Résumés
Résumé
Cette étude porte sur la dynamique, depuis le début du 20e siècle, d’une forêt de reboisement à pin noir d’Autriche (Pinus nigra ssp. nigricans) sur un talus d’éboulis, localisé dans le sud du massif des Grandes Rousses (Alpes du nord, France). Les différentes étapes de la dynamique forestière ainsi que la croissance des arbres étudiée par dendrologie révèlent les fluctuations climatiques du siècle écoulé (1896-2003). Une cartographie diachronique à grande échelle (1/5 000) au pas de temps bi-décennal, combinée à des prélèvements dendrologiques (157 arbres échantillonnés sur 12 placettes), font ressortir l’expansion rapide de la pinède entre 1950 et 1970, puis un ralentissement dans les décennies 1980, 1990 et le début des années 2000. Cette évolution concorde avec les variations des basses fréquences contenues dans les séries dendrochronologiques et météorologiques. La croissance du peuplement dans les décennies 1950-1970 coïncide avec une augmentation de 40 % de la croissance radiale et une succession d’étés frais et arrosés, d’après les séries climatiques de la station Besse en Oisans située à proximité du site. Inversement, depuis le milieu des années 1970, le peuplement connaît un déficit de croissance radiale de 20 % lié à une série d’étés chauds et secs, contribuant à une expansion ralentie. Cette sensibilité très forte des pins à la sécheresse pré-estivale (mai, juin et juillet) est confirmée, au pas de temps mensuel, par l’analyse dendroclimatologique. Dans un contexte d’épisodes de sécheresse intra-alpine de plus en plus prononcé, un phénomène récent encore peu abordé par les scénarios macroclimatiques, ces résultats conduisent à une série d’interrogations sur le devenir de ces peuplements et sur la fonction de sentinelle de ces forêts reboisées sur un substrat à faible capacité hydrique, capables d’enregistrer fortement les modifications des régimes pluviométriques et thermiques, encore mal modélisées en régions de montagnes.
Abstract
Forest dynamics were studied on a scree slope located at southern end of the Grandes Rousses Mountains (Northern Alps, France), which had been reforested with Austrian black pine (Pinus nigra ssp. nigricans) at the beginning of the 19th century. Dendrological study of the stages of forest dynamics and tree growth revealed climate fluctuations over the last century. Large scale (1 :5 000) bidecennial diachronic cartography combined with dendrologic samples (157 trees sampled on 12 plots) showed rapid expansion of Pinus nigra between 1950 and 1970, followed by a slow-down since the beginning of the 1980s. This tendency matches low-frequency variations in both dendrochronological and meteorological series. A rise in tree numbers between the 1950s and 1970s thus parallels an increase of 40 % in radial growth and a sequence of cool, wet summers recorded at the nearby meteorological station of Besse en Oisans. In the same way, radial growth has decreased by 20 % since the middle of the 1970s, in relation with a succession of hot, dry summers, resulting in a slow-down in colonization. The high sensitivity of pines to early summer drought (in May, June and July) was corroborated by monthly dendroclimatologic analysis. These results raise questions about the future survival of such pine populations in the context of increasingly-severe intra-alpine drought, a phenomenon as yet insufficiently predicted by macroclimatic scenarios. Further research is also needed into the suitability of protection forests on scree slopes to detect modifications in the pluviometric patterns of mountain regions, currently poorly predicted by climatic models.
Corps de l’article
Introduction
Sous l’influence du réchauffement climatique, les aires de répartition et la productivité des arbres forestiers évoluent. Les exemples les plus fréquemment cités sont les déplacements des limites latitudinales et altitudinales des arbres (Körner, 1994 ; Solomon et Kirilenko, 1997 ; Mac Donald et al., 1998), l’extension ou la contraction de l’aire d’occupation des espèces ou les fluctuations des populations (Theurillat, 1995). L’accélération de la croissance radiale des arbres au cours du 20e siècle a également fait l’objet de résultats concordants dans tout l’hémisphère nord (Lamarche et al., 1984 ; Diaz etal., 1997). Selon les espèces et les régions, l’origine de cette accélération remonte de quelques dizaines d’années à un peu plus d’un siècle. En Europe, cette augmentation de l’accroissement annuel moyen des arbres a été notée, tous types de traitements sylvicoles confondus (Spiecker et al., 1996). En France, elle a été constatée sur le chêne pubescent (Quercus pubescens Willd. ; Becker et al., 1994 ; Rathgeber et al., 1999), le hêtre (Fagus sylvatica L. ; Badeau et al., 1995), le pin laricio (Pinus laricio Poir. ; Lebourgeois et Becker, 1996), le pin d’Alep (Pinus halepensis Mill. ; Vennetier et Hervé, 1999). Dans les Alpes françaises du nord, elle concerne le mélèze (Larix decidua Mill. ; Belingard, 1996 ; Petitcolas, 1998), le sapin pectiné (Abies alba Mill. ; Rolland, 1993), l’épicéa (Picea abies (L.) Kars. ; Desplanque, 1997) et le pin à crochets (Pinus uncinata Mill. Ex Mirb ; Rolland, 1996 ; Petitcolas, 1998).
Les résultats obtenus montrent que des réactions significatives de la croissance radiale se produisent dans les zones de bordure des écosystèmes où les peuplements poussent près des limites de leur tolérance par rapport à certains facteurs du milieu (Hustich, 1983 ; Grace, 1987 ; Di Castri et al., 1988 ; Hansen et al., 1992 ; Holten et Carey, 1992 ; Kullman, 1993, 1997 ; Guisan et al., 1995 ; Didier et Brun, 1998 ; Jobaggy et Jackson, 2000 ; Moen et al., 2004). Ainsi, les transitions entre les biomes aux hautes latitudes (Holtmeier, 1973 ; Lavoie et Payette, 1994 ; Guisan et al., 1995), et surtout les limites supraforestières aux hautes altitudes (Graybill, 1987 ; Graumlich et al., 1989 ; De Beaulieu et al., 1990 ; Peterson et al., 1990 ; Belingard et Tessier, 1993 ; Nicolussi et al., 1995 ; Belingard et al., 1998 ; Didier et Brun, 1998), ont été utilisées comme bio-indicateurs dans l’étude des changements climatiques. Quelques travaux ont mis en évidence le rôle du réchauffement climatique sur la colonisation végétale arborée. Ils montrent, dans certaines chaînes de montagnes américaines ou européennes, que la fin du 19e et le début du 20e siècle coïncident avec une période d’établissement de jeunes arbres dans la zone subalpine (Heikkinen, 1984 ; Taylor, 1995 ; Didier et Brun, 1998).
En montagne, les paramètres climatiques influencent la physiologie (Kräuchi et al., 2000), la phénologie (Jeanneret et al., 1997), les stratégies de croissance et de développement et la productivité des espèces arborées (Körner, 1994). Ils conditionnent de surcroît la fréquence et l’intensité des processus d’éboulisation (Francou, 1988) qui perturbent, par une action mécanique directe (chutes de blocs) ou indirecte (rajeunissement périodique des profils pédologiques), l’installation des végétaux ligneux. De ce fait, sur les talus d’éboulis, les fluctuations du front forestier résultant de la variation de la fréquence et de l’intensité des processus d’érosion ont été utilisées comme des marqueurs des variations climatiques récentes (Hétu, 1990 ; Lafortune et al., 1997).
La capacité d’intégration des effets du climat par une forêt sur talus d’éboulis est également l’objet de cette étude. L’originalité du talus d’éboulis étudié repose sur la présence d’un reboisement de protection exempt de toute intervention depuis son implantation à la fin du 19e siècle et situé entre 1 000 et 1 500 m d’altitude. Dans cette tranche d’altitude, l’action humaine à travers le défrichement ou l’exploitation pastorale pluriséculaires des versants a pu perturber, plus encore qu’à haute altitude, l’évolution des forêts. L’exode rural généralisé du 20e siècle et la déprise agro-pastorale peuvent brouiller la réponse des écosystèmes vis-à-vis du signal climatique (Didier et Brun, 1998). Le choix de cette forêt de reboisement limite ces biais : d’une part, l’absence d’intervention subséquente en terme de restauration ou d’exploitation forestière est attestée par le service de Restauration des Terrains de Montagne et, d’autre part, la faiblesse des potentialités herbagères garantit des perturbations agro-pastorales limitées. Seules des perturbations naturelles comme les chutes de blocs ou les avalanches ont pu intervenir sur la dynamique de cette forêt. Cependant, les observations (cicatrices d’impact, arbres étêtés, rejets de croissance, etc.) montrent que ces événements n’ont affecté que très ponctuellement la pinède sans influencer, dans son ensemble, le processus de conquête forestière. Dans ces conditions, l’introduction d’espèces arborées correspond à un événement initial à partir duquel vont s’opérer des transformations spontanées (Oldeman, 1990).
L’objectif de l’étude est de mettre en évidence le lien entre le réchauffement climatique, la croissance des arbres plantés et les phases d’évolution de la forêt sur ce talus d’éboulis. Pour atteindre cet objectif, la méthode utilisée repose sur : (1) la mise en évidence des rythmes de colonisation ligneuse, (2) l’analyse des facteurs les plus favorables à la croissance et à la régénération des arbres à travers l’étude des variations temporelles de production ligneuse, (3) le calcul de l’ampleur des dérives climatiques récentes à proximité du site d’étude et (4) l’évaluation du rôle des changements climatiques dans le déplacement des écotones forestiers sur le versant.
Site d’étude
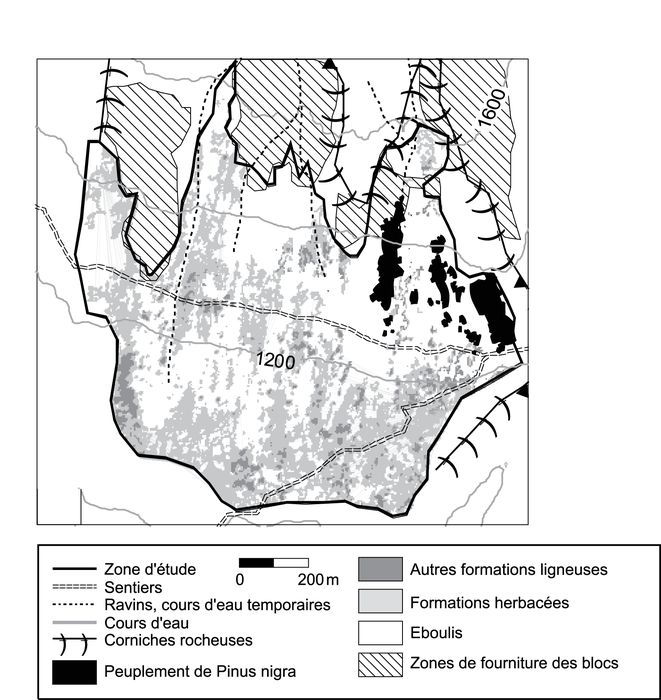
Le talus d’éboulis de la Courbe est situé dans les Alpes françaises, à l’extrémité sud du massif des Grandes Rousses (6° 07’ E, 45° 03’ N ; fig. 1), sur le versant adret de la vallée de la Romanche, dans la commune du Freney d’Oisans. Le climat, de type montagnard continental intra-alpin, est caractérisé par des hivers froids et rigoureux et des étés peu pluvieux, assez chauds. Les moyennes thermiques annuelles collectées à la station météorologique de Besse en Oisans (6° 09’ E, 45° 04’ N, 1 470 m ; fig. 1) atteignent respectivement, pour la période 1959-2003, 1,35 °C, 10,89 °C et 6,1 °C pour les températures minimales, maximales et moyennes. La température minimale moyenne de janvier est de –5,4 °C, tandis que les moyennes maximales de juillet et août avoisinent les 20 °C. D’une longueur de 780 m et d’une largeur de 920 m, le talus d’éboulis de la Courbe est dominé au nord et au nord-est par une corniche rocheuse de 120 m de hauteur, surbaissée (40 m) vers l’ouest. Constituée d’une alternance de gneiss migmatitiques, de brèches, de micaschistes et localement de grès d’âge jurassique, cette corniche subit 156 jours de gel annuel moyen mesurés sur la période 1959-2003 (Thevenon, 1999), particulièrement entre novembre et avril, les six mois où sont concentrés 90 % des jours connaissant au moins un cycle gel-dégel. Pour la période 1930-2003, les précipitations moyennes annuelles sont de 908 mm et le nombre moyen de jours de pluie s’établit à 119. Le coefficient de nivosité s’établit à 31 % pour la période 1951-1960 (Balseinte, 1966). La pluviométrie est caractérisée par un minimum en juillet et un maximum en automne (septembre à novembre). En ce qui concerne la période végétative (de mai à septembre), les totaux pluviométriques s‘élèvent en moyenne à 360 mm, avec 66 % de jours sans averse et 12 % de jours qui ont une température moyenne supérieure à 25 °C. L’angle de continentalité hydrique de Gams, un indicateur de sécheresse intra-alpine (Ozenda, 1985), est assez élevé comme dans le reste des Alpes internes à tendance continentale, soit Artg [précipitations (mm/an)/altitude (m)] = 58°. À propos du phénomène de sécheresse, il faut préciser que le diagramme ombrothermique ne fait pas apparaître de mois sec au sens de Gaussen (Pmm < 2T °C, relation qui définit la sécheresse méditerranéenne ; Ozenda, 1985) mais on note que le mois de juillet est proche de cette définition (Pmoyjuillet = 65 mm et 2 x Tmoy juillet = 30 °C) ; or, depuis une décennie, certaines années et notamment en 2003, les mois de juillet et août, à Besse, sont inclus dans cette catégorie.
Figure 1

Localisation du site d’étude et stratégie d’échantillonnage. Les coordonnées géographiques sont exprimées en latitude/longitude (WGS 84).
Localisation of the study area and the sampling strategy. Map coordinates are in latitude/longitude (WGS 84).
La pente du talus d’éboulis de la Courbe passe de 40° près de la paroi à moins de 28° à la base du versant. La portion nord-est du talus est occupée par la pinède de reboisement sur un peu moins de la moitié de sa longueur (300 m ; fig. 1). Le peuplement constitué de pins noirs d’Autriche s’échelonne entre 1 000 et 1 500 m d’altitude. D’après Allix (1929) et les documents d’archives du service de Restauration des Terrains en Montagne (RTM), trois îlots de pins ont été plantés en 1889, sur une surface de deux hectares, en haut du talus, afin de protéger le hameau des Chazeaux des chutes de blocs et des avalanches (fig. 1). D’après ces documents, aucune plantation ou intervention ultérieure n’a eu lieu. La densité et l’étendue (14 ha) actuelles découlent de la seule conquête spontanée depuis les îlots initiaux au cours du 20e siècle.
La moitié inférieure du talus d’éboulis est occupée par des formations xéro-thermophiles en îlots herbacés et arbustifs, ou par des lanières de pelouses steppiques sub-continentales, landes et fruticées sèches favorisées par la sécheresse estivale intra-alpine, encore renforcée par l’exposition adret du versant. Cette partie inférieure de l’éboulis, à structure grossière et forte porosité, favorable à une sécheresse édaphique importante, semble peu propice à l’expansion du pin noir (fig. 1) et seuls quelques individus, issus des arbres plantés, y sont observés.
Matériel et méthodes
Analyse spatiale de la dynamique de la pinède
La dynamique de la végétation a été étudiée à deux niveaux d’observation. À l’échelle du versant, une cartographie au 1/5 000 des secteurs de colonisation par Pinus nigra a été réalisée d’après les cartes topographiques d’État Major, les plans terriers du service de Restauration des Terrains de Montagne, les photographies anciennes et les photographies aériennes. Elle permet de connaître les rythmes d’évolution du peuplement depuis le début du 20e siècle à des intervalles de 20 ans (1913, 1935, 1952, 1967, 1981 et 2001 ; fig. 2). À l’échelle stationnelle, 12 placettes rectangulaires d’une dimension de 20 m x 20 m (400 m2) ont été cartographiées sur le terrain à l’échelle du 1/100, dont huit dans les îlots de recolonisation et quatre dans le reboisement originel (fig. 1). Cent-soixante-quinze (175) individus au total ont été cartographiés et ont fait l’objet de relevés dendrométriques systématiques (tabl. I).
Figure 2
A
1913

B
1935
C
1952

D
1967

E
1981

F
2001

Répartition du pin noir d’Autriche sur le talus d’éboulis de la Courbe entre 1913 et 2001.
Distribution of Austrian black pine on the scree slope between 1913 and 2001.
Tableau I
Caractéristiques de croissance des 175 pins noirs

N total : nombre de pins par placettes.
N échantillonné : nombre de pins échantillonnés par placettes.
SM : sensibilité moyenne, mesure de la variabilité inter-annuelle.
AC : coefficient d’autocorrélation de premier ordre.
Les deux paramètres, SM et AC, ont été calculés sur les séries brutes de largeur de cernes.
Dendroclimatologie et dendrochronologie
Dans chaque placette, tous les arbres de hauteur supérieure à 100 cm (157 au total) ont été sondés avec une tarière de Pressler, à une hauteur de 50 cm, à raison de deux carottes prises parallèlement à la courbe de niveau afin d’éviter la présence d’éventuel bois de réaction (Belingard etal., 1998). Les anomalies géométriques du tronc ont, dans la mesure du possible, été évitées. Le diamètre des arbres à la hauteur du carottage a également été mesuré. Les séries de cernes ont été mesurées avec une précision théorique de 1/100 mm au moyen d’une table de mesure Lintab (Rinn, 1996). Les séries de cernes ont été interdatées au moyen de deux méthodes complémentaires, tout d’abord à l’aide du logiciel COFECHA (Holmes, 1983) et ensuite, grâce au repérage des cernes diagnostiques sous une loupe binoculaire (Fritts et Swetnam, 1986). La sensibilité moyenne et le coefficient d’autocorrélation d’ordre 1 ont été calculés pour chaque arbre puis, une moyenne a été calculée par placette. Les effets du climat ont été évalués grâce à la méthode des fonctions de réponse (Cook et Kairiukstis, 1990). Cette méthode a été utilisée sur des séries standardisées au moyen d’une moyenne mobile d’ordre 3 qui permet d’éliminer une grande part de l’autocorrélation, de retirer les basses fréquences et d’obtenir des fonctions de réponse significatives (Rolland, 1993 ; Desplanque, 1997 ; Petitcolas, 1998). Les paramètres climatiques mensuels ont été utilisés comme régresseurs et les chronologies indicées comme variables dépendantes (Tessier, 1989 ; Guiot, 1991 ; Petitcolas, 1998). Les réponses ont été calculées au moyen d’un échantillonnage aléatoire avec remises (Efron, 1979). Les totaux mensuels de précipitation (mm) et les températures moyennes (°C) obtenus de la station météorologique de Besse en Oisans ont été utilisés comme paramètres climatiques. Ces données, disponibles sur les périodes 1930-2003 (précipitations) et 1959-2003 (températures), forment la plus longue série climatique existante à proximité du site.
L’étude des basses fréquences des courbes de croissance permet d’éliminer l’effet du signal climatique lié aux variations interannuelles du climat (Rolland, 1993 ; Lebourgeois et Becker, 1996) et de cerner les dérives sur le long terme. À cet effet, les chronologies élémentaires de largeur de cernes ont été standardisées en leur retirant uniquement la tendance d’âge. Pour chaque carotte, les données brutes sont transformées en indices (exprimés en pourcentage) correspondant, pour chaque année de croissance, au rapport des largeurs de cernes observées sur celles prédites par la courbe de tendance d’âge (Erlandsson, 1936 ; Briffa et al., 1992, 1996 ; Grudd et al., 2002).
Ajustement linéaire des données climatiques
Afin d’analyser de possibles dérives dans les données climatiques locales, cinq paramètres disponibles sur la période 1959-2003 ont été utilisés, c’est-à-dire les températures maximales (Tx), minimales (Tn), moyennes (Tm), l’amplitude thermique (Tx–Tn) et les totaux pluviométriques (P). La sécheresse du climat intra-alpin renforcée par la position en adret et la situation sur éboulis, substrat à forte sécheresse édaphique, nous ont également conduits à prendre en compte le déficit hydroclimatique mensuel (P–ETP, où P représente le total pluviométrique mensuel et ETP représente l’évapotranspiration potentielle mensuelle). La formule d’Hargreaves (Hargreaves et Samani, 1982) qui permet d’estimer l’évaporation potentielle mensuelle en prenant en compte les températures mensuelles moyenne, minimale et maximale de l’air a été utilisée :
où Go représente l’irradiation extraterrestre (en cal/cm2/jour), L, la chaleur latente d’évaporation de l’eau (en J/kg), T, la température moyenne mensuelle (en °C), tmax, la moyenne mensuelle des températures maximales (en °C) et tmin, la moyenne mensuelle des températures minimales (en °C).
À partir de ces données mensuelles, des moyennes ont été calculées pour l’année, les saisons et pour la période végétative. Les fluctuations climatiques ont été étudiées au moyen de courbes de tendance linéaire sur la période 1959-2003 et par le calcul de moyennes décennales. Les données thermiques ont été complétées par les températures moyennes saisonnières fournies pour la période 1840-1998 et pour les Alpes occidentales par Böhm et al. (2001). Les coefficients de détermination (r2) obtenus entre les matrices locales et régionales pour la période 1964-1998 (tabl. II) mettent en évidence une corrélation forte entre les températures moyennes estivales des deux séries.
Tableau II
Coefficients de détermination (r2) calculés entre les séries saisonnières de Besse en Oisans et des Alpes occidentales pour la période 1964-1998

Source : Météo France.
Source : Böhm et al., 2001.
Résultats
Augmentation rapide des surfaces boisées entre 1952 et 1967
Les cartes (fig. 2) dessinées d’après les documents d’archives ont permis de reconstituer l’évolution de la surface boisée sur le site au cours du siècle écoulé. Entre 1913 et 2001, le pin noir a colonisé plus de 12,2 ha d’éboulis, passant de 2,25 ha à 14,5 ha. Il occupe ainsi en 2001 plus de 30 % de la surface du dépôt (54 ha).
La carte de 1913 (fig. 2A) révèle que la partie apicale du talus d’éboulis était dépourvue de végétation et que la surface occupée par les espèces ligneuses ne dépassait pas 8 % de la superficie totale du versant, dont 4 % constitué par le reboisement initial (fig. 3). La carte de 1935 (fig. 2B) permet d’apprécier la progression du reboisement 40 ans après la fin des travaux. Pour cette période, l’extension du boisement dans la zone distale est relativement modérée et se limite à l’apparition d’individus isolés ou d’îlots arborés à proximité du peuplement originel. Par contre, la colonisation des parties apicale du versant par la pelouse xérophile apparaît très marquée (fig. 2B). Une expansion importante de la pinède a eu lieu entre 1935 et 1967 (fig. 3). Au cours de cette période, la surface occupée par les pins noirs d’Autriche est multipliée par 4, passant de 2,6 à 10,2 ha. Entre 1935 et 1952 (fig. 2C), la colonisation herbacée rapide de la zone apicale se poursuit. Cette extension s’accompagne d’une densification de la pinède originelle et de la mise en place d’individus isolés et d’îlots de faible densité sur pelouses. La période de 1952-1967 (fig. 2D) constitue le maximum de l’invasion par Pinus nigra. La surface occupée par le pin noir atteint 10,2 ha en 1967 (7 ha en 1952) et les décennies 1950, 1960 et 1970 sont les plus favorables pour l’établissement de jeunes individus (fig. 4). La poursuite de l’extension du peuplement depuis les lisières du reboisement et surtout la densification importante des îlots installés au cours de la période 1935-1952 caractérisent ce pic prolongé de colonisation.
Figure 3

Surfaces occupées par les différentes formations végétales sur le talus d’éboulis de la Courbe entre 1913 et 2001.
Occupied area by vegetation types on the scree slope between 1913 and 2001.
Figure 4

Nombres d’individus établis par décennie au sein des 12 placettes étudiées.
Decennial tree numbers on the 12 sample plots.
La période 1967-2001 (fig. 2E et 2F) marque un ralentissement de la colonisation végétale, amorcée au milieu des années 1970 (fig. 4). À partir de 1981, la surface occupée par les secteurs dénudés se stabilise autour de 10 ha (10,7 ha en 1981 et 10,6 ha en 2001) et la progression des surfaces arborées correspond à la colonisation des pelouses par les ligneux (fig. 3).
Tendance à long terme
Le signal de basse fréquence de la chronologie montre l’augmentation de la croissance radiale (fig. 5). L’ajustement, par une moyenne mobile des séries chronologiques d’indices, permet également de dater cette augmentation relative du début des années 1950 et de l’évaluer à +40 %. On note cependant l’existence d’une période de crise étalée entre 1975-1990 qui se traduit par un déficit relatif de croissance d’environ 20 %.
Figure 5

Évolution de l’indice moyen de croissance radiale (lc %) au cours de la période 1896-2003.
Evolution of the mean radial growth rate (lc %) over the period 1986-2003.
Analyse des fonctions de réponse
La sensibilité moyenne pour le cerne total s’échelonne entre 0,25 et 0,35 (tabl. I). Ces valeurs élevées indiquent des variations interannuelles appréciables dans les chronologies de cernes. Les coefficients d’autocorrélation de premier ordre (AC) atteignent une moyenne de 0,74. Ce coefficient élevé indique une forte dépendance entre le cerne d’une année donnée (n) et le cerne de l’année précédente (n–1).
Pour les années de calibration, la moyenne et l’écart-type atteignent respectivement 0,88 et 0,04 pour les fonctions de réponse calculées sur la matrice températures moyennes/précipitations (fig. 6A), 0,88 et 0,04 pour la matrice températures minimales/précipitations, 0,82 et 0,06 pour la matrice températures maximales/précipitations (fig. 6B) et 0,84 et 0,034 pour le déficit hydrique (fig. 6C). Les coefficients de vérification correspondants atteignent respectivement 0,44 (0,19), 0,48 (0,17), 0,02 (0,2) et 0,59 (0,14). Ces valeurs de vérification élevées (Lebourgeois, 2000) et des réponses significatives au seuil de 99,9 % pour trois des quatre matrices démontrent la validité de la régression estimée. Seules les températures maximales semblent peu influencer la largeur des cernes des années de croissance.
Figure 6
A

B

C

Fonctions de réponse calculées entre la chronologie indicée de largeurs de cernes du Freney et les données climatiques mensuelles (octobre (n–1) à septembre (n)) de la station météorologique de Besse (1959-2003). (A) Totaux pluviométriques et températures mensuelles moyennes. (B) Totaux pluviométriques, températures mensuelles maximales (Tx) et minimales (Tn). (C) Déficit hydrique (P–ETP) obtenu à partir de la formule de Hargreaves. Les valeurs présentées correspondent à un ratio entre les coefficients de régression partiels et leur écart-type, les lignes horizontales indiquent une significativité à des seuils de 90 %, 95 %, 99 % et 99,9 %.
Response function calculated from the tree-ring index at Le Freney and monthly climatic data (October (n–1) to September (n) at Besse Weather Station (1959-2003). (A) Grey bars refer to precipitation-mean monthly temperature matrix. (B) Black bars refer to the precipitation-mean maximum temperature (Tx) matrix, grey bars to the precipitation-mean minimum temperature (Tn) matrix. (C) Grey bars refer to evapotranspiration (P–ETP) as predicted by Hargreaves Evapotranspiration Model. The values shown are the ratio between partial regression coefficient and its standard deviation horizontal lines represent significance thresholds.
Les réponses climatiques mettent en évidence cinq mois significatifs et semblent corrélées plus étroitement aux précipitations qu’aux températures (fig. 6A et 6B). Seules les températures des mois d’avril (n) et juillet (n) paraissent significatives pour la croissance radiale qui paraît, en revanche, très sensible aux précipitations et au déficit hydrique des mois de mai (n), juin (n) et juillet (n). Ces coefficients suggèrent une très forte sensibilité à la sécheresse pré-estivale. Un déficit hydrique au cours des mois de mai (n) et juillet (n) apparaît ainsi défavorable à la croissance radiale (fig. 6C).
Ampleur des variations climatiques récentes
Les amplitudes thermiques (exceptées celles du printemps et du mois de mars en particulier : 0,51 °C/an, r = 0,2, p < 0,01), les précipitations et le déficit hydrique ne montrent pas de dérive significative (tabl. III). Cependant, une augmentation moyenne annuelle des températures maximales (Tx) de l’ordre de 0,039 °C/an (r = 0,48, p < 0,001) et de 0,041 °C/an (r = 0,43, p < 0,001) pour la saison de croissance a été mesurée durant la période 1959-2003 (tabl. III). Cette augmentation a été observée pour tous les mois, mais elle est nettement plus prononcée au cours des mois de mars (0,1 °C/an) à août (0,08 °C/an) (r = 0,46 et 0,48, p < 0,001). L’augmentation des températures minimales (Tn) pour ces mêmes mois est également significative mais légèrement inférieure à celle des températures maximales. Les températures moyennes (Tm) ont augmenté de façon significative au printemps et en été, notamment en mars (0,073 °C/an, r = 0,42, p < 0,001), mai (0,043 °C/an, r = 0,3, p < 0,001), juin (0,042 °C/an, r = 0,3, p < 0,001), juillet (0,042 °C/an, r = 0,34), août (0,062°C/an, r = 0,48, p < 0,001), sur l’année (0,029 °C/an, r = 0,42, p < 0,001) et au cours de la période végétative (0,037 °C/an, r = 0,42, p < 0,001).
Tableau III
Coefficients de régression linéaire (r) et coefficients de détermination (r2) calculés sur les températures mensuelles minimales (Tn), moyennes (Tm), maximales (Tx), sur les amplitudes thermiques (At), le déficit hydrique (P–ETP) (période 1959-2003) et les totaux pluviométriques (Pn) (période 1930-2003) de la station météorologique de Besse en Oisans

Ces régressions linéaires tendent cependant à masquer les rythmes d’évolution des paramètres climatiques qui influencent le plus la croissance des pins : les décennies 1910-1930 et 1950-1980 sont plutôt caractérisées par des étés frais, alors que les années 1940-1950 et 1980-2003 ont connu les étés les plus chauds du siècle dernier (fig. 7). Pour les précipitations moyennes, les décennies 1940-1960 sont caractérisées par une sécheresse printanière marquée (fig. 7). Les déficits pluviométriques printaniers par rapport à la période 1930-2003 (moyenne : 190 mm) atteignent respectivement –45 mm pour 1940-1950 (moyenne : 145 mm) et –35 mm pour 1950-1960 (moyenne : 154 mm).
Figure 7
A

Trait plein en gris : évolution des écarts à la moyenne de la saison estivale calculée pour les Alpes occidentales sur la période 1880-1998 (Böhm et al., 2001) ; trait gras en gris : moyenne mobile calculée sur 10 années.
Thin grey curve : mean summer temperature anomalies shown as deviation in °C from 1880-1998 for the Western Alps (Böhm et al., 2001) ; thick grey curve : 10-year running mean. Thin black curve : mean summer temperature anomalies shown as deviation in °C from 1959-2003 at Besse Weather Station ; thick black curve : 10-year running mean.
B et C

Trait plein en noir : écarts à la moyenne (1930-2003) des totaux pluviométriques mensuels ; trait gras en noir : moyenne mobile calculée sur 10 années.
Thin black curve : precipitation anomalies shown as deviation in mm from the 1930-2003 mean ; thick black curve : 10-year running mean.
Évolution des écarts à la moyenne des températures moyennes estivales (juin, juillet et août) (A) et des totaux pluviométriques mensuels de mai (B) et de juillet (C) à la station météorologique de Besse.
Mean summer temperature anomalies (June, July, August) (A) and mean monthly precipitation anomalies in May (B) and July (C) at Besse Weather Station.
Les déficits estivaux atteignent –15 mm pour les décennies 1940-1950 et 1970-1980 (moyenne : 200 mm) et –25 mm pour la décennie 1980-1990 (moyenne : 190 mm) par rapport à la période 1930-2003 (moyenne : 215 mm). La décennie 1950-1960 est excédentaire de 17 mm (moyenne : 232 mm). Deux périodes paraissent peu favorables à la croissance de Pinus nigra : 1940-1950 et 1980-1990.
Discussion
L’utilisation des méthodes dendroclimatologiques met en évidence, au pas de temps mensuel à annuel, le rôle du stress hydrique printanier et estival sur la croissance radiale des pins noirs d’Autriche. Quelques auteurs ont montré le rôle de la contrainte hydrique à la fois sur la croissance en hauteur et en diamètre des arbres (Aussenac, 1980 ; Badot et al., 1990 ; Granier et al., 1995). Des modèles climatiques simples reliant la croissance radiale et l’indice de stress ont également mis en évidence, pour le hêtre, des variations de croissance radiale de –15 à +15 % pour une variation pluviométrique de l’ordre de –20 à +20 % (Granier et al., 1995). En ce qui concerne les pins, le rôle de la disponibilité en eau des sols, qui prédispose les peuplements aux effets des variations climatiques, plus particulièrement pluviométriques, et qui peut localement accentuer les effets des sécheresses et entraîner des réductions importantes de la croissance radiale, a été mis en évidence dans l’ouest de la France pour Pinus nigra ssp. laricio (Lebourgeois et Becker, 1996). Dans les Alpes, la forte sensibilité de l’arole (Pinus cembra) aux précipitations de juillet a été observée dans le Briançonnais sur versant adret (Petitcolas, 1998). Pour différentes variétés de pins, les effets de la sécheresse estivale sont caractérisés uniquement par une corrélation négative forte entre les températures estivales (n) ou (n–1) et les séries indicées de largeurs de cernes. C’est le cas notamment de Pinus laricio (Lebourgeois, 2000), Pinus sylvestris (Graces et Norton, 1990 ; Richter et al., 1991 ; Briffa, 1992), Pinus taeda (Gregg et al., 1988), Pinus uncinata (Richter et al., 1991 ; Rolland et Schueller, 1996 ; Petitcolas, 1998), Pinus cembra (Kolischuk et Berko, 1967 ; Bednarz, 1981 ; Aniol et Eckstein, 1984 ; Petitcolas, 1998), Pinus mugho (Kolischuk et Berko, 1967 ; Bednarz, 1981) ou Pinus halepensis (Rathgeber, 2002).
Sur l’éboulis de la Courbe, la colonisation végétale depuis les îlots initiaux s’est élargie à l’ensemble de la zone apicale à matériaux fins, mais l’expansion du pin vers les zones distales à matériaux plus grossiers semble difficile. La limite inférieure de cette colonisation, approximativement calée sur la courbe de niveau 1 200 m (fig. 2), suggère l’existence d’un seuil granulométrique distal, à l’origine d’un déficit hydrique estival incompatible avec l’implantation et la survie du pin. Des études physiologiques confirment la sensibilité du pin noir à une trop forte sécheresse édaphique. Elles ont montré que le taux d’assimilation net de Pinus nigra décroissait rapidement lorsque le sol s’asséchait, en raison d’un contrôle rapide et efficace par les stomates de la perte en eau par transpiration, ce qui entraîne une diminution de la production de photosynthates et du stockage des carbohydrates (Aussenac, 1980 ; Lebourgeois et Becker, 1996). Un apport régulier en eau est donc fondamental à la croissance rapide des pins sur ce versant d’autant que la rétention édaphique est faible et que l’évapotranspiration, estivale en particulier, est favorisée par l‘exposition adret.
La réponse positive aux précipitations d’octobre indique les effets de l’année (n–1) sur la croissance au cours de l’année (n). Chez de nombreuses espèces de conifères, les réserves sont construites en automne et stockées tout l’hiver avant que la croissance ne démarre (Aussenac, 1980 ; Petitcolas, 1998). Pinus nigra ne semble ainsi pas capable de fixer, lors d’automnes secs, une quantité suffisante de réserves, ce qui peut causer une réduction de la croissance l’année suivante.
À l’échelle décennale, la forte hausse de productivité du début des années 1950 coïncide avec une série d’étés frais et plutôt humides (fig. 7). Cette période correspond également à une phase de régénération et d’expansion rapide du pin noir d’Autriche (fig. 3 et 4). L’arrivée à maturité des pins issus du reboisement initial de 1889, associée à des conditions climatiques estivales plus favorables à la croissance du pin, explique probablement le pic d’établissement durant cette période.
La réduction de croissance atteint son maximum au milieu des années 1980 et elle correspond à une succession d’étés secs (1979 : –40 % de précipitations par rapport à la moyenne 1930-2003, 1983 : –60 %, 1984 : –45 %, 1986 : –60 % et 1989 : –50 %). Elle s’accompagne d’un ralentissement de la colonisation végétale du versant par Pinus nigra et d’un déficit d’installation important (classe d’âge 15-25 ans, fig. 3 et 4). Ces résultats rejoignent, dans un premier temps, ceux de certains travaux qui ont montré le rôle de la sécheresse sur la régénération, la survie ou la mortalité de différentes espèces arborescentes : Abies alba (Bert, 1993), Quercus rubra (Jenkins et Pallardy, 1995), Picea abies (Mast et al., 1994 ; Mäkinen et al., 2001), Pinus edulis (Ogle et al., 2000), Acer saccharum (Payette et al., 1996), Cornus florida, Acer rubrum (Hanson et al., 2001), Austrocedrus chilensis (Villalba et Veblen, 1997) et Picea koraiensis (Liang et al., 2003).
En revanche, il existe peu de données concernant le seuil de sécheresse susceptible d’empêcher la germination ou d’induire une mortalité des semis ou des individus adultes. Dans le massif de la Harth (Alsace), il semble que la vague de mortalité des chênes durant les années 1980, trois fois supérieure à la moyenne, soit reliée au stress hydrique des années 1983, 1985 et 1989 (Bréda et Pfeiffer, 1999). Sur le versant de la Courbe, une diminution de 50 % des précipitations estivales abaisse de moitié les capacités de régénération du pin noir d’Autriche (fig. 4), sans toutefois entraîner de mortalité accrue chez les individus adultes (absence de troncs morts, pas de recul forestier observé sur le versant). Le déficit démographique au cours des années 1980 peut, dès lors, être lié à une réduction du taux de germination. Les semis ou les jeunes plants souffrent en raison de leur enracinement plus superficiel que celui des individus adultes et de leurs parties aériennes qui se trouvent à proximité du sol. Dans cet environnement, la température dans les premiers mètres au-dessus de la surface du sol et même les premiers centimètres au sein du substrat grossier peut être beaucoup plus élevée que la température de l’air mesurée sous abri (Bréda et al., 2000), surtout dans les portions les plus dénudées de l’éboulis où les plantules ne bénéficient pas de l’ombrage créé par les strates supérieures (Arquès, 2005 ; Corona, 2007). Dans ce contexte, l’interaction entre l’augmentation des températures estivales à partir du début des années 1980 et les sécheresses épisodiques a, fort probablement, affecté le métabolisme photosynthétique et a été létale pour les semis.
Conclusion
La réponse du pin noir d’Autriche au réchauffement climatique ne semble pas aussi linéaire que le laisseraient penser certains modèles climatiques. L’augmentation de la croissance radiale et de l’étendue au cours du siècle écoulé paraît certes importante où la vulnérabilité de l’espèce à l’égard du stress hydrique apparaît nettement au cours des 20 dernières années. Dans ce contexte édaphique contraignant, une poursuite de la hausse thermique à l’origine d’une évapotranspiration accrue aurait, à terme, des conséquences néfastes sur la croissance, la régénération et l’expansion du peuplement. La sécheresse de la période 1975-1990, à l’origine d’une décroissance de la surface terrière de l’ordre de 20 à 30 % et le ralentissement marqué de la colonisation végétale au cours de cette période, témoigne des risques encourus par ces peuplements de protection. Ces risques sont d’autant plus élevés qu’un accroissement de l’assèchement de l’été accompagné d‘une sécheresse accrue est probable aux moyennes latitudes au cours du 21e siècle (GIEC, 2001) que d’une exposition prolongée ou répétée à un stress hydrique qui affaiblit les arbres et favorise l’installation de pathogènes ou de ravageurs secondaires (Kräuchi, 1993 ; MAAPR, 2003).
La combinaison de la cartographie diachronique et des mesures dendrochronologiques permet de suivre avec précision le processus de recolonisation, de cerner ses rythmes et son expansion spatiale au cours du 20e siècle. Dans le contexte de l’éboulis de la Courbe, cette méthode montre clairement que les mosaïques forestières sur éboulis d’adret sont très sensibles aux variations climatiques, à la pluviométrie et au stress hydrique estival. Le suivi de ces mosaïques, surtout dans le cas de forêts de reboisement anciennes sur dépôts détritiques, pourrait être utilisé dans la détection d’une éventuelle modification du régime pluviométrique en régions de montagnes, encore mal modélisée et très variable selon les scénarios climatiques actuellement proposés (GIEC, 2001).
Parties annexes
Remerciements
Les auteurs remercient les personnes qui ont contribué à la préparation de cet article. Les commentaires et suggestions formulés par les évaluateurs critiques, Pierre Pech et Laurent Astrade, complétés par les remarques de Louise Filion, rédactrice associée, de Pierre J.H. Richard, rédacteur en chef, et de Sophie Roberge, secrétaire de rédaction, ont grandement contribué à l’amélioration du manuscrit.
Références
- Allix, A., 1929. Un pays de haute montagne : l’Oisans, étude géographique. Thèse Lettres, Université de Grenoble, 915 p.
- Aniol, R.W. et Eckstein, D., 1984. Dendroclimatological studies at the northern timberline, p. 273-279. In N.A. Mörner et W. Karlén, édit., Climatic Changes on a Yearly to Millennial Basis : Geological, Historical and Instrumental Records. D. Reidel, Dordrecht, 667 p.
- Arquès, S., 2005. Géodynamique, colonisation végétale et phytodiversité des talus d’éboulis dans le massif de la Grande Chartreuse (Préalpes françaises du Nord). Caractéristiques géo-écologiques et sensibilité aux changements environnementaux. Thèse de doctorat, Université de Grenoble, 568 p.
- Aussenac, G., 1980. Comportement hydrique de rameaux excisés de quelques espèces de sapins et de pins noirs en phase de dessiccation. Annales des Sciences Forestières, 37 : 201-215.
- Badeau, V., Dupouey, J.L., Becker, M. et Picard, J.F., 1995. Long-term growth trends of Fagus sylvatica L. in northeastern France. A comparison between high and low density stands. Acta Ecologica, 16 : 571-583.
- Badot, P.M., Perrier, P., Garrec, J.P., Badot, J.M. et Mercier, J., 1990. Impacts des récentes sécheresses et de la pollution atmosphérique dans le dépérissement de l’épicéa dans les forêts jurassiennes. Annales scientifiques de l’Université de Franche-Comté (Besançon), Biologie Écologie, 5 : 43-49.
- Balseinte, R., 1966. Climats montagnards et stations climatiques d’altitude en France. Thèse de doctorat, Université de Grenoble, 520 p.
- Becker, M., Nieminen, T.-M. et Geremia, F., 1994. Short term variations and long term changes in oak productivity in northeastern France. The role of climate and atmospheric CO2. Annales des Sciences Forestières, 51 : 477-492.
- Bednarz, Z., 1981. Relationship of tree-ring widths in the Tatra Mountains to variations in monthly temperature and precipitation. Mitteilungen der Forstlichen Bundesversuchsanstalt, 142 : 357-390.
- Belingard, C. et Tessier, L., 1993. Étude dendroécologique comparée de vieux peuplements de Larix decidua Mill. dans les Alpes Françaises du sud. Dendrochronologia, 11 : 69-78.
- Belingard, C., 1996. Étude dendroécologique de la dynamique de la limite supérieure de la forêt dans les Alpes du Sud, en relation avec les facteurs climatique et anthropique. Thèse de doctorat, Université Aix-Marseille, 103 p.
- Belingard, C., Tessier, L. et Édouard, J.-L., 1998. Reboisement et dynamique naturelle dans les forêts sub-alpines (Haut-Verdon, Alpes du Sud, France). Géographie physique et Quaternaire, 52 : 227-236.
- Bert, G.D., 1993. Impact of ecological factors, climatic stresses, and pollution on growth and health of Silver fir (Abies alba Mill.) in the Jura Mountains — an ecological and dendrochronological study. Acta Oecologica 14 : 229-246.
- Böhm, R., Auer, I., Auer, M., Maugeri, M., Nanni, T. et Schöner, W., 2001. Regional temperature variability in the European Alps : 1760-1998 from homogenized instrumental time series. International Journal of Climatology, 21 : 1779-1801.
- Breda, N. et Pfeiffer, M., 1999. Bilan hydrique et impact des épisodes de sécheresse sur la croissance radiale des chênes. Dépérissement forestier en vallée du Rhin. Rapport ONF-INRA, 60 p.
- Breda, N., Granier, A. et Aussenac, G., 2000. Évolutions possibles des contraintes climatiques et conséquences pour la croissance des arbres. Revue Forestière Française, 52 : 73-90.
- Briffa, K.R., 1992. Increasing productivity of natural growth conifers in Europe over the last century, p. 64-71. In LUNDQUA Report, vol. 34, Proceedings of the International Dendrochronological Symposium, held in Ystad, south Sweden, 3-9 September 1990 (T.S. Bartholin, B.E. Berglund, D. Eckstein and F.H. Schweingruber, eds.), Department of Quaternary Geology, Lund University, Sweden.
- Briffa, K.R., Jones, P.D., Bartholin, T.S., Eckstein, D., Schweingruber, F.H., Karlen, W., Zetterberg, P. et Eronen, M., 1992. Fennoscandian summer from A.D. 500 : temperature changes on short and long timescales. Climate Dynamics, 7 : 111-119.
- Briffa, K.R., Jones, P.D., Schweingruber, F.H., Karlen, W. et Shiyatov, S.G., 1996. Tree-ring variables as proxy-climate indicators : problems with low frequency signals, p. 9-41. In P.D. Jones, R.S. Bradley et J. Jouzel, édit., Climate Variations and Forcing Mechanisms of the Last 2000 Years. Springer-Verlag, Berlin.
- Cook, E.R. et Kairiukstis, L.A., 1990. Methods of Dendrochronology : Applications in the Environmental Sciences. Kluwer Academic Publishers, Dordrecht, 314 p.
- Corona, C., 2007. Évolution biostasique du paysage, géodynamique nivéo-périglaciaire et fluctuations climatiques récentes dans la haute vallée de la Romanche (Alpes du Nord France). Thèse de doctorat, Université de Grenoble, 752 p.
- De Beaulieu, J.-L., Édouard, J.-L., Ponel, P., Rolando, C., Tessier, L., Thinon, M. et Thomas, H., 1990. Timberline and human impacts in the French Alps. The state of the art and research programs. PACT, 31 : 63-80.
- Desplanque, C., 1997. Dendroécologie comparée du sapin et de l’épicéa dans les Alpes internes franco-italiennes. Rôle des facteurs climatiques et anthropiques sur leur répartition. Thèse de doctorat, Université de Grenoble, 151 p.
- Diaz, H.-F., Beniston, M. et Bradley, R.S., 1997. Climatic Change at High Elevation Sites. Kluwer Academic Publishers, Dordrecht, 530 p.
- Di Castri, F., Hansen, A.J. et Holland, M.M., 1988. A new look at ecotones. Biology International, Special issue 17, 163 p.
- Didier, L. et Brun, J.-J., 1998. Limite supraforestière et changements environnementaux : pour une approche pluriscalaire et spatialisée des écosystèmes d’altitude. Géographie physique et Quaternaire, 52 : 245-253.
- DIREN Rhône-Alpes, 2004. Inventaire des zones naturelles d’intérêt écologique, versant montagneux de la Courbe. Direction Régionale de l’Environnement Rhône-Alpes, Lyon, no. 38220004, 3 p.
- Efron, B., 1979. Bootstrap methods : another look at the jackknife. Annals of Statistics, 7 : 1-26.
- Erlandsson, S., 1936. Dendro-chronological studies. Thèse de doctorat, University of Uppsala.
- Francou, B., 1988. L’éboulisation en haute montagne. Thèse de doctorat, Université de Caen, 2 tomes, 696 p.
- Fritts, H.C. et Swetnam, T.W., 1986. Dendroecology : A Tool for Evaluating Variations in Past and Present Forest Environments. Hunton and Williams Printers, Washington D.C., 61 p.
- Grace, J., 1987. Climatic tolerance and the distribution of plants. New Phytologist, Supplement 106 : 113-130.
- Groupe d’Experts Intergouvernemental sur l’Évolution du Climat, 2001. Bilan 2001 des changements climatiques : les éléments scientifiques. Rapport du Groupe de travail I du GIEC, Genève, 205 p.
- Graces, J. et Norton, D.A., 1990. Climate and growth of Pinus sylvestris at its upper altitudinal limit in Scotland : evidence from tree growth-rings. Journal of Ecology, 78 : 601-610.
- Granier, A., Badeau, V. et Bréda, N., 1995. Modélisation du bilan hydrique des peuplements forestiers. Revue Forestière Française, 47 : 59-68.
- Graumlich, L.J., Brubaker, L.B. et Grier, C.C., 1989. Long-term trends in forest net primary productivity : Cascade Mountains, Washington. Ecology, 70 : 405-410.
- Graybill, D.A., 1987. A network of high elevation conifers in the western U.S. for detection of tree-ring growth response to increasing atmospheric carbon dioxide. In Proceedings International Symposium on Ecological Aspects of Tree-Ring Analysis, August 17th, 1986 : 463-474.
- Gregg, B.M., Dougherty, P.M. et Hennessey, T.C., 1988. Growth and wood quality of young loblolly pine trees in relation to stand density and climatic factors. Canadian Journal of Forest Research, 18 : 851-858.
- Grudd, H., Briffa, K.R., Karlén, W., Bartholin, T.S., Jones, P.D. et Kromer, B., 2002. A 7400-year tree-ring chronology for Northern Sweden. The Holocene 12 : 657-665.
- Guiot, J., 1991. The bootstrapped response function. Tree-Ring Bulletin, 51 : 39-41.
- Guisan, A., Holten, J.I., Spichiger, R. et Tessier, L. (édit.), 1995. Potential ecological impacts of climate change in the Alps and Fennoscandian mountains. Conservatoires et jardins botaniques de la ville de Genève, Publication hors série 8, 194 p.
- Hansen, A.J., Risser, P.G. et Di Castri, F., 1992. Epilogue : Biodiversity and ecological flows across ecotones, p. 423-438. In A.J. Hansen et F. Di Castri, édit., Landscape Boundaries : Consequences for Biotic Diversity and Ecological Flows. Springer-Verlag, Berlin, 452 p.
- Hanson, P.J., Todd, D.E. et Amthor, J.S., 2001. A six-year study of sampling and large-tree growth and mortality responses to natural and induced variability in precipitation and throughfall. Tree Physiology, 2 : 345-358.
- Hargreaves, G.H. et Samani, Z.A., 1982. Estimating potential evapotranspiration. Journal of the Irrigation and Drainage Division of the American Society of Civil Engineers, 108 : 223-230.
- Heikkinen, O., 1984. Forest expansion in the subalpine zone during the past hundred years, Mount Baker, Washington, USA. Erdkunde, 38 : 194-202.
- Hétu, B., 1990. Évolution récente d’un talus d’éboulis en milieu forestier, Gaspésie, Québec. Géographie physique et Quaternaire, 44 : 199-215.
- Holmes, R.L., 1983. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 43 : 47-50.
- Holten, J.I. et Carey, P.D., 1992. Responses of climate change on natural terrestrial ecosystems in Norway. NINA forskningsraport 29, 59 p.
- Holtmeier, F.K., 1973. Geoecological aspects of timberlines in northern and central Europe. Arctic and Alpine Research, 5 : 45-54.
- Hustich, I., 1983. Tree-line and tree growth studies during 50 years : some subjective observations. Collection Nordicana, 47 : 181-188.
- Jeanneret, F., Brügger, R. et Vassella, A., 1997. La phénologie forestière : les variations climatiques et l’état des arbres. Paysages glaciaire, karstique et forestier. Société neuchâteloise de géographie, 41 : 55.
- Jenkins, M.A. et Pallardy, S.G., 1995. The influence of drought on red oak group species growth and mortality in the Missouri Ozarks. Canadian Journal of Forest Research, 25 : 1119-1127.
- Jobaggy, E.G. et Jackson, R.B., 2000. Global controls of forest line elevation in the northern and southern hemispheres. Global Ecology and Biogeography, 9 : 253-268.
- Kolischuk, V.G. et Berko, J.M., 1967. Growth of Pinus spp. and fluctuations of climate in the Ukrainian Carpathians. Ukrains’kii Botanichnii Zhurnal, 24 : 39-47.
- Körner, C., 1994. Impact of atmospheric changes on high mountain vegetation, p. 155-166. In M. Beniston, édit., Mountain Environments in Changing Climates. Routledge Publishing Company, London.
- Kräuchi, N., 1993. Potential impacts of a climate change on forest ecosystems. European Journal of Forest Pathology, 23 : 28-50.
- Kräuchi, N., Brang, P. et Schönenberger W., 2000. Forest of mountainous regions : gaps in knowledge and research needs. Forest Ecology and Management, 132 : 73-82.
- Kullman, L., 1993. Holocene thermal trend inferred from tree-limit history in the Scandes mountains. Global Ecology and Biogeography Letters, 2 : 181-188.
- Kullman, L., 1997. Tree-limit stress and disturbance. A 25 year survey of geoecological change in the Scandes Mountains of Sweden. Geografiska Annaler, 79A : 139-165.
- Lafortune, M., Filion, L. et Hétu, B., 1997. Dynamique d’un front forestier sur un talus d’éboulis actif en climat tempéré froid (Gaspésie, Québec). Géographie physique et Quaternaire, 51 : 67-80.
- LaMarche, V.C., Graybill, D.A., Fritts, H.C. et Rose, M.R., 1984. Increasing atmospheric carbon dioxide : tree ring evidence for growth enhancement in natural vegetation. Science, 225 : 1019-1021.
- Lavoie, C. et Payette, S., 1994. Recent fluctuations of the lichen-spruce forest limit in subarctic Québec. Journal of Ecology, 82 : 725-734.
- Lebourgeois, F. et Becker, M., 1996. Dendroécologie du pin laricio de Corse dans l’ouest de la France. Évolution du potentiel de croissance au cours des dernières décennies. Annales des Sciences forestières, 53 : 931-94.
- Lebourgeois, F., 2000. Climatic signals in corsican pine rings. Annals of Forest Science, 57 : 155-164.
- Liang, E., Shao, X., Kong, Z. et Lin, J., 2003. The extreme drought in the 1920s and its effect on tree growth deduced from tree ring analysis : a case study in North China. Annals of Forest Science, 60 : 145-152.
- MAAPR, 2003. Sécheresse et canicule. Les conséquences sur les peuplements forestiers. Ministère de l’Agriculture, de l’Alimentation, de la Pêche et de la Ruralité, Information santé des forêts, Publications du département de la santé des forêts, 4 p.
- MacDonald, G.M., Szeicz, J.M., Claricoates, J. et Dale, K.A., 1998. Response of the central Canadian treeline to recent climatic changes. Annals of the Association of American Geographers, 88 : 183-208.
- Mäkinen, H., Nöjd, P. et Mielikäinen, K., 2001. Climatic signal in annual growth variation in damaged and healthy stands of Norway spruce (Picea abies (L.) Karst.) in southern Finland. Trees, 15 : 177-185.
- Mast, J.N., Veblen, T.T. et Thomas, T., 1994. A dendrochronological method of studying tree mortality patterns. Physical Geography, 15 : 529-542.
- Moen, J., Aune, K., Edenius, L. et Angerbjörn, A., 2004. Potential effects of climate change on treeline position in the Swedish mountains. Ecology and Society, 9 : 16.
- Nicolussi, K., Bortenschlager, S. et Körner, C., 1995. A possibly CO2-related increase in tree-ring width in subalpine Pinus cembra of the central alps. Trees, 9 : 181-189.
- Ogle, K., Whitham, T.G. et Cobb, N.S., 2000. Tree-ring variation in pinyon predicts likelihood of death following severe drought. Ecology, 81 : 3237-3243.
- Oldeman, R.A.A., 1990. Forests : Elements of Silvology. Springer-Verlag, Heidelberg, 624 p.
- Ozenda, P., 1985. La végétation de la chaîne alpine dans l’espace montagnard européen. Masson, Paris, 331 p.
- Payette, S., Fortin, M.-J. et Morneau, C., 1996. The recent sugar maple decline in southern Quebec : probable causes deduced from tree rings. Canadian Journal of Forest Research, 26 : 1069-1078.
- Peterson, D.L., Arbaugh, M.J., Robinson, L.J. et Derderian, B.R., 1990. Growth trends of whitebark pine and lodgepole pine in a subalpine Sierra Nevada forest, California, USA. Arctic and Alpine Research, 22 : 233-243.
- Petitcolas, V., 1998. Dendroécologie comparée de l’Épicéa, du Mélèze, du Pin cembro et du Pin à crochets en limite supérieure de la forêt dans les Alpes françaises : influence de la variabilité macro-écologique. Thèse de doctorat, Université de Grenoble, 186 p.
- Rathgeber, C., Guiot, J., Roche, P. et Tessier, L., 1999. Augmentation de productivité du chêne pubescent en région méditerranéenne française. Annales des Sciences Forestières, 56 : 211-219.
- Rathgeber, C., 2002. Impact des changements climatiques et de l’augmentation du taux de CO2 atmosphérique sur la productivité des écosystèmes forestiers : exemple du pin d’Alep (Pinus halepensis Mill.) en Provence calcaire (France). Thèse de doctorat, Université Aix-Marseille, 312 p.
- Richter, K., Eckstein, D. et Holmes, R.L., 1991. The dendrochronological signal of pine trees (Pinus ssp.) in Spain. Tree Ring Bulletin, 51 : 1-13.
- Rolland, C., 1993. Fonctionnement hydrique et croissance du sapin (Abies alba Mill.) dans les Alpes françaises. Thèse de doctorat, Université de Grenoble, 180 p.
- Rolland, C. et Schueller, J., 1996. Dendroclimatologie du Pin à crochets (Pinus uncinata Mill. ex Mirb.) dans le Briançonnais et le Queyras en fonction des conditions stationnelles. Schweizerische Zeitschrift für Forstwesen, 147 : 351-363.
- Rolland, C., 1996. Croissance du Pin à crochets (Pinus uncinata Mill. ex Mirb.) dans la réserve naturelle des Hauts-Plateaux du Vercors. Revue forestière française, 48 : 144-152.
- Solomon, A.M. et Kirilenko, A.P. 1997. Climate change and terrestrial biomass : What if trees do not migrate ! Global Ecology and Biogeography Letters, 6 : 139-148.
- Spiecker, H., Mielikaïnen, K., Köhl, M. et Skovsgaard, J., 1996. Growth Trends in European Forests. Springer-Verlag, Heidelberg, 372 p.
- Taylor, A.H., 1995. Forest expansion and climate change in the Mountain Hemlock (Tsuga mertensiana) zone, Lassen volcanic National Park, California, USA. Arctic and Alpine Research, 27 : 207-216.
- Tessier, L., 1989. Spatio-temporal analysis of climate-tree ring relationships. New Phytologist, 111 : 517-529.
- Theurillat, J.P., 1995. Climate Change and the Alpine flora : some perspectives, p. 121-128. In A. Guisan, J.L. Holten, R. Spichiger et L. Tessier, édit., Potential Ecological Impacts of Climate Change in the Alps and Fennoscandian Mountains. Conservatoires et jardins botaniques de la Ville de Genève, Publication hors série 8.
- Thevenon, O., 1999. Les changements climatiques récents en montagne : l’exemple du massif de l’Oisans. Mémoire de maîtrise, Université de Grenoble, 117 p.
- Vennetier, M. et Hervé, J.-C., 1999. Short and long term evolution of Pinus halepensis (Mill.) height growth in Provence (France) and its consequences for timber production. In Causes and Consequences of Accelerating Tree Growth in Europe, International Seminar held in Nancy, May 14-16 1998. European Forest Institute Proceedings 27 : 263-265.
- Villalba, R. et Veblen, T.T., 1997. Spatial and temporal variation in Austocedrus growth along the forest-steppe ecotone in northern Patagonia. Canadian Journal of Forest Research, 27 : 580-597.
 10.7202/004767ar
10.7202/004767arListe des figures
Figure 1
Localisation du site d’étude et stratégie d’échantillonnage. Les coordonnées géographiques sont exprimées en latitude/longitude (WGS 84).
Localisation of the study area and the sampling strategy. Map coordinates are in latitude/longitude (WGS 84).
A
1913
B
1935
C
1952
D
1967
E
1981
F
2001
Répartition du pin noir d’Autriche sur le talus d’éboulis de la Courbe entre 1913 et 2001.
Distribution of Austrian black pine on the scree slope between 1913 and 2001.
Figure 3
Surfaces occupées par les différentes formations végétales sur le talus d’éboulis de la Courbe entre 1913 et 2001.
Occupied area by vegetation types on the scree slope between 1913 and 2001.
Figure 4
Nombres d’individus établis par décennie au sein des 12 placettes étudiées.
Decennial tree numbers on the 12 sample plots.
Figure 5
Évolution de l’indice moyen de croissance radiale (lc %) au cours de la période 1896-2003.
Evolution of the mean radial growth rate (lc %) over the period 1986-2003.
A
B
C
Fonctions de réponse calculées entre la chronologie indicée de largeurs de cernes du Freney et les données climatiques mensuelles (octobre (n–1) à septembre (n)) de la station météorologique de Besse (1959-2003). (A) Totaux pluviométriques et températures mensuelles moyennes. (B) Totaux pluviométriques, températures mensuelles maximales (Tx) et minimales (Tn). (C) Déficit hydrique (P–ETP) obtenu à partir de la formule de Hargreaves. Les valeurs présentées correspondent à un ratio entre les coefficients de régression partiels et leur écart-type, les lignes horizontales indiquent une significativité à des seuils de 90 %, 95 %, 99 % et 99,9 %.
Response function calculated from the tree-ring index at Le Freney and monthly climatic data (October (n–1) to September (n) at Besse Weather Station (1959-2003). (A) Grey bars refer to precipitation-mean monthly temperature matrix. (B) Black bars refer to the precipitation-mean maximum temperature (Tx) matrix, grey bars to the precipitation-mean minimum temperature (Tn) matrix. (C) Grey bars refer to evapotranspiration (P–ETP) as predicted by Hargreaves Evapotranspiration Model. The values shown are the ratio between partial regression coefficient and its standard deviation horizontal lines represent significance thresholds.
A
Trait plein en gris : évolution des écarts à la moyenne de la saison estivale calculée pour les Alpes occidentales sur la période 1880-1998 (Böhm et al., 2001) ; trait gras en gris : moyenne mobile calculée sur 10 années.
Thin grey curve : mean summer temperature anomalies shown as deviation in °C from 1880-1998 for the Western Alps (Böhm et al., 2001) ; thick grey curve : 10-year running mean. Thin black curve : mean summer temperature anomalies shown as deviation in °C from 1959-2003 at Besse Weather Station ; thick black curve : 10-year running mean.
B et C
Trait plein en noir : écarts à la moyenne (1930-2003) des totaux pluviométriques mensuels ; trait gras en noir : moyenne mobile calculée sur 10 années.
Thin black curve : precipitation anomalies shown as deviation in mm from the 1930-2003 mean ; thick black curve : 10-year running mean.
Évolution des écarts à la moyenne des températures moyennes estivales (juin, juillet et août) (A) et des totaux pluviométriques mensuels de mai (B) et de juillet (C) à la station météorologique de Besse.
Mean summer temperature anomalies (June, July, August) (A) and mean monthly precipitation anomalies in May (B) and July (C) at Besse Weather Station.
Liste des tableaux
Tableau I
Caractéristiques de croissance des 175 pins noirs
N total : nombre de pins par placettes.
N échantillonné : nombre de pins échantillonnés par placettes.
SM : sensibilité moyenne, mesure de la variabilité inter-annuelle.
AC : coefficient d’autocorrélation de premier ordre.
Les deux paramètres, SM et AC, ont été calculés sur les séries brutes de largeur de cernes.
Tableau II
Coefficients de détermination (r2) calculés entre les séries saisonnières de Besse en Oisans et des Alpes occidentales pour la période 1964-1998
Source : Météo France.
Source : Böhm et al., 2001.
Tableau III
Coefficients de régression linéaire (r) et coefficients de détermination (r2) calculés sur les températures mensuelles minimales (Tn), moyennes (Tm), maximales (Tx), sur les amplitudes thermiques (At), le déficit hydrique (P–ETP) (période 1959-2003) et les totaux pluviométriques (Pn) (période 1930-2003) de la station météorologique de Besse en Oisans