nuclear factor-1) sont impliqués dans la stabilisation de la liaison du facteur HIF-1 (hypoxia-inducible factor-1) sur le promoteur d’ET-1 en induisant le recrutement de la protéine p300/CBP (CREB binding protein) [18]. Le facteur de transcription, Vezf1/DB1 (vascular endothelial zinc finger 1), se lie au promoteur d’ET-1 pour stimuler sa production dans l’endothélium durant le développement, et maintenir le tonus vasculaire [19].
Fonctions in vivo du système endothéline
Les endothélines agissent de manière paracrine et autocrine dans une multitude d’organes, avec des rôles bénéfiques ou délétères chez les mammifères (Tableau II).
Tableau II
Fonctions in vivo proposées pour les principaux constituants du système endothéline.
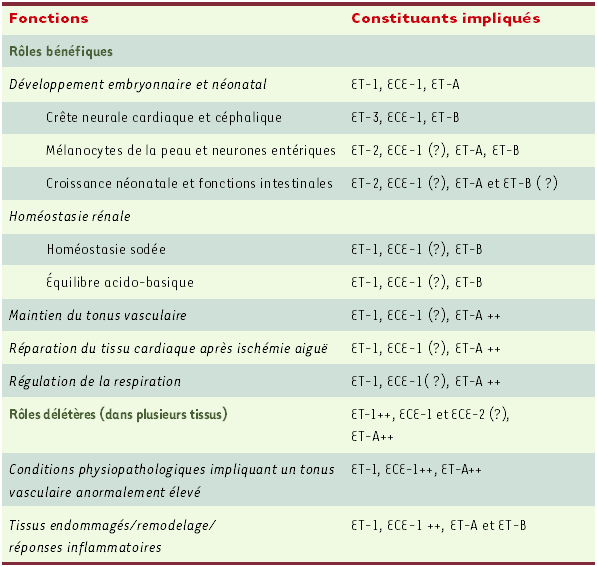
La présence d’un point d’interrogation correspond à un rôle possible; ++ signifie essentiellement.
Rôles bénéfiques du système endothéline
Développement embryonnaire et néonatal
Les expériences d’invalidation de gènes du système ET chez la souris ont montré l’importance de ce système dans le développement. Les gènes ET-1, ET-A et ECE-1 sont nécessaires au développement des structures cranio-faciales et des éléments du système d’éjection cardiaque dérivés des crêtes neurales [20, 21]. Les souris invalidées pour les gènes codant pour ET-1 et ET-A meurent à la naissance par asphyxie, en raison de malformations cranio-faciales et pharyngo-laryngées. D.E. Clouthier et al. [22] ont montré que le récepteur ET-A devait être activé par ET-1 au stade embryonnaire E10 pour assurer une prolifération normale des cellules de la crête neurale. Ces souris présentent également des anomalies artérielles [20-22].
Les souris invalidées pour les gènes ET-B ou ET-3 sont viables à la naissance, mais elles ne se développent pas et meurent après trois à six semaines. Les souris déficitaires en ET-3, ET-B ou ECE-1 n’ont pas de mélanocytes au niveau de l’épiderme, ce qui entraîne un défaut de pigmentation de la peau [21, 23]. De plus, il n’y a pas de développement correct des neurones mésentériques, ce qui entraîne un dysfonctionnement intestinal et la mort des animaux [21, 23]. Récemment, l’expression de l’ECE-1 a été détectée dans les kératinocytes, permettant la maturation et la sécrétion d’ET-1 par les kératinocytes, ce qui stimule la pigmentation de l’épiderme [24].
Les souris invalidées pour le gène de l’ECE-1 ont un phénotype additif correspondant à celui des souris invalidées pour ET-1 ou ET-A et ET-3 ou ET-B. Des anomalies cardiaques plus graves ont été observées chez les souris doublement invalidées pour ECE-1 et ECE-2 en comparaison de celles observées chez les souris ECE-1-/- [8].
En utilisant le modèle de l’embryon de poulet, l’inactivation pharmacologique du récepteur ET-A par des antagonistes spécifiques a confirmé l’implication de ce récepteur dans l’organogenèse cranio-faciale et cardiovasculaire [25]. L’intérêt de ce modèle réside dans sa simplicité et dans la rapidité d’obtention des résultats avec cependant le problème de la disponibilité d’antagonistes ou d’agonistes stables in vivo chez le poulet.
Homéostasie rénale
Dans le rein, le système ET contrôle l’excrétion d’eau et de sodium, ainsi que l’équilibre acido-basique. Il a été montré in vitro que l’acidose métabolique chronique augmentait l’expression d’ET-1 par un accroissement de l’activité de l’antiporteur NHE3 (échangeur Na+/H+) via le récepteur ET-B [26]. Cette induction serait due à une augmentation du trafic intracellulaire de NHE3 au pôle apical des cellules épithéliales [27].
Maintien du tonus vasculaire
Depuis sa découverte, le peptide ET-1 a été uniquement identifié au niveau des vaisseaux dans lesquels il est sécrété par les cellules endothéliales [1]. La concentration locale d’ET-1 dans la paroi vasculaire est 100 fois supérieure à celle qui est observée dans le plasma. L’utilisation d’antagonistes des récepteurs ET a mis clairement en évidence l’implication du système ET dans le tonus vasculaire [2].
Après la survenue d’un infarctus du myocarde, le système ET semble jouer un double rôle : bénéfique concernant la réparation tissulaire et la restauration de la fonction cardiaque, délétère dans le développement de l’insuffisance cardiaque.
En utilisant le modèle de souris invalidées pour le gène du récepteur ET-B, D. Ivy et al. [28] ont montré que ce récepteur était impliqué dans la modulation du tonus vasculaire pulmonaire et en réponse à une hypoxie aiguë.
Rôle dans l’angiogenèse
La membrane chorio-allantoïdienne (CAM) de poulet est un modèle expérimental très utilisé dans le domaine de l’angiogenèse. Il a été montré qu’une libération continue d’ET-1 sur la CAM induisait la formation d’un réseau vasculaire entraînant une augmentation du nombre de vaisseaux [29]. Dans un autre modèle d’angiogenèse induite par ischémie de l’artère fémorale chez le rat, ET-1 seule n’a aucun effet. En revanche, le blocage des récepteurs ET par le bosentan (antagoniste des récepteurs de l’endothéline) induit un effet proangiogénique [30].
Rôle dans la reproduction
De fortes concentrations d’ECE-1 ont été détectées dans les cellules de Leydig, les cellules de la granulosa et de la thèque [6]. F.Miceli etal. [31] ont montré que les cellules lutéales produisaient de l’ET et contrôlaient la production de progestérone induite par l’hCG (human choriogonadotrophin) via les récepteurs ET-A, suggérant une interaction entre la progestérone, les prostaglandines et l’ET dans la physiologie du corpus luteum (corps jaune).
Rôles délétères du système endothéline
Hypertension
Le rôle des ET dans l’hypertension n’est pas clairement défini. Récemment, chez certains patients modérément hypertendus, une augmentation de l’ARNm codant pour la PPET-1 dans l’endothélium des artères de résistance a été observée [32].
Un polymorphisme G/T dans l’exon 5 du gène codant pour la PPET-1, substituant une Lys en Asn en position 198, a été associé à une augmentation du risque d’hypertension artérielle [33]. L’impact de ce polymorphisme sur un changement de la réactivité vasculaire dans les artères mammaires de patients hospitalisés pour un pontage a été étudié. Les patients porteurs de l’allèle T ont une potentialisation augmentée de la réponse à la phényléphrine par l’endothéline [34].
Athérosclérose
Dans cette maladie, ET-1 active les récepteurs ET-A présents sur les macrophages, les cellules musculaires lisses et les fibroblastes. La synthèse d’ET-1 est stimulée par les LDL (low density lipoproteins) oxydées dans les cellules endothéliales [35], les macrophages [36] et les cellules musculaires lisses d’artères coronaires [37]. Une coexpression d’ECE-1 et d’ET-1, corrélée à l’évolution des plaques d’athérosclérose, a été observée dans les artères humaines [38].
Rôle dans l’angiogenèse tumorale
L’angiogenèse tumorale nécessite des facteurs angiogéniques, tels que le VEGF (vascular endothelial growth factor), produits par les cellules cancéreuses pour affecter le tissu hôte [39]. L’ET-1 et ses récepteurs sont des modulateurs négatifs de la réponse angiogénique dans la progression du cancer du côlon, en réprimant la différenciation des fibroblastes et en induisant l’apparition des récepteurs ET-B [40].
Rôle dans la maladie d’Alzheimer
Dans toutes les formes de la maladie d’Alzheimer, on retrouve un dépôt de peptides β-amyloïdes dans le cerveau. Il a été montré que l’ECE-1 était une nouvelle enzyme de dégradation des peptides β-amyloïdes [41].
Conclusions
Beaucoup de connaissances ont été accumulées sur le système endothéline depuis la découverte du peptide ET-1. La disponibilité d’agonistes et d’antagonistes sélectifs des deux types de récepteurs, et de modèles animaux génétiquement modifiés, a permis de montrer l’importance du système endothéline. Ce système n’est pas seulement impliqué dans le système cardiovasculaire mais dans beaucoup d’autres domaines, comme cet article non exhaustif s’est efforcé de le montrer.
Parties annexes
Références
- 1. Yanagisawa M, Kurihara H, Kimura S, et al. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 1988 ; 332 : 411-5.
- 2. Miyauchi T, Masaki T. Pathophysiology of endothelin in the cardiovascular system. Annu Rev Physiol 1999 ; 61 : 391-415.
- 3. Hooper NM. Families of zinc metalloproteases. FEBS Lett 1994 ; 354 : 1-6.
- 4. Xu D, Emoto N, Giaid A, et al. ECE-1 : a membrane-bound metalloprotease that catalyzes the proteolytic activation of big endothelin-1. Cell 1994 ; 78 : 473-85.
- 5. Emoto N, Yanagisawa M. Endothelin-converting enzyme-2 is a membrane-bound, phosphoramidon-sensitive metalloprotease with acidic pH optimum. J Biol Chem 1995 ; 270 : 15262-8.
- 6. Korth P, Bohle RM, Corvol P, Pinet F. Cellular distribution of endothelin-converting enzyme-1 in human tissues. J Histochem Cytochem 1999 ; 47 : 447-62.
- 7. Parnot C, Le Moullec JM, Cousin MA, et al. A live-cell assay for studying extracellular and intracellular endothelin-converting enzyme activity. Hypertension 1997 ; 30 : 837-44.
- 8. Yanagisawa H, Hammer RE, Richardson JA, et al. Disruption of ECE-1 and ECE-2 reveals a role for endothelin-converting enzyme-2 in murine cardiac development. J Clin Invest 2000 ; 105 : 1373-82.
- 9. Valdenaire O, Lepailleur-Enouf D, Egidy G, et al. A fourth isoform of endothelin-converting enzyme (ECE-1) is generated from an additional promoter molecular cloning and characterization. Eur J Biochem 1999 ; 264 : 341-9.
- 10. Valdenaire O, Barret A, Schweizer A, et al. Two di-leucine-based motifs account for the different subcellular localizations of the human endothelin-converting enzyme (ECE-1) isoforms. J Cell Sci 1999 ; 112 : 3115-25.
- 11. Johnson GD, Stevenson T, Ahn K. Hydrolysis of peptide hormones by endothelin-converting enzyme-1. A comparison with neprilysin. J Biol Chem 1999 ; 274 : 4053-8.
- 12. Kedzierski RM, Yanagisawa M. Endothelin system : the double-edged sword in health and disease. Annu Rev Pharmacol Toxicol 2001 ; 41 : 851-76.
- 13. Egidy G, Baviera E, Ciuffo G, et al. Localization of the endothelin system in aldosterone-producing adenomas. Hypertension 2001 ; 38 : 1137-42.
- 14. Yokota J, Kawana M, Hidai C, et al. Retinoic acid suppresses endothelin-1 gene expression at the transcription level in endothelial cells. Atherosclerosis 2001 ; 159 : 491-6.
- 15. Wright MF, Sayre B, Keith Inskeep EK, Flores JA. Prostaglandin F(2alpha) regulation of the bovine corpus luteum endothelin system during the early and midluteal phase. Biol Reprod 2001 ; 65 : 1710-7.
- 16. Quehenberger P, Exner M, Sunder-Plassmann R, et al. Leptin induces endothelin-1 in endothelial cells in vitro. Circ Res 2002 ; 90 : 711-8.
- 17. Hasselblatt M, Lewczuk P, Loffler BM, et al. Role of the astrocytic ET(B) receptor in the regulation of extracellular endothelin-1 during hypoxia. Glia 2001 ; 34 : 18-26.
- 18. Yamashita K, Discher DJ, Hu J, et al. Molecular regulation of the endothelin-1 gene by hypoxia. Contributions of hypoxia-inducible factor-1, activator protein-1, GATA-2, and p300/CBP. J Biol Chem 2001 ; 276 : 12645-53.
- 19. Aitsebaomo J, Kingsley-Kallesen ML, Wu Y, et al. Vezf1/DB1 is an endothelial cell-specific transcription factor that regulates expression of the endothelin-1 promoter. J Biol Chem 2001 ; 276 : 39197-205.
- 20. Kurihara Y, Kurihara H, Oda H, et al. Aortic arch malformations and ventricular septal defect in mice deficient in endothelin-1. J Clin Invest 1995 ; 96 : 293-300.
- 21. Yanagisawa H, Yanagisawa M, Kapur RP, et al. Dual genetic pathways of endothelin-mediated intercellular signaling revealed by targeted disruption of endothelin converting enzyme-1 gene. Development 1998 ; 125 : 825-36.
- 22. Clouthier DE, Williams SC, Yanagisawa H, et al. Signaling pathways crucial for craniofacial development revealed by endothelin-A receptor-deficient mice. Dev Biol 2000 ; 217 : 10-24.
- 23. Baynash AG, Hosoda K, Giaid A, et al. Interaction of endothelin-3 with endothelin-B receptor is essential for development of epidermal melanocytes and enteric neurons. Cell 1994 ; 79 : 1277-85.
- 24. Hachiya A, Kobayashi T, Takema Y, Imokawa G. Biochemical characterization of endothelin-converting enzyme-1alpha in cultured skin-derived cells and its postulated role in the stimulation of melanogenesis in human epidermis. J Biol Chem 2002 ; 277 : 5395-403.
- 25. Kempf H, Linares C, Corvol P, Gasc JM. Pharmacological inactivation of the endothelin type A receptor in the early chick embryo : a model of mispatterning of the branchial arch derivatives. Development 1998 ; 125 : 4931-41.
- 26. Laghmani K, Preisig PA, Moe OW, et al. Endothelin-1/endothelin-B receptor-mediated increases in NHE3 activity in chronic metabolic acidosis. J Clin Invest 2001 ; 107 : 1563-9.
- 27. Peng Y, Amemiya M, Yang X, et al. ET(B) receptor activation causes exocytic insertion of NHE3 in OKP cells. Am J Physiol Renal Physiol 2001 ; 280 : F34-42.
- 28. Ivy D, McMurtry IF, Yanagisawa M, et al. Endothelin B receptor deficiency potentiates ET-1 and hypoxic pulmonary vasoconstriction. Am J Physiol Lung Cell Mol Physiol 2001 ; 280 : L1040-8.
- 29. Cruz A, Parnot C, Ribatti D, et al. Endothelin-1, a regulator of angiogenesis in the chick chorioallantoic membrane. J Vasc Res 2001 ; 38 : 536-45.
- 30. Iglarz M, Silvestre JS, Duriez M, et al. Chronic blockade of endothelin receptors improves ischemia-induced angiogenesis in rat hindlimbs through activation of vascular endothelial growth factor-no pathway. Arterioscler Thromb Vasc Biol 2001 ; 21 : 1598-603.
- 31. Miceli F, Minici F, Garcia PM, et al. Endothelins enhance prostaglandin (PGE2 and PGF2alpha) biosynthesis and release by human luteal cells : evidence of a new paracrine/autocrine regulation of luteal function. J Clin Endocrinol Metab 2001 ; 86 : 811-7.
- 32. Schiffrin EL. Role of endothelin-1 in hypertension and vascular disease. Am J Hypertens 2001 ; 14 : 83S-9.
- 33. Tiret L, Poirier O, Hallet V, et al. The Lys198Asn polymorphism in the endothelin-1 gene is associated with blood pressure in overweight people. Hypertension 1999 ; 33 : 1169-74.
- 34. Iglarz M, Benessiano J, Philip I, et al. Preproendothelin-1 gene polymorphism is related to a change in vascular reactivity in the human mammary artery in vitro. Hypertension 2002 ; 39 : 209-13.
- 35. Boulanger CM, Tanner FC, Bea ML, et al. Oxidized low density lipoproteins induce mRNA expression and release of endothelin from human and porcine endothelium. Circ Res 1992 ; 70 : 1191-7.
- 36. Martin-Nizard F, Houssaini HS, Lestavel-Delattre S, et al. Modified low density lipoproteins activate human macrophages to secrete immunoreactive endothelin. FEBS Lett 1991 ; 293 : 127-30.
- 37. Haug C, Schmid-Kotsas A, Zorn U, et al. Endothelin-1 synthesis and endothelin B receptor expression in human coronary artery smooth muscle cells and monocyte-derived macrophages is up-regulated by low density lipoproteins. J Mol Cell Cardiol 2001 ; 33 : 1701-12.
- 38. Ihling C, Szombathy T, Bohrmann B, et al. Coexpression of endothelin-converting enzyme-1 and endothelin-1 in different stages of human atherosclerosis. Circulation 2001 ; 104 : 864-9.
- 39. Folkman J. Tumor angiogenesis : therapeutic implications. N Engl J Med 1971 ; 285 : 1182-6.
- 40. Egidy G, Juillerat-Jeanneret L, Jeannin JF, et al. Modulation of human colon tumor-stromal interactions by the endothelin system. Am J Pathol 2000 ; 157 : 1863-74.
- 41. Eckman EA, Reed DK, Eckman CB. Degradation of the Alzheimer’s amyloid beta peptide by endothelin-converting enzyme. J Biol Chem 2001 ; 276 : 24540-8.
Liste des figures
Figure 1
Genèse des endothélines.
Les précurseurs des endothélines sont clivés par deux groupes de protéases pour produire les endothélines matures et actives. Les préproendothélines (prépro ET-1, -2, -3) constituées d’environ 200 résidus sont clivés au niveau de sites dibasiques par des endopeptidases de type furine pour former des intermédiaires biologiquement inactifs appelés big endothélines (big ET-1, 2, 3) qui sont des peptides de 37 à 41 acides aminés. Les big endothélines sont ensuite clivées au niveau de la liaison Trp-Val de la big ET-1 et de la big ET-2 ou au niveau de la liaison Trp-Ile de la big ET-3 pour former le peptide final de 21 acides aminés (ET-1, 2). Cette dernière étape est effectuée par des métalloprotéases membranaires à zinc de la superfamille des néprilysines [3], appelées enzymes de conversion de l’endothéline (ECE-1, 2). Les peptides endothélines (ET-1, 2, 3) produits interagissent de manière sélective avec le récepteur ET-A et de manière non sélective avec le récepteur ET-B. Ces deux récepteurs sont couplés à des protéines G variées (Gq, Gs, Gi) permettant différentes réponses physiologiques. L’affinité du récepteur ET-A pour les peptides ET-1 et ET-2 est de l’ordre de la nanomole (flèches en trait plein) et celle pour ET-3 est plus faible de deux ordres de grandeur (flèche en trait pointillé). En revanche, le récepteur ET-B a la même affinité pour les trois peptides (de l’ordre de la nanomole).
Figure 2
Isoformes de l’enzyme de conversion humaine de l’endothéline (ECE-1).
ECE-1 est codée par un seul gène qui subit un épissage alternatif permettant la création de quatre isoformes, appelées a, b, c et d. A. Schéma de la structure partielle du gène ECE-1. Seuls quatre premiers exons du gène sont représentés (1c, 1b, 2, 3). La position du site d’initiation de la traduction est indiquée (atg). Les ellipses bleues représentent les isoformes du promoteur (P) présomptif du gène. B. Production des ARN messagers ECE-1a, 1b, 1c, 1d par épissage alternatif. Pour la clarté du schéma, seuls les traits en pointillé indiquant l’épissage des formes ECE-1a et 1b sont figurés. C. Séquence amino-terminale des quatre isoformes. Pour ECE-1b, 1c et 1d, les traits en pointillé indiquent l’identité des résidus avec la ligne précédente. Les résidus en gras correspondent à la séquence codée par l’exon 3 pour ECE-1a, par l’exon 1b pour ECE-1b, par l’exon 1c pour ECE-1c, et par l’exon 2 pour ECE-1d.
Liste des tableaux
Tableau I
Profil d’expression des principaux constituants du système endothéline.
Le signe + indique la présence du composant, le signe - son absence. ND : non déterminé. La distribution d’ET-2, d’ET-3 et d’ECE-2 n’est pas indiquée car ces constituants n’ont pas été étudiés de manière exhaustive.
Tableau II
Fonctions in vivo proposées pour les principaux constituants du système endothéline.
La présence d’un point d’interrogation correspond à un rôle possible; ++ signifie essentiellement.