Résumés
Abstract
The harvesting of narwhals by Baffin Island Inuit represents an important relationship in terms of the continuous utilization of an indigenous marine resource. However, research on Inuit hunting with respect to narwhals has been mainly confined to harvest counts despite the major role narwhals play in the local northern Baffin subsistence system. The present research examines Pond Inlet Inuit foraging behaviour for narwhals in the spring floe-edge environment. While sea ice is one of the most dominant features of the arctic marine environment for much of any year, it is in the spring that the dynamism of its physical and biological characteristics is most notable. This was especially evident at the fast ice-open water interface, or floe-edge, where rapid physical change in the condition of the ice is frequent and summer migratory marine mammals and birds are present in large numbers. In this paper, analysis of 14 observed hunts indicates that Inuit utilization of the spring floe edge for narwhal hunting, in contrast to most other hunt types, follows a sit-and-wait mode of foraging. The study also explicates aspects of Pond Inlet hunters traditional ecological knowledge necessary to travel and conduct harvesting operations successfully in this complex environment.
Résumé
La chasse au narval par les Inuit de la Terre de Baffin représente une relation importante en terme d’une utilisation continue d’une ressource marine indigène. Cependant, la recherche portant sur la chasse au narval par les Inuit a surtout été limitée aux dénombrements des récoltes, malgré le rôle majeur que les narvals jouent dans le système local de subsistance. La présente recherche explore le comportement alimentaire des Inuit face aux narvals dans l’environnement printanier de la lisière des banquises (floe-edge). Tandis que la glace marine est l’aspect dominant de l’environnement arctique marin pendant la majeure partie de l’année, c’est au printemps que le dynamisme de ses caractéristiques physiques et biologiques est le plus notable. Ceci est particulièrement évident à l’interface rapide de la glace et des eaux dégagées, nommément la lisière des banquises, où le changement physique rapide dans la condition de la glace est fréquent et où les mammifères marins et les oiseaux migrateurs estivaux sont présents en grand nombre. Dans cet article, une analyse de 14 voyages observés indique que l’utilisation par les Inuit de la lisière des banquises au printemps pour chasser le narval, comparativement aux autres types de chasse, suit un mode de chasse à l’affût (sit-and-wait). Cette étude expose les aspects du savoir écologique traditionnel appartenant aux Inuit de Pond Inlet, nécessaire pour voyager et conduire les opérations de chasse avec succès dans cet environnement complexe.
Corps de l’article
Introduction
Research on Inuit hunting of narwhals (Monodon monoceros; Inuktitut: qilalugaq) in the floe-edge environment has been limited both in number of studies and depth of analysis (e.g., Bissett 1968; Degerbøl and Freuchen 1935; Mary-Rousselière 1984). For example, most data on Inuit-narwhal interaction consists only of a few numbers in harvest tables (see Baffin Regional Inuit Association 1982, 1983, 1984, 1985; Donaldson 1988). Clearly, there is a general paucity of analytical literature on Inuit cetacean hunting, despite the acknowledged ethnographic and current importance of narwhal to Inuit (notable exceptions are Dahl 1990, 2000 and Smith 1991).
Nevertheless, the actual use of narwhals by Inuit has not gone completely unremarked. Both anthropological and biological literatures have explored the culture of the hunt (Mathiassen 1928), the animal’s dietary importance (Bissett 1968; Treude 1977) and its socioeconomic value (Land 1977; Mary-Rousselière 1984; Matthiasson 1992; Reeves 1992). With reference to the actual hunt, Finley and others (Finley et al. 1980; Finley and Miller 1982; Roberge et al. 1990; Weaver and Walker 1988) have investigated harvesting technologies while Remnant and Thomas (1992), Thomsen (1993) and Stewart et al. (1995) have investigated features of the traditional ecological knowledge of modern narwhal hunting. As much as these works contribute to our understanding of the Inuit and narwhal relationship, none provide a comprehensive analysis of contemporary narwhal hunting behaviour.
The main objective of this paper is to redress this gap by employing a behavioural ecology approach to study the Inuit floe-edge narwhal hunt. An analysis of Inuit hunting strategies is critical to understand the organization and pattern of Inuit hunting behaviour with respect to the prey species and the environment. In the present study, we specifically focus on one of the most important components of the Inuit narwhal hunt during the spring floe-edge season: the foraging or search phase (see Table 1). We believe this stage to be crucial for the overall foraging cycle as it is time-consuming, precarious and is most influential in the successful capture of a narwhal.
Table 1
Inuit floe-edge narwhal foraging cycle.

A second objective is to examine whether the foraging mode currently used at the floe-edge to hunt narwhal in the spring is specific to this species and environment or represents a widespread strategy. Hunting processes for other species such as caribou hunting, winter breathing hole sealing, and spring basking seal hunting will be compared to narwhal hunting to present the variation in foraging mode. Winter and spring sealing represent two harvest activities that might be considered to “compete” with whaling at this season. Finally, it is important to mention that no comparison will be made here between floe-edge and open-water narwhal modes of foraging as this is the subject of a future paper (Lee, in preparation).
Next, we review the relationship between Pond Inlet Inuit and narwhals to illustrate the contemporary pattern and importance of Inuit narwhal harvesting. We will also review the human behavioural ecology framework as it will be the basis of our present analysis of Inuit narwhal hunting behaviour.
Pond Inlet, narwhals and Inuit
Narwhals and their products have widespread cultural importance for Inuit. The skin (maktaaq), with some attached fat (uqsuq), is the most important food item furnished by narwhal hunting and is often considered a delicacy (Reeves 1992; personal observations). Historically, the narwhal also provided raw materials for domestic use, such as dried sinews for waterproof seams. However, it was the species’ ivory tusk that was the principal object of late 19th and early 20th centuries trade between Inuit and Europeans (Bernier 1909, 1911).
The narwhal is found today in the open water environment of the Eastern Canadian Arctic from early May to late September. The narwhal mainly frequents northwestern Baffin Bay, Lancaster and Jones Sounds, western Foxe Basin, and eastern Barrow Strait (Kingsley et al. 1994; Koski and Davis 1994; Reeves 1992; Richard et al. 1994; Silverman 1979; Smith et al. 1985, Strong 1988). Pond Inlet (Mittimatalik) is the northernmost community on Baffin Island located at 72°41”N, 77°58”W, and is adjacent to the Baffin Bay-Pond Inlet floe-edge (Figure 1). This location was selected for the present study because of its large annual narwhal quota of 100 animals. The area of Inuit-narwhal interaction of direct interest here is the Pond Inlet spring floe-edge (Figure 2). This floe-edge represents a primary habitat that has long provided Mittimatalingmiut (Pond Inlet Inuit) with access to northward migrating narwhals (Kingsley et al. 1994; Koski and Davis 1994; Reeves 1992; Richard et al. 1994; Silverman 1979).
Although the archaeological record is intermittent, it seems that narwhals have been hunted in this region for some two millennia (Savelle 1994). More specifically, it is clear that Mittimatalingmiut have possessed the technology, expertise and ecological knowledge to hunt this species intensively since at least the early 19th century (Mary-Rousselière 1984). Their unique skill developed as a result of their geographic situation (Figure 2) that provides exceptional opportunity to intercept spring-migrating, as well as summering narwhals (Kingsley et al. 1994; Koski and Davis 1994; Richard et al. 1994; Silverman 1979; Smith et al. 1985). More recently, the introduction of modern hunting and transportation technologies (notably high-powered rifles, motor-equipped canoes and snowmobiles) has allowed Mittimatalingmiut to continue to exploit narwhals despite centralization from their former dispersed pattern of settlement to residence in modern Pond Inlet (Brody 1975; Damas 2002; Matthiasson 1967).
Mittimatalingmiut commence narwhal hunting in spring (approximately mid-May) and continue hunting from the floe-edge until break-up (approximately mid-June). In early May, the first narwhals begin to arrive at the eastern floe-edge (Hay 1984; Silverman 1979), and a few hunters, considered by their peers to be particularly adept at floe-edge hunting, begin to seek whales at this time. These hunters often stage their forays onto the sea ice from Button Point (Sannirut) on southwest Bylot Island where there are two community-supported cabins (Figure 2).
By mid-May, the number of narwhal at the eastern floe-edge substantially increases (Hay 1984; Silverman 1979). At this time, the movement pattern of hunters undergoes a shift. While a few individuals continue to camp at Button Point, most hunters now stop there only to rest briefly and to acquire information on floe-edge and weather conditions, narwhal sightings, and camp locations along the floe-edge from other Inuit camped and moving through Button Point. This pattern of proceeding to Button Point before actually beginning operations along the floe-edge continues into early June; by then, whales are present along the entire extent of the floe-edge, except when pack ice jams along the floe.
Figure 1
Location of Pond Inlet (Mittimatalik) and study area.

Human behavioural ecology: mode of foraging
Our study of Inuit hunting behaviour adds to the present literature of human behavioural ecology (HBE; see Winterhalder and Smith 2000) through our analysis of mode of foraging. Historically, since the mid 1970s, HBE studies have applied optimal foraging models to study the subsistence practices of indigenous peoples (e.g., Hawkes et al. 1991; Hawkes and O’Connell 1982, 1985, 1992; Hames and Vickers 1982; Hill 1988; Sih and Milton 1985; Smith 1991; Winterhalder 1977, 1983). Optimal foraging models assume that foragers attempt to maximize their immediate energy gains over time (Stephens and Krebs 1986). These are mathematical models where ecological parameters are entered in formulated equations to make testable predictions of foraging behaviour such as diet composition, group size and patch selection among different groups in different habitats. As robust as the optimal foraging approach is in its predictions, it has received criticism for its lack of descriptive utility and its inability to incorporate the rich qualitative data found in ethnographic studies (e.g., Smith 1983).
We address such criticisms by departing from an optimal foraging approach as applied in HBE studies; instead, we focus on the role of foraging mode (see Holling 1959; Kramer 2001). Whereas optimal foraging models attempt to condense and generalize foraging behaviour in order to address universal questions such as what individuals eat, where they look for food, and how long they search for their food, our proposed focus on foraging mode offers a more descriptive and qualitative analysis of specific search behaviour of prey. We hope this alternative theoretical perspective will place Inuit hunting behaviour in a richer and more realistic context. Our application of foraging mode to a human population is the result of our extensive observations which suggest a strong correspondence between the foraging mode of Inuit and other animals. The application of foraging mode is useful because it describes the actual hunting behaviour Inuit display in their habitats. It also elucidates some of the observable ecological factors that influence this behaviour.
In the ethological literature on animal foraging mode, it was observed that some predators attack their prey from ambush, whereas others usually hunt while on the move. Pianka (1966) termed these modes of foraging respectively "sit-and-wait" and "widely-foraging." In the sit-and-wait foraging mode, a forager remains stationary for long periods of time, waiting for a prey item to come within short pursuit or striking distance. In the widely-foraging mode, a forager spends much of its time actively searching for prey (Huey and Pianka 1981). Although this dichotomy can be somewhat artificial, numerous animal and human groups (see Binford 1980) seem to fall into one or the other category.
Research methodology
The primary data presented here were collected from 1996 through 1998, utilizing a participant-observation methodology supplemented by directed interviews. In total, almost 11 months were spent accompanying and observing Mittimatalingmiut narwhal activities in the floe-edge and open water environments. The floe-edge hunting component of the research comprised 14 trips. The accompanied hunters were varied with respect to age and individual experience in narwhal hunting (Table 2).
This participatory approach was ideal for recording hunter movements, stops and campsites, while also allowing the observation of contiguous or passing narwhal hunters. Additionally, all search, pursuit, and handling activities were timed and amounts of fuel, ammunition and other expendables used were all recorded. Finally, travel routes, encountered hazards, camping/resting sites, and narwhal sightings/firing events were recorded.
Table 2
Social and demographic characteristics of hunting groups.

Floe-edge mode of foraging
Active foraging mode
At 11:30, we reach the floe-edge almost directly across from Sannirut […] D. is scanning the area as we continue to travel south.
Lee, Field Notes, 14/06/96
During the course of the research, an active or widely foraging mode was characterized as a relatively continuous movement where hunters travelled parallel to the floe-edge in one direction while simultaneously scanning the open water for narwhal. Movements between camps were also considered as part of the active foraging mode because hunters continued to search for narwhal while traveling. Arguably, inter-camp movements could be considered simply as travel time; however, the scanning for prey supported inclusion as active foraging search.
Active foraging by snowmobile began as soon as hunters arrived at the floe-edge (Table 3). Upon arrival at the floe-edge, initial scans for narwhals were made and a travel direction was then chosen. This phase was particularly important because critical environmental information for travel safety and campsite selection was collected and given a priority greater than, or at least equal to, the search for narwhal. For example, the structural stability of the ice edge, sea conditions, weather patterns and the movements of drifting ice were all collected and synthesized. Equally important to the physical characteristics, this phase also provided the hunters with the number and distribution of the status of campsites located within the regional vicinity.
Snowmobile foraging was frequently punctuated by scan-stops. Scan-stops were brief pauses that generally lasted from 10 to 30 minutes and almost always occurred if a narwhal was sighted in order to evaluate the narwhal’s position and/or to hunt the narwhal with sit-and-wait foraging tactics. Such tactics should not be confused with the sit-and-wait foraging mode. Sit-and-wait foraging tactics refer to manoeuvring and positioning to hunt narwhal over a short term. These tactics were always employed because narwhal would readily dive and be very challenging or impossible to track actively in the pack ice environment. Scan-stops were also executed to evaluate the stability of an ice platform with an ice probe before travelling upon it.
Hunters who carried out extensive active searching by snowmobile incurred two kinds of costs. The first was monetary as a snow machine consumed about CDN $25.00 in gasoline and oil for roughly every three to four hours of operation (average price as calculated at the time of study, 1996-1998). Perhaps even more important was that the noise associated with snowmobile use was said by Inuit to affect how closely narwhal would approach the floe-edge and ultimately affect hunting opportunities.
Finally, a negligible amount of active foraging along the floe-edge based from the campsite was also done on foot. Walking provided the advantage of almost no disturbance. It did, however, have disadvantages. First, only a limited extent of floe-edge could be covered. Second, if a narwhal was shot, the chance of a successful retrieval without a boat was unlikely. Third, venturing any significant distance from camp meant risking exposure to possible deteriorating ice or rapid weather change. Last, it was difficult to be perfectly quiet while walking; some Inuit stated that the sounds of a hunter’s footsteps are interpreted by nearby narwhal as the sound of a polar bear shuffling on the sea ice.
Table 3
Number and duration of active foraging events (AFE) based from campsite and total duration of all active foraging (all duration in minutes).

*Includes active foraging before first camp, AFE, and foraging between campsites.
Sit-and-wait foraging mode
At 13:40 hours, we sighted three narwhals moving south from our camp location. They are approximately 200 metres away from our location. They breathe for 8-10 minutes and then one takes a very deep breath and dives. The others follow almost immediately afterwards. They were very large in size with white coloration.
Lee, Field Notes, 29/06/96
The sit-and-wait foraging mode was observed when hunters deployed a campsite where they searched for narwhal by remaining stationary for an extended period of time. The sit-and-wait foraging mode comprised all of the time spent by hunters who were stationary in their campsites. Although this included additional activities such as cooking, cleaning, resting, preparing and repairing equipment, hunters were always vigilant for narwhal while remaining stationary in their campsite. Overall, the average length of stay at the floe-edge for the 14 hunts observed over these three years was two and a half days (SD = 1.74; Range = 0.2 - 7.4 days). Given this average length of stay at the floe-edge, each hunt established at least one camp (Table 4). In fact, the average number of campsites for all trips was two (see Table 4) (SD = 0.97).
Figure 2
Spring floe edge Inuit use area and narwhal migration routes.

Figure 3
Sit-and-wait (cross-hatched bars) and active (white bars) foraging behavior distributions for each trip relative to total foraging time is presented. The sit-and-wait mode is clearly the most dominant search behavior in all participant observed floe-edge narwhal trips.

Table 4
Number of camps established and camp residence duration, and total duration of all sit-and-wait foraging (all duration in minutes).

*Includes camp residence duration but not active foraging activity based from campsite (see AFE in Table 3).
Camp routine was as such that at least one person was on alert and scanning for whales at all times in multi-occupant camps. This explains why the average number of groups per camp in all three years was two even though seven[1] of the 14 hunts that were accompanied through their entirety were by single hunters (see Table 2). Solitary camping was actually relatively rare as visits from passing hunters were frequent. This was usually due to the high camp density location chosen by the solitary camper. Although narwhal was the primary objective while waiting in camp, Inuit were also continuously scanning the local environment for deteriorating ice conditions, weather conditions and polar bears.
Comparison of foraging mode
The time spent engaged in each foraging mode reveals the dominant search behaviour of Inuit narwhal hunting at the floe-edge. Of the overall 840 hours spent at the floe-edge for the 14 trips, an average of 94% of hunters’ time was spent in the sit-and-wait foraging mode (SD = 3.62; Figure 3; Table 5).
Table 5
Duration of sit and wait vs. active foraging activity (in minutes).
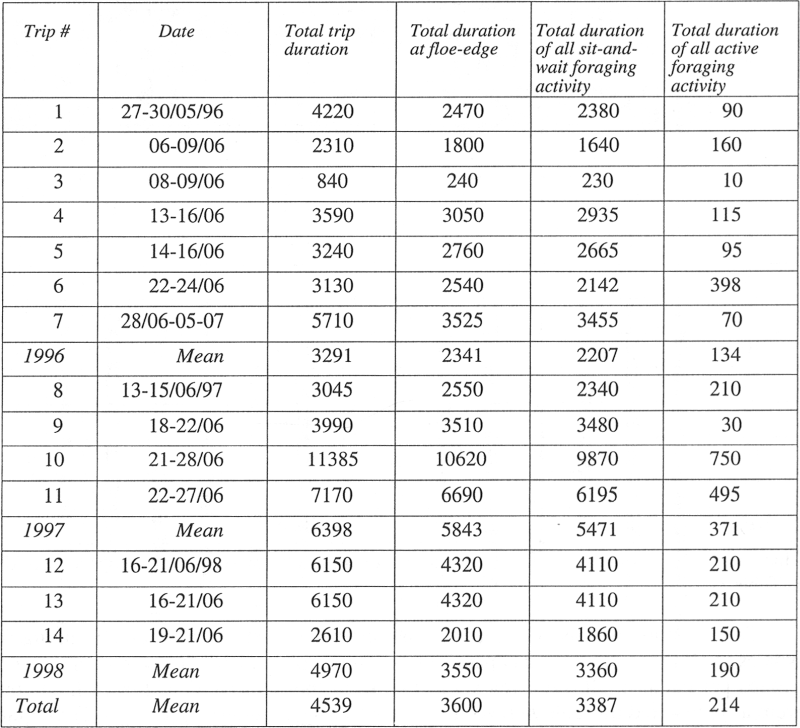
A 3 (years) x 2 (foraging mode) mixed-model ANOVA was conducted, where years was set as the between-subject factor and foraging mode was set as the within-subject factor. The analysis revealed a significant main effect for foraging mode, F (1, 11) = 42.95, p < 0.01; that is, hunters spent significantly more time in sit-and-wait foraging than in active foraging. As expected, the interaction between years and foraging mode was not significant, F (2, 11) = 3.42, p = NS; that is, the pattern of relatively higher time spent in sit-and-wait foraging was consistent over the years.
Despite the large number of trips reviewed here, and the large amount of time spent in both modes of foraging, the rate of successful narwhal capture was low. Overall, only one narwhal was captured. Generally, even the most assiduous hunter was often prevented from having success because of numerous environmental barriers, such as high waves or fog along the floe edge that obscured narwhal target, to broken ice inhibiting any opportunity for retrieval of a kill. Still, there were more actual firing events (capture opportunities) when hunters were engaged in the sit-and-wait foraging mode (Table 6).
Table 6
Floe-edge narwhal sightings and firing events.

Campsite selection
Given the inherent importance of campsites for the sit-and-wait foraging mode, campsite selection bears some discussion. Indeed, when camp residence duration and total trip duration are compared, it indicates that 79% of hunters’ time was spent in camp. This comparison was even more salient when the total time spent at the floe-edge was compared to the camp residence duration: hunters spent 99% of their time near their campsite.
A) Seasonality and geography
Historically, hunters mostly established their campsites near Sannirut during the spring season (Degerbøl and Freuchen 1935; Mary-Rousselière 1984). Presently, the majority of hunters from Pond Inlet actually venture across a larger expanse of the floe-edge. In terms of scheduling and seasonality, more specialized Pond Inlet Inuit concentrate their campsite selection in the southern extent of the floe-edge (see Figure 2), the geographical area where the probability of sighting and intercepting northward-migrating narwhals is highest (Hay 1984; Silverman 1979). By early June, all hunters disperse their campsites along the entire extent of the floe-edge.
Interviews with Mittimatalingmiut elders suggest another reason for why some hunters concentrate on the southern section of the floe-edge in early spring. They stated that when noise disturbance is minimal, narwhals often travel in very close proximity to the floe-edge (as close as 5m). This was the case before mechanized transport (that is before ca.1960) became ubiquitous in the Pond Inlet area. In fact, this a point remarked upon by Degerbøl and Freuchen (1935: 259) in their section on narwhal: “[…] the Eskimos were continually catching those that come near the ice edge, without the others apparently being disturbed in the least by it.” In early spring it is possible, because of the paucity of snowmobile traffic, to take narwhals at much closer range than later in the season. Presently, active hunters generally concurred with the elders’ conclusion that, by the time spring is well advanced, snowmobile and other noise sources make it increasingly difficult to hunt narwhals in close proximity to the floe-edge. Thus, the selection of the southern region of the floe-edge in early spring appears to be an attempt by hunters to select narwhal that exhibit a propensity to travel in close proximity to the floe-edge before excessive noise affects their migratory behaviour.
During the mid stage of the hunting season, distribution of hunters along the entire expanse of the floe-edge hunting area was observed. However, as the season progressed into late June, hunters did not proceed near Sannirut as the ice conditions were observed to be hazardous and elders advised avoidance of this area.
B) Physiognomic features
The physiognomy of the ice front also plays an important role in campsite selection. This is because there are indented patches along the floe-edge, essentially areas where embayments are formed, that offer certain tactical advantages to hunters. Accordingly, it was observed that most hunters establish their campsites near embayments.
Hunters explained that narwhals tend to dive toward embayments, which allows a clear view of, and shot at, the animal’s spine (dive hump). Also, narwhals take a large amount of air into their lungs when about to make a deep dive, which helps keep these animals afloat after a fatal shot. The additional float time associated with this deep dive buoyancy gives hunters critical additional time to launch a boat to effect retrieval. Therefore, hunters target the “dive hump” in order to immobilize an animal and increase the probability of retrieval. Inuit contrast this situation to times when narwhals are swimming or resting at the surface, as at these times a “dive hump” is not visible and, thus, this vital area is less likely to be struck when fired upon.
While historically, narwhals would have been harpooned while travelling along the floe-edge, (i.e. while swimming very close to and parallel to the floe-edge), harpooning was rarely observed during the trips that were accompanied for the present study. Instead, in almost every hunting event that occurred, narwhals were fired upon while making deep dives as they swam toward the floe-edge in one of these embayments. Clearly, while rifles and outboard motor-equipped boats now play a critical technological role in narwhal hunting at the floe-edge, Inuit have also had to adapt their foraging behaviour to compensate for some of the changes in narwhal behaviour precipitated by these tools.
Discussion
First, we summarize our analysis of Inuit foraging mode for narwhal hunts during the floe-edge spring season. Second, we discuss foraging modes for other species with special reference to breathing hole seal hunting and basking seal hunting. Finally, we explore the dynamic effect of the floe-edge environment upon the Inuit foraging mode for narwhals and we discuss the impact of rifle and snowmobile technologies upon foraging mode.
Our observations show that Inuit foraging behaviour for narwhals in the floe-edge environment can clearly be categorized into two major foraging modes: active foraging and sit-and-wait foraging. Our observations also reveal that the sit-and-wait foraging mode was the dominant one utilized to hunt narwhals in the floe-edge environment during the spring hunting season.
The decision to engage in a sit-and-wait strategy is due to several key factors related both to the highly mobile and acoustically sensitive nature of narwhals (Cosens and Dueck 1988; Finley et al. 1990; Ford 1976) and the dynamic ice environment (Parkinson 1999, 2000). For example, the reluctance to engage in active foraging trips by snowmobile seems to indicate the importance of minimizing noise disturbance and maximizing firing opportunities. This is supported by interviews with both elders and active hunters who related their perception that narwhals were extremely acoustically sensitive to noise. Mittimatalingmiut elders also stated that historically (ca. 1900), there were strict rules on human behaviour while at the floe-edge that were utilized in order to minimize noise disturbance. For example, an Inuk elder stated that while hunting narwhals at the floe-edge during spring, any movement among family members not engaged in hunting was not permitted. Furthermore, several elders mentioned that the contemporary snowmobile noise along the floe-edge had drastically changed narwhal migration behaviour along the floe-edge. Even though active trips by foot created much less noise disturbance along the floe-edge, they offered very limited range beyond the camp deployment area.
Notwithstanding, the active foraging mode was shown to play a key role in the initial phase of the hunting trip. In every accompanied trip a preliminary evaluation of the geographical, physical and environmental conditions began as soon as the hunters reached the floe-edge. In these evaluations, hunters first examined the spatial geography and physical status of the floe-edge within viewable range in order to gauge the costs and challenges involved in traveling along the floe-edge and in locating potential campsite areas along the entire floe-edge extent. The foremost concern was whether an area or stretch of floe-edge might rapidly erode or break away. Other factors included whether near-edge travel would require moving over or crossing between separated floes, traversing compacted and uplifted ice ridges, or the possibility of encountering snow conditions that might impede movement. Finally, local weather patterns were considered.
Once a preliminary scan of the physical conditions was done, hunters turned to an assessment of the migration stage of narwhals in the immediate area. The number of narwhal sightings made in these initial scans ultimately influenced a hunter’s decision to travel either north or south along the floe-edge and where camps might be feasible. It should be noted that, along with their own preliminary scans, hunters also frequently benefited from information provided by other hunters already on or returning from the floe-edge through direct contact and radio. Approaching the floe-edge from Button Point (see Figure 2) aided this scanning as Button Point not only provided hunters with shelter if poor or violent weather developed or floe-edge conditions deteriorated, but also allowed an extended view of the floe-edge.
Foraging mode for other species
The hunt for narwhals at the floe-edge represents the most intensive example of the sit-and-wait method employed by Inuit for any species and differs markedly from virtually every other Inuit foraging activity, except for spring and autumn arctic char fishing. For instance, caribou hunting today involves a large component of active search and pursuit. Notably, this is a recent change, as ethnographic studies in the 19th and early 20th century described caribou hunting as a more passive activity in which interception or ambush at water bodies was the dominant hunt form. Other types of Inuit large game hunting, such as polar bear, walrus or large and small whales in certain open-water environments also follow an active foraging mode and pursuit mode in which the active search mode is critical to locating the prey species.
Although, there is an important sit-and-wait foraging component with respect to winter breathing hole sealing, it is still unique from the sit-and-wait mode executed in the floe-edge environment with respect to narwhal. We will discuss both the foraging mode of breathing hole seal hunting and basking seal hunting to elucidate the unique behaviour Inuit currently employ for narwhal in the spring floe-edge season.
A) Breathing hole seal hunting (mauliqpuq)
Mauliqpuq sealing is conducted in virtually every community in Nunavut from the onset of the ocean freezing (for most areas by December) until roughly late May or early June when the snow and ice that cover the seals’ agluit (breathing holes) melt. Thereafter, seals begin to haul out onto the ice surface. Ethnographic accounts of this type of sealing from the last two centuries (Boas 1888; Jenness 1922; Rasmussen 1929, 1931) are notable for their emphasis on the patience of hunters and the extended duration of some hunts, indicating that a hunter might wait up to several hours at an active aglu. Even in the early 1970s, it was recorded that hunters from the village of Arviqtuuq, north of Clyde River on average spent approximately 50 minutes at an aglu (Wenzel 1991). It was not extraordinary for a hunter to wait beyond an hour: the longest wait recorded by Wenzel was 94 minutes.
More recently, Wenzel (1991) has pointed out that the rifle has had a negligible impact in terms of overall efficiency relative to the traditional use of the harpoon. However, this is not to deny that the traditional mode of breathing hole hunting has not undergone modification due to introduced technology. Rather, the search phase of mauliqpuq has been significantly affected by the replacement of the dog team with the snowmobile. Traditionally, once a suitable sealing area was located (in this dogs proved themselves superior to snowmobiles due to their keen sense of smell), there was only a very limited amount of time expended in search and travel between active holes (see Wenzel 1981, 1991). By the mid-1980s, the advent of the snowmobile significantly increased time spent searching and travelling between seal breathing holes. Data recorded by Wenzel (1991) for 14 winter seal hunts at Clyde River indicated that this active search element of such hunts consumed, on average, 30% of total hunt. A hunter would usually wait at a hole for no more than 30 minutes and then return to his snowmobile to search for other breathing holes. This pattern was confirmed in 2000 (Wenzel n.d.b.). Far from the exemplar sit-and-wait foraging that breathing hole hunting once was, hunters now invest more time searching and travelling between holes. Significantly, this means that narwhal hunting at the floe-edge currently represents a more intensive sit-and-wait foraging activity than mauliqpuq and one of the most intensive sit-and-wait foraging behaviours displayed by Pond Inlet Inuit today. It is interesting to note that the advent of the snowmobile led to the shift from a sit-and-wait mode of foraging to an active one in the case of mauliqpuq hunting, but not in the case of narwhal hunting.
B) Basking Seal Hunting (uuttuq)
Uuttuq commences when two changes occur in the North Baffin sea ice environment. The first is that daylight is lengthening and daytime temperatures begin to melt the snow cover. The other is that this warming collapses the ice domes that cover the seals’ agluit, exposing these openings and also widening them. As these conditions begin to be dominant, virtually any bright-hazy and near windless conditions will elicit ringed seals to come out through these exposed breathing holes onto the surface of the ice. Similarly, as pressure and tidal cracks widen, seals will use these openings to gain the ice.
By mid to late May, these physical conditions mitigate an almost complete end to mauliqpuq sealing and a shift to uuttuq. With respect to the amount of active searching as opposed to sit-and-wait foraging involved in modern breathing hole hunting, uuttuq sealing could be perceived as a perfect example of the active mode of foraging; indeed, it is only when a hunter sights a seal that the pace slows.
The most basic element in this mode of foraging, whether done by dog team or snowmobile, is near-constant movement. As seals can be seen at distances up to two kilometres, hunters are always scanning the area around them as they move. Dog team hunters frequently stood on their sleds to gain a better view as the dogs moved along, while snowmobilers will kneel on the seats of their machines as they drive. The only interruption from searching in basking seal hunting is when an animal is spotted and a stalk is begun. When dog teams were in use, the actual approach segment of the hunt required that the hunter dismount and either crawl toward the animal or carry out a slow walk while holding a white cloth screen in front of him until he was in shooting range. Approaching on dog team meant that a seal, alerted by the team’s barking, would escape before a hunter came within shooting range so hunters often walked as much as one-half kilometre, with a stalk lasting 15 to 20 minutes (Wenzel 1991).
Not surprisingly, the snowmobile has also precipitated a change in this type of sealing affecting both the search and pursuit phases of uuttuq hunting. Change is most notable in the pursuit phase, as hunters no longer try to approach seals on foot (Wenzel 1991). Instead they cover the snowmobile’s hood with either a white cloth or a white plywood board and drive directly at the seal, stopping at about 40-50 metres distance to shoot. Such a method would seem counter-intuitive, especially as it is in such contrast to the quiet, slow foot approach, but observations, and continued use, suggest that it is almost as effective. Frequently overlooked, however, is the effect that snowmobiles have had upon the search phase of this hunt. The snowmobile’s speed allows hunters to cover far more area than was previously possible with dogs. Given that the nature of basking seal hunting is to search until an animal is sighted and because dog teams were extremely slow, much of a hunter’s time in the pre-snowmobile era was consumed by the pace dictated by dog use and the need to carry out a stalk on foot. Indeed, the snowmobile has made uuttuq hunting a process of nearly constant movement in sharp contrast to floe-edge narwhal hunting.
Conclusion
This study of foraging mode provides a number of insights into the nature of the spring floe-edge hunt. Our analysis revealed various features of Inuit hunting behaviour that would have otherwise been excluded in an optimal foraging modeled study. For example, we were able to reveal and investigate the role of the initial phase of active search upon arrival at the floe-edge. It represented an essential process of Inuit environmental and campsite sampling. Furthermore, previous optimal foraging diet breadth studies have treated environments such as the floe-edge as one relatively homogenous habitat to be compared with different habitats (Smith 1991). One purpose of this study was to show how foraging theory can contribute to our understanding of hunting behaviour within one habitat by displaying the heterogeneity that exists within that habitat. For example, the use of embayments suggests that the floe-edge is anything but homogenous in terms of its campsite and narwhal hunting value.
Last, this research suggests that Inuit ecological knowledge remains an essential aspect of the spring floe-edge narwhal hunt. This knowledge includes information specific to narwhals and their behaviour under a range of physical and human-affected conditions. It is critical given the rapid manner in which the floe-edge can be adversely changed by wind and currents, and also by the rapid manner in which technologies change. This knowledge is also obviously important in terms of utilizing the various technologies now available to hunters to the greatest effect.
With regard to spring floe-edge whaling, the dynamic effect that this particular environmental situation imposes on foragers, even when they are equipped with new technologies (e.g., rifles, snowmobiles), is worthy of note. In contrast to the winter and spring seal foraging situations that have been described, these technologies appeared to offer little advantage relative to the effect they have had upon both mauliqpuq and uuttuq sealing. This is the case despite the fact that the rifle appears to be beneficial in extending a narwhal hunter’s striking range; this, however, is offset by the fact that narwhals killed or wounded even a few tens of metres from the floe-edge may be irretrievable. In fact, the data suggest that when whaling at the floe-edge, Inuit are still reliant on traditional knowledge of the habitat, on endurance in the face of difficult physical conditions, and on individual patience. As much as the snowmobiles and rifle have “modernized” floe-edge whaling for Pond Inlet hunters, the foraging mode still practiced is essentially unchanged from that of the pre-snowmobile and pre-rifle “era.” The impact of the snowmobile on narwhal hunting has been to extend the area that Pond Inlet Inuit use to hunt narwhals. This is a significant change as hunters can now venture into areas that were not previously utilized on a continuous basis. However, the foraging mode having reached these areas has not significantly changed.
Parties annexes
Acknowledgements
This research would have been impossible without the generous cooperation of the Inuit of Pond Inlet—especially the hunters who allowed a Qallunaaq to accompany them, the elders who acceded to be interviewed, and the Pond Inlet Hunters and Trappers Organization. Dozens of persons deserve personal thanks. To all Mittimatalingmiut, many thanks. We would like to thank the following families for their invaluable assistance: Agnetsiak, Anaviapik, Enookoolook, Inootik, Inuaraq, Kautainuk, Kiliktee, Komangapik, Koonoo, Kyak, Maktar, Merkosak, Milton, Qamaniq, Alooloo, Nuturak, Paniloo, Peterloosie, Pewatoaluk, Sangoya, and Tigilluraq. Finally, the following individuals provided important logistical support: Lynn Cousins, David Parks and family, and Valentina de Krom. We apologize for any people that we may have missed and we sincerely thank the community, Hamlet, Hunters and Trappers Organization, and Elders Group of Pond Inlet for their tremendous support.
Thanks also to O. Coomes, J. M. Savelle and D. L. Kramer for their guidance, and to J. Caouette for her thoughtful comments and invaluable editing assistance and to Prof. J. Ramsay for statistical assistance. This work was supported by the Max Bell, David Stewart Memorial and J.W. McConnell Fellowships of McGill University, Department of Indian Affairs and Northern Development (DIAND) Northern Scientific Training Program (NSTP) and the Social Science and Humanities Research Council of Canada (SSHRC). Despite the number of contributors to this paper, the views expressed are solely the authors'.
Note
-
[1]
It should be noted that this may be an overestimation since the hunter could have perceived the participant observer as a member of the hunt group.
References
- BAFFIN REGIONAL INUIT ASSOCIATION, 1982 Annual Report of the Baffin Regional Inuit Association, Iqaluit.
- BAFFIN REGIONAL INUIT ASSOCIATION, 1983 Annual Report of the Baffin Regional Inuit Association, Iqaluit.
- BAFFIN REGIONAL INUIT ASSOCIATION, 1984 Annual Report of the Baffin Regional Inuit Association, Iqaluit.
- BAFFIN REGIONAL INUIT ASSOCIATION, 1985 Annual Report of the Baffin Regional Inuit Association, Iqaluit.
- BERNIER, Joseph E., 1909 Report on the Dominion Government Expedition to Arctic islands and the Hudson Strait on board the C.G.S. “Arctic” 1906-1907, Ottawa, Dept. of Marine and Fisheries, C.H. Parmalee.
- BERNIER, Joseph E., 1911 Report on the Dominion Government Expedition to the Northern Waters and Arctic Archipelago of the D.G.S. “Arctic” in 1910, Ottawa, Dept. of Marine and Fisheries, C.H. Parmalee.
- BISSETT, Don, 1968 Northern Baffin Island: An Area Economic Survey, A.E.S.R. no. 67/1, Ottawa, Industrial Division, Dept. of Indian Affairs and Northern Development.
- BOAS, Franz, 1888 The Central Eskimo, Sixth Annual Report of the Bureau of American Ethnology, 1884-85, Washington, Government Printing Office.
- BRODY, Hugh, 1975 The People’s Land: Eskimos and Whites in the Eastern Arctic, Harmondsworth, England, Penguin Books.
- COSENS, Susan E. and Larry P. DUECK, 1988 Responses of Migrating Narwhal and Beluga to Icebreaker Traffic at the Admiralty Inlet Ice-Edge, N.W.T. in 1986, in W.M. Sackinger and M.O. Jeffries (eds), Port and Ocean Engineering under Arctic Conditions, Fairbanks, University of Alaska, Alaska Geophysical Institute: 39-54.
- DAHL, Jens, 1990 Beluga hunting in Saqqaq, North Atlantic Studies, 2(1): 166-169.
- DAHL, Jens, 2000 Saqqaq: An Inuit hunting community in the modern world, Toronto, University of Toronto Press.
- DAMAS, David, 2002 Arctic Migrants/Arctic Villagers, Montreal, McGill-Queen's University Press.
- DEGERBØL, Magnus and Peter FREUCHEN, 1935 Report of the Mammals Collected by the Fifth Thule Expedition to Arctic North America: Part I. Systematic Notes, Copenhagen, Gyldendalske Boghandel, Report of the Fifth Thule Expedition, 2(4): 1-67.
- DONALDSON, Judith L., 1994 The Economic Ecology of Hunting: A Case Study of the Canadian Inuit, Ph.D. thesis, Harvard University.
- FINLEY, Kerry J., Rolph A. DAVIS and Helen B. SILVERMAN, 1980 Aspects of the Narwhal Hunt in the Eastern Canadian Arctic, Report of the International Whaling Commission, 30: 459-464.
- FINLEY, Kerry J. and Gary W. MILLER, 1982 The 1979 Hunt for Narwhals (Monodon monoceros) and an Examination of Harpoon Gun Technology near Pond Inlet, Northern Baffin Island, Report of the International Whaling Commission, 32: 449-460.
- FINLEY, Kerry J., Gary W. MILLER, Rolph A. DAVIS and Charles R. GREENE, 1990 Reactions of Belugas (Delphinapterus leucas) and Narwhals (Monodon monoceros) to Ice-Breaking Ships in the Canadian High Arctic, Can. Bull. Fish. Aquatic Sci., 224: 97-117.
- FORD, John, K.B., 1976 Underwater Acoustic Signals of the Narwhal (Monodon monoceros), B.Sc. thesis, Vancouver, University of British Columbia.
- HAMES, Raymond B. and William T. VICKERS, 1982 Optimal Foraging Theory as a Model to Explain Variability in Amazonia Hunting, American Ethnologist, 9: 358-378.
- HAWKES, Kristen and James F. O’CONNELL, 1982 Why Hunters Gather: Optimal Foraging and the Aché of East Paraguay, American Ethnologist, 9: 379-398.
- HAWKES, Kristen and James F. O’CONNELL, 1985 Optimal Foraging Models and the Case of the !Kung, American Anthropologist, 87: 401-426.
- HAWKES, Kristen and James F. O’CONNELL, 1992 On Optimal Foraging Models and Subsistence Transitions, Current Anthropology, 33: 63-66.
- HAWKES, Kristen, James F. O’CONNELL and Nicholas G. BLURTON JONES, 1991 Hunting Income Patterns among the Hadza: Big Game, Common Goods, Foraging Goals, and the Evolution of the Human Diet, Philosophical Transactions of the Royal Society, 334: 243-251.
- HAY, Keith, 1984 The Life History of the Narwhal (Monodon monoceros L.) in the Eastern Canadian Arctic, Ph.D. dissertation, Department of Oceanography, McGill University.
- HILL, Kim, 1988 Macronutrient Modifications of Optimal Foraging Theory: An Approach Using Indifference Curves Applied to Some Modern Foragers, Human Ecology, 16: 157-197.
- HOLLING, Crawford S., 1959 The Components of Predation as Revealed by a Study of Small-Mammal Predation of the European Pine Sawfly”, Can. Entomol., 91: 293-320.
- HUEY, Raymond B. and Eric R. PIANKA, 1981 Ecological Consequences of Foraging Mode, Ecology, 62: 991-999.
- JENNESS, Diamond, 1922 The Life of the Copper Eskimos, Ottawa, Report of the Canadian Arctic Expedition, 1913-1918, 12.
- KINGSLEY, Michael C.S., Holly J. CLEATOR and Michael A. RAMSAY, 1994 Summer Distribution and Movements of Narwhals (Monodon, monoceros) in Eclipse Sound and Adjacent Waters, North Baffin Island, N.W.T., Meddelelser øm Grønland:Bioscience, 39: 163-174.
- KOSKI, William R. and Rolph A. DAVIS, 1994 Distribution and Numbers of Narwhals (Monodon monoceros) in Baffin Bay and Davis Strait, Meddelelser øm Grønland: Bioscience, 39: 15-40.
- KRAMER, Donald L., 2001 “Foraging Behavior”, in C.W. Fox, D.A. Roff and D.J. Fairbairn (eds) Evolutionary Ecology: Concepts and Case Studies, New York, Oxford University Press: 232-246.
- LAND, E.M., 1977 The Narwhal and the Walrus: A Problem of Ivory in T. Mosquin and C. Suchal (eds), Proceedings of the Symposium on Canada’s Threatened Species and Habitats Co-sponsored by the Canadian Nature Federation and the World Wildlife Fund (Canada) held in Ottawa May 20-24, 1976, Ottawa, Canadian Nature Federation, Special Publication, 6: 79-81.
- MARY-ROUSSELIÈRE, Guy, 1984 Iglulik, in David Damas (ed.), Handbook of North American Indians, vol. 5, Arctic, Washington, Smithsonian Institution: 431-475.
- MATHIASSEN, Therkel, 1928 Material Culture of the Iglulik Eskimos, Copenhagen, Gyldendalske Boghandel, Report of the Fifth Thule Expedition, 6(1).
- MATTHIASSON, John, 1967 Eskimo Legal Acculturation: The Adjustment of Baffin Island Eskimos to Canadian Law, Ph.D. dissertaion, Ithaca, Cornell University.
- MATTHIASSON, John, 1992 Living on the Land: Change among the Inuit of Baffin Island, New York, Broadview.
- PARKINSON, Claire L., 1999 Arctic Sea Ice Extents, Areas and Trends, 1978-1996, J. Geophys. Res., 104: 20837-20856.
- PARKINSON, Claire L., 2000 Variability of Arctic Sea Ice: The View from Space, an 18-year Record, Arctic, 53: 341-358.
- RASMUSSEN, Knud Johan Victor, 1929 Intellectual Culture of the Iglulik Eskimos, Copenhagen, Nordisk forlag, Report of the Fifth Thule Expedition 1921-24, 7(1).
- RASMUSSEN, Knud Johan Victor, 1931 The Netsilik Eskimos: Social Life and Spiritual Culture, Copenhagen, Nordisk forlag, Report of the Fifth Thule Expedition 1921-1924, 8.
- REEVES, Randall, 1992 What is a Narwhal Worth?, Ph.D. dissertaion, Montreal, McGill University.
- REMNANT, Richard A. and M.L. THOMAS, 1992 Inuit Traditional Knowledge of the Distribution and Biology of High Arctic Narwhal and Beluga, Winnipeg, North/South Consultants.
- RICHARD, P., P. WEAVER, Larry DUECK and D. BARBER, 1994 Distribution and Numbers of Canadian High Arctic Narwhals (Monodon monoceros) in August 1984, Meddelelser øm Grønland: Bioscience, 39: 41-50.
- ROBERGE, Michelle M. and J.B. DUNN., 1990 Assessment of the Subsistence Harvest and Biology of Narwhal (Monodon monoceros L.) from Admiralty Inlet, Baffin Island, N.W.T. 1983 and 1986-89, Can. J. Fish. Aquat. Sci., Technical report 1747.
- SAVELLE, James M., 1994 Prehistoric Exploitation of White Whales (Delphinapterus leucas) and Narwhals (Monodon monoceros) in the Eastern Canadian Arctic, Meddelelser øm Grønland: Bioscience, 39: 101-117.
- SIH, Andrew and Katherine MILTON, 1985 Optimal Diet Theory: Should the !Kung Eat Mongongos?, American Anthropologist, 87: 396-401.
- SILVERMAN, Helen, 1979 Social Organization of the Narwhal, Monodon monoceros, in Lancaster Sound, Pond Inlet, and Tremblay Sound, Northwest Territories, M.Sc. thesis, Montreal, McGill University.
- SMITH, Eric Alden, 1991 Inujjamiut Foraging Strategies: Evolutionary Ecology of an Arctic Hunting Economy, New York, Walter de Gruyter.
- SMITH, Thomas.G., Michael O. HAMMIL, D.J. BURRAGE, and G.A. SLENO, 1985 Distribution and Abundance of Belugas, Delphinapterus leucas, and Narwhals, Monodon monoceros, in the Canadian High Arctic, Can. J. Fish. Aquat. Sci., 42: 676-684.
- STEWART, David B., A. Akeeagok, S. Amarulalik, S. Panipakutsuk, and A. Taqtu, 1995 Local knowledge of Beluga and Narwhal from Four Communities in Arctic Canada, Can. Tech. Rep. Fish. Aquat. Sci., Technical report 2065.
- STRONG, John T., 1988 Status of the Narwhal, Monodon monoceros in Canada, Can. Field-Nat., 42: 676-684.
- THOMSEN, M.L., 1993 Local Knowledge of the Distribution, Biology, and Hunting of Beluga and Narwhal: A Survey among Inuit hunters in West and North Greenland. Nuuk: Greenland Hunters’ and Fishermen’s Association, Nuuk, Greenland Home Rule Authorities and Inuit Circumpolar Conference.
- TREUDE, Erhard, 1977 Pond Inlet, Northern Baffin Island: The Structure of an Eskimo Resource Area, Polar Geography, 1: 95-122.
- WEAVER, P.A. and R.S. WALKER, 1988 The Narwhal (Monodon monoceros L.) Harvest in Pond Inlet, Northwest Territories: Hunt Documentation and Biological Sampling 1982-1983, Can. Manuscript Rep. of Fisheries and Oceans, Technical report 1975.
- WENZEL, George, 1991 Animal Rights, Human Rights: Ecology, Economy, and Ideology in the Canadian Arctic, Toronto, University of Toronto Press.
- WINTERHALDER, Bruce, 1977 Foraging Strategy Adaptations of the Boreal Forest Cree: An Evaluation of Theory and Models from Evolutionary Ecology, Ph.D. dissertation, Ithaca, Cornell University.
- WINTERHALDER, Bruce, 1983 Boreal Foraging Strategies, in A. Theodore Steegmann, Jr. (ed.), Boreal Forest Adaptations: The Northern Algonkians, New York, Plenum Press: 201-241.
- WINTERHALDER, Bruce and Eric Alden SMITH, 2000 Analyzing Adaptive Strategies: Human Behavioral Ecology at Twenty-five, Evolutionary Anthropology, 9: 51-72.
Liste des figures
Figure 1
Location of Pond Inlet (Mittimatalik) and study area.
Figure 2
Spring floe edge Inuit use area and narwhal migration routes.
Figure 3
Sit-and-wait (cross-hatched bars) and active (white bars) foraging behavior distributions for each trip relative to total foraging time is presented. The sit-and-wait mode is clearly the most dominant search behavior in all participant observed floe-edge narwhal trips.
Liste des tableaux
Table 1
Inuit floe-edge narwhal foraging cycle.
Table 2
Social and demographic characteristics of hunting groups.
Table 3
Number and duration of active foraging events (AFE) based from campsite and total duration of all active foraging (all duration in minutes).
*Includes active foraging before first camp, AFE, and foraging between campsites.
Table 4
Number of camps established and camp residence duration, and total duration of all sit-and-wait foraging (all duration in minutes).
*Includes camp residence duration but not active foraging activity based from campsite (see AFE in Table 3).
Table 5
Duration of sit and wait vs. active foraging activity (in minutes).
Table 6
Floe-edge narwhal sightings and firing events.