Corps de l’article
Voilà deux ans qu’une nouvelle molécule, l’activation-induced cytidine deaminase (AID) était décrite et son rôle dans la maturation du répertoire des anticorps clairement démontré. En effet, AID est indispensable, aussi bien chez l’homme que chez la souris, aux deux événements majeurs qui surviennent au cours de la différenciation terminale des lymphocytes B, dans les centres germinatifs des organes lymphoïdes secondaires: la commutation isotypique et la création des mutations somatiques.
Les immunoglobulines M (IgM) qui portent une chaîne lourde μ, représentent la première classe d’anticorps synthétisée par les lymphocytes B quand ils se différencient en plasmocytes. Dans le gène codant pour les immunoglobulines, les régions constantes (définissant les différents isotypes) sont organisées de telle façon que l’exon μ est le premier en 5’, suivi de l’exon δ, puis γ, ε, et finalement α. La commutation isotypique consiste en la substitution de la chaîne lourde (H) de l’IgM par celle d’un autre isotype (IgG, IgA, IgE) sans que la spécificité antigénique (portée par la région variable [V] des immunoglobulines) soit modifiée. Cet événement nécessite la recombinaison somatique entre deux régions S (switch) situées en amont de chaque région constante CH (constant heavy) des chaînes lourdes des immunoglobulines.
Le second événement, l’introduction de mutations somatiques dans la région V, est un préalable nécessaire à la sélection positive des cellules B présentant un récepteur de l’antigène (BCR) de forte affinité. Les cellules B exprimant un récepteur de faible affinité, ou bien reconnaissant un auto-antigène, sont sélectionnées négativement et meurent par apoptose.
Des mutations du gène codant pour AID sont responsables chez l’homme d’un syndrome d’hyper-IgM (HIGM2) caractérisé par un défaut de commutation isotypique des immunoglobulines et une absence de production des mutations somatiques [1] ((→) m/s 2000, n° 10, p. 1142). Ce phénotype est retrouvé chez les souris dont le gène AID a été inactivé par recombinaison homologue [2]. Pour la première fois, on mettait en évidence un mécanisme commun, dépendant de AID, aux deux processus que représentent la commutation isotypique et la création de mutations somatiques. L’observation, dans les régions de switch S, de mutations somatiques comparables à celles que portent les régions V suggérait elle aussi qu’une voie commune était utilisée dans ces deux événements de la maturation terminale des lymphocytes B [3].
À quel niveau agit AID?
AID agit probablement lors d’un événement moléculaire commun à la commutation isotypique et à la création des mutations somatiques. Au cours de la commutation isotypique, on sait qu’une première étape d’activité transcriptionnelle a lieu dans la région I-CH (intronic-constant heavy chain) aboutissant à la formation de transcrits stériles; cette première étape de transcription est également requise lors de la production des mutations somatiques. Cette activité n’est pas altérée en l’absence de AID comme en témoigne la détection de transcrits stériles [1, 2].
La formation de cassures double-brins de l’ADN est nécessaire à la commutation isotypique afin que la recombinaison entre deux régions S puisse survenir [4]. Cette cassure double-brins est soit « primitive », soit la résultante de cassures « simple-brin » situées sur les deux brins de l’ADN. Plusieurs groupes ont récemment décrit la formation de cassures double-brins de l’ADN dans les régions V au cours de la création des mutations somatiques [5-8], mais ces observations sont de plus en plus controversées et apparaissent non spécifiques [7, 9].
Or, les foyers de réparation des cassures de l’ADN comprenant l’histone H2AX phosphorylée et la protéine de réparation nibrine, qui sont normalement présentes dans les régions S au cours de la commutation isotypique, ne sont pas détectés dans des lymphocytes B de souris déficientes en AID [10]. Ce résultat suggère que AID intervient dans l’induction des cassures double-brins de l’ADN au cours de la commutation isotypique.
Mode d’action de AID
Si l’on ne peut pas affirmer aujourd’hui que le point commun entre commutation isotypique et mutations somatiques est le passage par une étape de cassure double-brins de l’ADN, il est en revanche établi que, dans les deux cas, des altérations de l’ADN sont nécessaires au bon déroulement de ces événements.
Le mode d’action de AID n’est pas encore connu: du fait de son homologie avec APOBEC1, une enzyme capable d’« éditer » l’ARN[*] [11], et de son activité de désamination des cytidines in vitro, l’hypothèse initiale était que AID « éditait » un ARN commun à ces deux événements de la maturation terminale des lymphocytes B. AID pourrait ainsi éditer un ARN codant pour une endonucléase impliquée dans les altérations de l’ADN (Figure 1). Si comme APOBEC1, AID n’est active qu’avec l’aide de co-facteurs, ceux-ci, de même que son substrat, ne sont pas exprimés uniquement dans les lymphocytes B puisque la transfection de AID dans des fibroblastes est nécessaire et suffisante pour induire la commutation isotypique de substrats artificiels appropriés [12].
Figure 1
Hypothèses sur le mode d’action de l’activation-induced cytidine deaminase (AID).
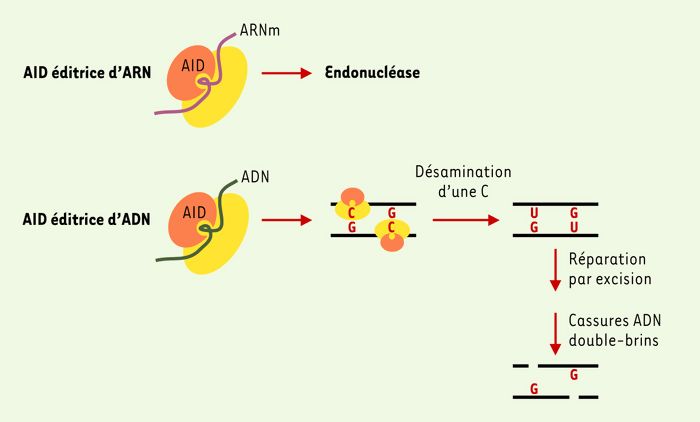
Une seconde hypothèse prévaut actuellement: il s’agit en fait d’une action directe de AID sur l’ADN. L’enzyme, par son activité de désamination des cytidines, entraînerait la formation de mésappariements nucléotidiques qui à leur tour déclencheraient une cascade d’événements de réparation de l’ADN. En effet, des données récentes obtenues in vitro par transfection de AID dans E. coli démontrent que AID peut créer des mésappariements nucléotidiques [13]. De plus, l’observation par le même groupe qu’une souris dont le gène codant pour UNG (une molécule capable d’exciser les mésappariements causés par la désamination des cytidines, créant de ce fait des cassures simple-brin de l’ADN) a été inactivé par recombinaison homologue présente un défaut de commutation isotypique des immunoglobulines et un biais dans la production des mutations somatiques conforte cette hypothèse [14]. Ces données permettent de proposer le schéma suivant (Figure 1).
AID, par son activité désaminase, modifierait au niveau de l’ADN des cytidines en uridines dans les régions S et dans les régions V. UNG interviendrait alors en excisant ces uridines mésappariées, créant par là même des cassures simple-brin de l’ADN. Si, au cours de la commutation isotypique, ces cassures étaient nombreuses et situées sur les deux brins, elles aboutiraient à des cassures double-brins de l’ADN. Une machinerie de recombinaison et de réparation serait alors recrutée et activée, utilisant probablement, outre des molécules intervenant dans la recombinaison non homologue (NHEJ, non-homologous end joining), des molécules de la réparation plus spécifiques de la commutation isotypique. Dans le processus de création des mutations somatiques, les cassures simple-brin de l’ADN créées par AID seraient immédiatement réparées par le recrutement de molécules favorisant l’incorporation d’erreurs (incluant la polymérase non fidèle Pol iota) [15].
Cependant, le rôle direct de AID sur l’ADN est remis en question par l’observation selon laquelle APOBEC-1, une enzyme dont on sait qu’elle est capable d’éditer l’ARN, peut aussi avoir une activité sur l’ADN lorsqu’elle est surexprimée dans E. coli [16]. Plusieurs points obscurs subsistent.: AID édite-t-elle l’ARN ou l’ADN? Existe-t-il des co-facteurs de AID? Quels sont les mécanismes qui permettent de cibler son activité dans les régions S et V des gènes des immunoglobulines? Comment différents systèmes de réparation et/ou de recombinaison sont-ils recrutés en fonction de la région altérée?
AID est le sujet de recherche de nombreuses équipes et il est vraisemblable que l’activité précise de AID sera clarifiée dans un avenir proche, ce qui permettra de préciser les mécanismes responsables de la maturation et la diversification du répertoire des anticorps.
Parties annexes
Notes
-
[*]
L’editing représente tout « traitement » de l’ARNm (ajout, suppression, substitution d’un ou plusieurs nucléotides) aboutissant à un ARNm dont la séquence diffère de celle du brin de l’ADN sens.
Références
- 1. Revy P, Muto T, Levy Y, et al. Activation-induced cytidine deaminase (AID) deficiency causes the autosomal recessive form of the hyper-IgM syndrome (HIGM2). Cell 2000; 102: 565-75.
- 2. Muramatsu M, Kinoshita K, Fagarasan S, Yamada S, Shinkai Y, Honjo T. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell 2000; 102: 553-63.
- 3. Nagaoka H, Muramatsu M, Yamamura N, Kinoshita K, Honjo T. Activation-induced deaminase (AID)-directed hypermutation in the immunoglobulin Smu region: implication of AID involvement in a common step of class switch recombination and somatic hypermutation. J Exp Med 2002; 195: 529-34.
- 4. Wuerffel RA, Du J, Thompson RJ, Kenter AL. IgS gamma3 DNA-specifc double strand breaks are induced in mitogen-activated B cells and are implicated in switch recombination. J Immunol 1997; 159: 4139-44.
- 5. Bross L, Fukita Y, McBlane F, Demolliere C, Rajewsky K, Jacobs H. DNA double-strand breaks in immunoglobulin genes undergoing somatic hypermutation. Immunity 2000; 13: 589-97.
- 6. Papavasiliou FN, Schatz DG. Cell-cycle-regulated DNA double-stranded breaks in somatic hypermutation of immunoglobulin genes. Nature 2000; 408: 216-21.
- 7. Bross L, Muramatsu M, Kinoshita K, Honjo T, Jacobs H. DNA double-strand breaks: prior to but not sufficient in targeting hypermutation. J Exp Med 2002; 195: 1187-92.
- 8. Papavasiliou FN, Schatz DG. The activation-induced deaminase functions in a postcleavage step of the somatic hypermutation process. J Exp Med 2002; 195: 1193-8.
- 9. Faili A, Aoufouchi S, Gueranger Q, et al. AID-dependent somatic hypermutation occurs as a DNA single-strand event in the BL2 cell line. Nat Immunol 2002; 3: 815-21.
- 10. Petersen S, Casellas R, Reina-San-Martin B, et al. AID is required to initiate Nbs1/gamma-H2AX focus formation and mutations at sites of class switching. Nature 2001; 414: 660-5.
- 11. Blanc V, Farré JC, Litvak S, Araya A. Réécriture du matériel génétique: fonctions et mécanismes de l’édition de l’ARN. Med Sci 2002; 18: 181-92.
- 12. Okazaki IM, Kinoshita K, Muramatsu M, Yoshikawa K, Honjo T. The AID enzyme induces class switch recombination in fibroblasts. Nature 2002; 416: 340-5.
- 13. Petersen-Mahrt SK, Harris RS, Neuberger MS. AID mutates E. coli suggesting a DNA deamination mechanism for antibody diversification. Nature 2002; 418: 99-104.
- 14. Rada C, Williams GT, Nilsen H, Barnes DE, Lindahl T, Neuberger MS. Immunoglobulin isotype switching is inhibited and somatic hypermutation perturbed in UNG-deficient mice. Curr Biol 2002; 12: 1748-55.
- 15. Faili A, Aoufouchi S, Flatter E, Gueranger Q, Reynaud CA, Weill JC. Induction of somatic hypermutation in immunoglobulin genes is dependent on DNA polymerase iota. Nature 2002; 419: 944-7.
- 16. Harris RS, Petersen-Mahrt SK, Neuberger MS. RNA editing enzyme apobec1 and some of its homologs can act as DNA mutators. Mol Cell 2002; 10: 1247-53.
Liste des figures
Figure 1
Hypothèses sur le mode d’action de l’activation-induced cytidine deaminase (AID).