
Résumés
Résumé
L’émotion est une « arme à double tranchant » qui peut soit améliorer, soit détériorer des aspects divers de notre cognition et de notre comportement. Par exemple, la charge émotionnelle d’un événement peut attirer l’attention et renforcer la mémoire de cet événement, par contre une information émotionnelle, sans pertinence pour la tâche cognitive en cours, peut être considérée comme une distraction et ainsi entraîner la détérioration de la performance à cette tâche. Ces effets opposés de l’émotion peuvent être identifiés à différents niveaux, à la fois au sein du même processus cognitif et à travers des processus cognitifs différents, ou à un niveau plus général, comme dans le cas de la réponse au stress. Cette revue présente des données nouvelles qui permettent de mieux comprendre les facteurs qui influencent les effets opposés de l’émotion sur la cognition chez le sujet sain ainsi que dans le cas de dysfonctionnements liés à des désordres affectifs. Les questions discutées dans cette revue sont d’une grande importance pour arriver à mieux comprendre les mécanismes impliqués dans les relations entre émotion et cognition dans le fonctionnement sain ainsi que dans les troubles émotionnels où les effets de l’émotion sur la cognition sont intensifiés. Globalement, la présente revue souligne la nécessité de considérer et identifier les facteurs qui peuvent avoir une influence sur les effets de facilitation et les effets de dégradation des émotions sur la cognition.
Mots-clés :
- émotion,
- attention,
- mémoire émotionnelle,
- distraction émotionnelle,
- troubles émotionnels
Abstract
Objectives Emerging evidence suggests that emotion can have both enhancing and impairing effects on various cognitive processes. These opposing effects can be identified at different levels, both within the same cognitive process and across different processes, as well as at more general levels, such as in the case of the response to stress. The aim of the present review is to discuss recent advances in the mechanisms underlying the enhancing and impairing effects of emotion on different aspects within the same process (e.g., episodic memory) and across specific cognitive processes (perception vs. episodic memory, working memory vs. episodic memory), as well as in the context of the response to stress.
Emerging Evidence The available evidence points to a number of aspects that dissociate the opposing effects of emotion on cognition. (i) Opposing effects within episodic memory can be attributed to different accounts, involving dissociation at different levels: central vs. peripheral trade-off, high vs. low prioritization of information processing, and items encoding vs. the formation of complex associations. (ii) The opposing effects across cognitive processes, such as perception and episodic memory, can be linked to dissociation between immediate/impairing vs. long-term/enhancing effects, which are mediated by common and dissociable neural mechanisms, involving bottom-up and top-down processes. (iii) Finally, in the larger context of the response to stress, emotional stress can lead to opposing effects depending on the degree, context, and controllability of the stressors.
Conclusions Overall, the present review highlights the need to consider the various factors that can influence enhancing or impairing effects of emotion on cognition, in studies investigating emotion-cognition interactions. These issues are important for understanding mechanisms of emotion-cognition interactions not only in healthy functioning but also in emotional disturbances, where these opposing effects of emotion are exacerbated and deleterious.
Keywords:
- emotion,
- attention,
- emotional memory,
- emotional distraction,
- affective disorders
Corps de l’article
Introduction
L’émotion est une « arme à double tranchant » qui peut soit améliorer, soit détériorer des aspects divers de notre cognition et de notre comportement. Par exemple, la charge émotionnelle d’un événement peut attirer l’attention et renforcer la mémoire de cet événement, par contre, une information émotionnelle sans pertinence pour la tâche cognitive en cours peut être considérée comme une distraction et ainsi entraîner la détérioration de la performance à cette tâche. Ces effets opposés de l’émotion peuvent être identifiés à différents niveaux, à la fois au sein du même processus cognitif et à travers des processus cognitifs différents, et également à un niveau plus général, comme dans le cas de la réponse au stress. Par exemple, le même événement émotionnel peut conduire à des effets opposés sur des aspects différents « au sein du même processus cognitif » (par exemple, la mémoire épisodique). Être menacé avec une arme à feu peut renforcer la mémoire pour les aspects qui sont au coeur de l’événement (par exemple, la couleur de l’arme), tout en altérant la mémoire des détails périphériques (par exemple, la façon dont l’attaquant était habillé ou des détails sur son identité). L’information émotionnelle peut également conduire à des effets opposés « à travers des processus cognitifs différents » : une distraction émotionnelle peut altérer le processus cognitif en cours, tout en améliorant la mémoire à long terme pour la distraction elle-même. Par exemple, être témoin d’un accident tragique pendant que nous conduisons notre voiture peut momentanément nous distraire de la tâche principale (la conduite), tout en facilitant la rétention en mémoire de la scène de l’accident. En plus, dans le contexte plus global de la réponse au stress, les réactions émotionnelles associées à des niveaux optimaux de stress (eustress) peuvent augmenter la performance (par exemple, les émotions positives associées aux préparatifs de mariage), alors que les émotions associées à des niveaux élevés de stress (par exemple, inquiétude désespérante avant un examen difficile) peuvent diminuer la performance. Élucider les effets opposés de l’émotion est d’une grande importance pour arriver à bien comprendre les mécanismes impliqués dans les relations entre émotion et cognition dans le fonctionnement sain, ainsi que dans les troubles émotionnels, où ces effets sont intensifiés et néfastes.
Cette revue présente des données nouvelles permettant de mieux comprendre les facteurs qui influencent les effets opposés de l’émotion sur la cognition. Le premier chapitre présente des données provenant des études ayant dissocié les effets facilitateurs des effets délétères de l’émotion « au sein du même processus cognitif ». Le deuxième chapitre analyse les données provenant des études ayant examiné les effets facilitants et les effets altérants de l’émotion « à travers des processus cognitifs différents ». Enfin, le troisième chapitre présente des données provenant des études ayant examiné l’impact de l’émotion « à un niveau plus général », notamment dans le contexte de la réponse au stress. Cette revue se termine par des conclusions finales et une brève discussion des questions ouvertes et des orientations futures de la recherche.
1. Dissocier les effets opposés de l’émotion au sein de la mémoire épisodique
Alors qu’il existe des preuves solides provenant de la recherche animale (McGaugh, 2004), ainsi que de la recherche humaine (Dolcos, LaBar & Cabeza, 2006) que les événements chargés en émotion sont globalement mieux rappelés que les événements neutres, il y a également des données qui montrent qu’une telle facilitation n’est pas observée pour tous les aspects de l’événement. Les effets de l’émotion sur la mémoire épisodique chez l’homme ont été typiquement étudiés en utilisant des stimuli de laboratoire, notamment des listes de mots ou des séries d’images, qui ont des niveaux différents d’intensité et de valence émotionnelles, et sont encodés et rappelés à des intervalles différents après l’encodage (par exemple, minutes, heures, jours) dans le contexte d’examen en laboratoire. Ces études ont démontré que les effets facilitateurs de l’émotion sur la mémoire peuvent être attribués à l’implication de l’amygdale et son interaction avec le système mnésique du lobe temporal médian (comprenant l’hippocampe, le cortex entorhinal, périrhinal et parahippocampique). De plus, ces effets facilitateurs peuvent également bénéficier de l’engagement des régions cérébrales cognitives d’ordre supérieur (par exemple, le cortex préfrontal et le cortex pariétal) (Dolcos & Denkova, 2008 ; Dolcos et al., 2012).
L’effet de facilitation mnésique par l’émotion a été typiquement observé pour des éléments émotionnels isolés et leurs propriétés intrinsèques, tandis que la mémoire pour des aspects extrinsèques et des détails contextuels de l’événement n’est pas améliorée et peut même être détériorée (Kensinger, 2009 ; Mather, 2007). Plus précisément, la dissociation entre facilitation et détérioration de la mémoire par l’émotion a été posée en termes de dichotomies entre éléments centraux vs éléments périphériques, éléments de haute priorité vs éléments de faible priorité, élément unique vs association d’éléments. Ces dichotomies vont être présentées ci-dessous.
Le compromis entre éléments centraux et éléments périphériques
L’observation que l’émotion peut améliorer la mémoire pour les aspects centraux et la détériorer pour les détails périphériques a été originellement rapportée dans la littérature au sujet des témoignages oculaires. Elle est connue sous le terme de weapon focus désignant la tendance des témoins du crime à se concentrer sur l’arme, en dépit d’autres détails de l’événement (Christianson, 1992 ; Loftus, Loftus & Messo, 1987). Des travaux de recherche plus récents, ayant étudié l’influence de l’émotion sur la mémoire, ont fait référence à ce phénomène en termes de central/peripheral trade-off (compromis entre éléments centraux et éléments périphériques), terme désignant le fait que la mémoire est améliorée pour les éléments centraux et diminuée pour les éléments périphériques (Kensinger, 2009). Cet effet est généralement étudié par la présentation d’images illustrant des items à forte charge émotionnelle dans un contexte neutre (c’est-à-dire que les éléments qui se trouvent dans l’arrière-plan de l’image sont neutres, par exemple un serpent près d’une rivière) et des images illustrant des items à charge émotionnellement neutre dans un contexte neutre (par exemple, un écureuil dans une forêt). Ces études ont montré que les items émotionnels sont mieux rappelés que les items neutres, alors que le contexte associé avec un item émotionnel est moins bien rappelé que le contexte associé avec un item neutre (Figure 1) (Kensinger, Garoff-Eaton & Schacter, 2007 ; Waring, Payne, Schacter & Kensinger, 2010). Du point de vue neuronal, l’amygdale a été impliquée uniquement dans l’amélioration de la mémoire pour les éléments qui sont intrinsèquement liés à l’item émotionnel lui-même, comme son apparence physique, mais pas pour tous les éléments extrinsèques d’un événement émotionnel, comme son contexte (Dougal, Phelps & Davachi, 2007 ; Kensinger, Garoff-Eaton & Schacter, 2007 ; Kensinger & Schacter, 2006).
Figure 1

Taux de reconnaissance correcte pour l’item central et le contexte (l’arrière-plan) de l’image démontrant le compromis entre éléments centraux et éléments périphériques. Alors que les items négatifs ont été mieux rappelés que les items neutres, le contexte associé avec un item émotionnel a été moins bien rappelé que le contexte associé avec un item neutre.
Le rôle des processus de priorisation dans l’impact de l’émotion sur la mémoire
Des preuves supplémentaires suggèrent que les effets opposés de l’émotion sur la mémoire peuvent être liés à des processus de priorisation, comme le souligne la théorie de la compétition biaisée par le niveau d’activation (Arousal-Biased Competition Theory, Mather & Sutherland, 2011). Selon ce modèle, l’activation émotionnelle améliore l’encodage des informations de haute priorité au détriment des informations de faible priorité (Mather & Sutherland, 2011). Dans une série d’études, Mather et ses collègues ont montré que la présentation d’un item émotionnel peut améliorer la mémoire de l’item précédent émotionnellement neutre, si ce dernier était privilégié (item précédent prioritaire), par contre, la présentation d’un item émotionnel peut diminuer la mémoire de l’item précédent émotionnellement neutre, si ce dernier n’a pas été privilégié (item précédent non-prioritaire) (Figure 2) (Lee, Itti & Mather, 2012 ; Lee, Sakaki, Cheng, Velasco & Mather 2014 ; Sakaki, Fryer & Mather, 2014). À noter que la priorité peut être attribuée soit par la saillance ascendante (bottom-up) du stimulus, soit par sa pertinence (top-down) pour le but présent (Lee et al., 2012 ; Sakaki, Fryer et al., 2014). En matière neuronale, les données mettent en évidence une dissociation concernant l’implication de l’amygdale qui dépend de ce que l’information soit prioritaire ou non (Lee et al., 2014). Surtout, la théorie de la compétition biaisée par le niveau d’activation peut être liée à des modèles qui prennent en compte des facteurs de motivation pour comprendre l’impact de l’émotion sur la mémoire (Levine & Edelstein, 2009 ; Sander, Grandjean & Scherer, 2005).
Figure 2

L’effet de priorisation au niveau de la performance mnésique lors d’un test de mémoire où les participants doivent retenir des images d’objets en mémoire en portant leur attention soit sur un objet encadré, soit sur l’objet qui le précède. Comparées à des images neutres, les images à charge émotionnelle négative améliorent la mémoire pour l’image précédente neutre (Objet -1) quand celle-ci est privilégiée lors de l’encodage (Objet -1 prioritaire), et non pas quand elle n’est pas privilégiée (Objet prioritaire).
Le rôle de l’unification vs les associations complexes d’éléments dans l’impact des émotions sur la mémoire
Une autre explication possible des effets opposés de l’émotion sur la mémoire épisodique (Chiu, Dolcos, Gonsalves & Cohen, 2013) peut être liée à la dissociation entre la mémoire pour des éléments isolés, par opposition à la mémoire pour les relations entre les éléments (la mémoire associative) (Cohen et al., 1999 ; Cohen & Eichenbaum, 2001). Un nombre croissant d’arguments empiriques provenant de la recherche animale, ainsi que de la recherche humaine, suggère que des régions différentes du système mnésique du lobe temporal médian peuvent jouer un rôle différentiel dans la mémoire pour un objet isolé en comparaison de la mémoire des associations (un exemple de mémoire d’objet isolé peut être le souvenir d’une raquette de tennis, alors qu’un exemple de mémoire associative peut être l’association entre l’objet et sa couleur, sa taille ou son contexte, comme la raquette rouge dans le coffre d’une voiture). Alors que le cortex périrhinal joue un rôle important dans l’encodage d’éléments isolés, l’hippocampe joue un rôle fondamental pour relier entre elles des représentations distinctes en mémoire (Davachi, Mitchell & Wagner, 2003 ; Ranganath, Cohen, Dam & D’Esposito, 2004 ; Tubridy & Davachi, 2011). D’autres études ont révélé que le cortex périrhinal peut également contribuer à certaines formes simples d’apprentissage associatif (Staresina & Davachi, 2010), basées sur l’unification (unitization), ce qui implique la représentation de composantes distinctes en une seule unité (Graf & Schacter, 1989). L’unification implique, par exemple, l’assemblage de différents aspects d’un objet en une représentation unique (comme l’association entre un objet et sa couleur, dans l’exemple ci-dessus – la raquette de tennis rouge). Par conséquent, la mémorisation d’éléments isolés ou d’items unifiés peut être sous-tendue par des mécanismes similaires. Par contre, la mémorisation d’associations plus complexes impliquant des éléments différents et variés d’un événement, ainsi que celle d’associations entre événements temporellement séparés, dépend de mécanismes sous-tendus par l’hippocampe (Ezzyat & Davachi, 2014). En considérant ces données, Chiu et al. (2013) proposent que l’émotion conduit à l’amélioration de la mémoire pour des items isolés ainsi que des items unifiés, mais elle détériore la mémoire de représentations plus complexes, celle-ci étant sous-tendue par l’hippocampe. Cependant, cette proposition nouvelle n’a pas été testée directement, de manière empirique (mais voir Luck, Lecleck & Lepage, 2014). Une façon potentielle de la tester serait de manipuler à la fois la valence émotionnelle des objets (par exemple, pistolet vs une raquette de tennis) et les demandes d’association qui peuvent inclure d’une part l’unification, impliquant la « fusion » d’un objet avec sa couleur en une seule représentation (pistolet argenté) et d’autre part, des associations complexes, impliquant des informations liées aux aspects spatiaux, situationnels et temporels de la scène (comme le pistolet dans le taxi, à la station d’essence, tôt le matin).
Globalement, les effets opposés de l’émotion sur la mémoire épisodique ont été identifiés en tant que dichotomies entre éléments centraux vs éléments périphériques, éléments de haute priorité vs éléments de faible priorité et élément unique vs association d’éléments. Des recherches futures devraient examiner ces dissociations, mieux délimiter l’impact de l’émotion selon le type d’association, ainsi qu’en fonction de représentations préexistantes et identifier les corrélats neuronaux sous-jacents (Sakaki, Ycaza-Herrera & Mather, 2014).
2. Dissocier les effets opposés de l’émotion à travers des processus cognitifs différents
De nouvelles données suggèrent que les effets opposés de l’émotion peuvent être identifiés en fonction de la distinction entre effet « immédiat » (dégradant) et effet « à long terme » (améliorant) de l’information émotionnelle de nature distractive à travers des processus cognitifs différents. Plus précisément, il a été démontré que des distracteurs émotionnels, qui ne sont pas pertinents pour le processus cognitif en cours, peuvent altérer le traitement cognitif (par exemple, la perception), tout en conduisant à l’amélioration de la mémoire pour les distracteurs eux-mêmes. Ces études permettent d’identifier les bases neuronales impliquées dans les effets de l’émotion dans des situations proches de la vie réelle, où l’information émotionnelle non pertinente pour l’activité en cours (la scène d’un accident tragique pendant la conduite de voiture) peut temporairement nous distraire de la tâche principale (la conduite), tout en menant à une meilleure mémoire pour le distracteur (voiture accidentée).
Les effets opposés de l’émotion sur la perception et la mémoire épisodique
Des données récentes suggèrent que la distraction émotionnelle peut avoir des effets néfastes immédiats sur le traitement perceptif de bas niveau tout en conduisant à l’amélioration à long terme de la mémoire pour les distracteurs eux-mêmes (Shafer & Dolcos, 2012). En particulier, cette étude a démontré que l’effet dégradant immédiat et l’effet bénéfique à long terme sont différemment influencés par la disponibilité des ressources de traitement. Plus précisément, alors que la dégradation la plus robuste du traitement perceptif est observée lorsque peu de ressources sont sollicitées pour le traitement cognitif en cours (charge basse), et donc qu’il y a plus de ressources de traitement disponibles, l’amélioration la plus robuste de la mémoire pour les distracteurs émotionnels est produite lorsque les ressources de traitement sont moins disponibles (charge élevée). Du point de vue neuronal, la dissociation entre effet altérant immédiat et effet facilitant à long terme a été observée au niveau des mécanismes de base impliquant l’amygdale et l’hippocampe d’une part, ainsi qu’au niveau des régions cérébrales impliquées dans des processus d’ordre supérieur d’autre part (cortex préfrontal médian et cortex pariétal). Plus précisément, les résultats indiquent une dissociation hémisphérique au niveau des mécanismes impliquant l’amygdale et l’hippocampe, avec un engagement bilatéral pour l’effet altérant et un engagement du côté gauche pour l’effet de facilitation mnésique par l’émotion (Figure 3). Ce résultat peut être lié à des travaux préalables suggérant un engagement plus prononcé de l’amygdale gauche lors de traitements plus élaborés de stimuli émotionnels, ce qui contribue également à une facilitation de la mémoire (Glascher & Adolphs, 2003 ; Phelps et al., 2001).
Il est intéressant de noter que ces dissociations ont été observées dans des sous-régions de l’amygdale et de l’hippocampe impliquées dans les deux effets, mais des régions cérébrales en dehors du lobe temporal médian ont également présenté des modèles de dissociation en fonction des effets opposés. Plus précisément, le cortex préfrontal médian a été associé avec l’effet altérant immédiat au niveau du processus perceptif, alors que le cortex pariétal supérieur a été associé avec l’effet facilitant à long terme sur la mémoire (Figure 3). Étant donné que le cortex préfrontal médian est sensible aux stimuli émotionnels (Keightley et al., 2003 ; Scheuerecker et al., 2007) et que le cortex pariétal supérieur fait partie du réseau attentionnel descendant impliqué dans l’évaluation et la sélection des informations pertinentes (Corbetta & Shulman, 2002), leur engagement spécifique pour chacun des effets peut être respectivement attribué à une augmentation du traitement émotionnel et de l’évaluation de la pertinence des distracteurs.
Figure 3

Dissociation entre effet immédiat et effet à long terme de la distraction émotionnelle : (a) Vue coronale de l’amygdale illustrant l’activation bilatérale liée à l’effet altérant immédiat (en gris foncé) et l’activation gauche liée à l’effet facilitant à long terme (en blanc) ; à noter que l’activation d’une sous-région de l’amygdale gauche est positivement corrélée avec la performance mnésique ; (b) Vue en 3-D du cerveau illustrant l’implication du cortex préfrontal médian dans l’effet altérant immédiat (en gris foncé) et l’implication du cortex pariétal dans l’effet facilitant à long terme (en blanc). D = hémisphère droit, G = hémisphère gauche.
Des effets opposés de l’émotion sur la mémoire de travail et la mémoire épisodique
La distraction émotionnelle peut produire des effets néfastes non seulement au niveau du traitement perceptif primaire, mais aussi au niveau de traitements plus complexes, comme la mémoire de travail (Dolcos, Diaz-Granados, Wang & McCarthy, 2008 ; Dolcos & McCarthy, 2006 ; Dolcos, Miller, Kragel, Jha & McCarthy, 2007). Les résultats d’une étude récente ayant examiné les effets opposés des distracteurs émotionnels chez les mêmes participants ont démontré que les distracteurs émotionnels présentés pendant l’intervalle de temps entre la mémorisation et la récupération dans une tâche de mémoire de travail ont un effet altérant immédiat sur la performance, de mémoire de travail, tout en améliorant la mémoire à long terme (la mémoire épisodique) pour les distracteurs (Dolcos et al., 2013). Ceci suggère que les distracteurs émotionnels détournent les ressources de traitement de la tâche principale et ainsi détériorent la performance, tout en engendrant un traitement qui conduit à une meilleure mémoire pour les distracteurs eux-mêmes (Dolcos & McCarthy, 2006).
Au niveau neuronal, les essais expérimentaux liés aux deux effets opposés (mémoire de travail détériorée et mémoire épisodique améliorée) ont été associés à une diminution de l’activité dans le cortex préfrontal (CPF) dorso-latéral (liée à l’effet négatif immédiat sur la performance de mémoire de travail) vs une augmentation de l’activité dans les régions du lobe temporal médian (liée à la performance accrue de mémoire épisodique) (Figure 4). Il est intéressant de noter que la même région de l’amygdale était liée aux deux effets, de même que les essais expérimentaux associés à une performance améliorée en mémoire épisodique sans détérioration de la performance en mémoire de travail ont été liés à un engagement prononcé des mécanismes descendants du cortex préfrontal (cortex préfrontal ventro-latéral). Ceci suggère que l’amélioration de la mémoire pour les distracteurs émotionnels bénéficie également de l’engagement de stratégies d’adaptation pour faire face à la présence de ces distracteurs au cours de la tâche de mémoire de travail (Dolcos et al., 2013), ce qui peut refléter un encodage approfondi dû à un traitement plus élaboré des distracteurs (Dillon, Ritchey, Johnson & Labar, 2007).
Figure 4
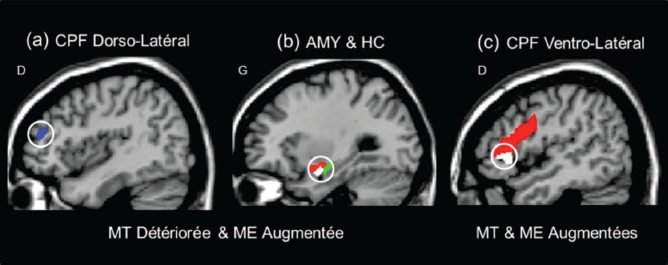
Les activations cérébrales liées à la détérioration de la mémoire de travail (MT) et/ou l’amélioration de la mémoire épisodique (ME) par la distraction émotionnelle : (a) Désactivation plus prononcée dans le cortex préfrontal (CPF) dorso-latéral (en bleu) ; (b) Activation plus prononcée dans l’amygdale (AMY, en rouge) et l’hippocampe (HC, en vert) liée à une mémoire de travail détériorée et une mémoire épisodique améliorée ; (c) Activation plus prononcée dans le cortex préfrontal ventro-latéral (en rouge) liée à l’amélioration de la mémoire de travail et mémoire épisodique. Il est intéressant de noter que des sous-régions de l’amygdale et du cortex préfrontal ventro-latéral (en blanc) contribuent différemment à l’effet de la distraction émotionnelle sur la mémoire de travail, l’amygdale étant liée à une performance détériorée (corrélation négative) et le cortex préfrontal ventro-latéral étant lié à une performance améliorée (corrélation positive). D = hémisphère droit, G = hémisphère gauche.
Collectivement, ces résultats démontrent que l’effet altérant immédiat de la distraction émotionnelle sur la perception ou la mémoire de travail et l’effet facilitant à long terme au niveau de la mémoire épisodique sont médiés par des systèmes neuronaux dissociables, ainsi que des mécanismes communs, impliquant des processus ascendants (bottom-up) et des processus descendants (top-down).
3. Dissocier les effets opposés de l’émotion à la réponse au stress
L’impact de l’émotion sur la cognition peut être également étudié dans le contexte de la réponse au stress. Les expériences stressantes déclenchent l’activation de l’axe hypothalamus-hypophyso-surrénalien (Joels & Baram, 2009 ; Lupien, Maheu, Tu, Fiocco & Schramek, 2007), qui affecte le fonctionnement des régions du cerveau impliquées dans les émotions (amygdale) (Roozendaal, McEwen & Chattarji, 2009) et dans les processus cognitifs (hippocampe et cortex préfrontal) (Lupien, McEwen, Gunnar & Heim, 2009 ; Roozendaal et al., 2009). Ces régions sont parmi celles qui sont les plus sensibles aux hormones du stress, à cause de la forte densité des récepteurs glucocorticoïdes et elles sont aussi les principales structures cérébrales impliquées dans l’apprentissage et la mémoire émotionnels. De nombreuses études provenant de la recherche chez l’animal et l’homme montrent que le stress peut avoir des effets à la fois bénéfiques et néfastes sur l’apprentissage et la mémoire (Lupien et al., 2007 ; McEwen, 2007 ; Roozendaal et al., 2009). Comme nous le verrons ci-dessous, les effets spécifiques du stress sur la cognition ont été liés à divers facteurs, comprenant la durée de l’exposition (aiguë vs chronique), le niveau de stress (élevé vs bas), ainsi que le niveau de contrôlabilité du stress.
Stress aigu vs chronique et niveau de stress élevé vs optimal
Le stress aigu peut conduire à l’hyperexcitation « transitoire », ce qui favorise la détection des menaces et la mémorisation d’événements émotionnels grâce à l’implication de l’amygdale et de ses connexions avec les structures cérébrales associées à la mémoire (McGaugh, 2000, 2004), ceci pouvant avoir des fonctions adaptatives. En revanche, l’exposition répétée au stress (stress chronique) peut conduire à un état d’excitation physiologique « continue » et peut ainsi avoir des effets délétères sur l’hippocampe et le cortex préfrontal (Roozendaal et al., 2009 ; Arnsten, 2000, 2009 ; Hains & Arnsten, 2008), conduisant à des conséquences mal adaptées. Des données convergentes entre les études chez l’homme et chez l’animal suggèrent que l’effet du stress aigu sur la cognition, en particulier sur la mémoire, suit une fonction en forme d’U inversé, avec un niveau modéré de stress conduisant à l’amélioration de la mémoire et les niveaux de stress trop bas ou trop élevés conduisant à la réduction de la mémoire (Diamond, Campbell, Park, Halonen & Zoladz, 2007 ; Park et al., 2006 ; Sandi & Pinelo-Nava, 2007). Fait intéressant, des effets similaires ont été également observés au niveau de l’hippocampe, en réponse au stress (fonction en forme d’U inversé) (Nadel & Jacobs, 1998). Il est important de noter que des événements d’une intensité émotionnelle très aiguë et l’exposition chronique à des expériences stressantes peuvent conduire à des désordres affectifs tels que la dépression et le trouble de stress post-traumatique.
Présence vs absence de contrôlabilité dans la réponse au stress
Des travaux récents ont montré que les effets du stress sur la cognition ne sont pas seulement influencés par la durée de l’exposition (aiguë vs chronique) et le niveau de stress, mais aussi par d’autres facteurs, tels que la contrôlabilité subjective ou objective du stress (Buetti & Lleras, 2012 ; Henderson, Snyder, Gupta & Banich, 2012 ; Mereu & Lleras, 2013). De nouvelles données montrent que la présence de la contrôlabilité peut améliorer les performances cognitives, tandis que dans des situations de stress incontrôlable, le stress peut avoir des effets néfastes sur le fonctionnement cognitif. Par exemple, l’étude d’Henderson et al. (2012) a démontré que le stress contrôlable perçu comme modérément intense est lié à l’amélioration des performances (évaluée par la réduction des interférences dans le test du Stroop), tandis que l’exposition à un stress incontrôlable ou une réponse extrême au stress nuit à la performance (Figure 5).
En matière neuronale, la présence de la contrôlabilité a été associée à l’implication du cortex préfrontal médian, lequel est associé à la présence du contrôle, inhibant très probablement les réponses au stress générées dans l’amygdale et conduisant à un comportement résilient (Kerr, McLaren, Mathy & Nitschke, 2012). À noter que la réduction de l’activité dans le cortex préfrontal médian a été également observée au cours d’une tâche stressante répétée chez des sujets ayant vécu des situations stressantes au début de leur vie (Wang, Paul, Stanton, Greeson & Smoski, 2013), et ceci pourrait être lié au ressenti d’incontrôlabilité chez ces individus.
Figure 5

L’interférence au test de Stroop en fonction de la contrôlabilité et le ressenti subjectif du stress : (a) Dans le groupe avec stress contrôlable (SC) – les niveaux de stress subjectivement perçus comme étant moyens sont liés à une meilleure performance (interférence réduite), alors que les niveaux de stress subjectivement perçus comme étant trop bas ou trop élevés sont liés à une performance détériorée ; (b) Dans le groupe avec stress incontrôlable (SI) – les niveaux de stress subjectif ne sont pas liés à la performance au Stroop.
En général, les données disponibles suggèrent qu’un niveau de stress optimal et contrôlable pourrait avoir des effets bénéfiques sur la cognition et le comportement, tandis que l’exposition répétée à des niveaux de stress extrêmes pourrait altérer la cognition et entraîner le développement de perturbations affectives, comme le trouble de stress post-traumatique. Du point de vue neuronal, la présence objective ou le sentiment subjectif de contrôle de la situation stressante implique l’engagement du cortex préfrontal qui joue un rôle important dans la régulation des réactions émotionnelles générées dans l’amygdale.
Conclusions et pistes de recherches futures
L’objectif primordial de la présente revue était d’examiner les résultats des études portant sur les effets opposés de l’émotion sur la cognition. La recherche dans ce domaine fournit la preuve que les effets d’amélioration et d’altération de l’émotion au sein du même processus cognitif, comme la mémoire épisodique, peuvent être attribués à différentes dissociations, comme la dissociation entre éléments centraux vs éléments périphériques, la dissociation en fonction de la priorité accordée et la dissociation entre encodages d’items isolés vs l’association complexe d’éléments. Les recherches actuellement menées suggèrent également que les effets opposés de l’émotion à travers des processus cognitifs différents pourraient être liés à la dissociation entre effet altérant immédiat et effet bénéfique à long terme. Ces effets engagent à la fois des mécanismes neuronaux communs et dissociables, impliquant des processus ascendants (bottom-up) et descendants (top-down).
Malgré le progrès important ayant été fait dans la compréhension des mécanismes sous-jacents des effets opposés de l’émotion, plusieurs questions doivent encore être clarifiées dans les recherches futures. Premièrement, les effets opposés de l’émotion sur la cognition ont été étudiés principalement dans un contexte de laboratoire, en utilisant des stimuli de laboratoire (par exemple, des images à charge émotionnelle), alors que l’impact adverse des émotions sur des événements personnels de la vie réelle (par exemple, les souvenirs autobiographiques émotionnels) reste peu étudié. Deuxièmement, alors que l’influence des ressources attentionnelles sur l’impact de l’émotion sur les processus perceptifs de bas niveau a été étudiée, on ne sait pas comment la manipulation de ressources attentionnelles dans les processus cognitifs de plus haut niveau influence l’impact de l’émotion. Troisièmement, les travaux de recherche étudiant les effets opposés de l’émotion doivent également considérer son implication dans le cas de troubles affectifs comme le trouble de stress post-traumatique, où ces effets sont exacerbés et dévastateurs. Il est tout à fait possible que le souvenir incontrôlable d’événements traumatiques puisse agir comme un distracteur interne puissant, pouvant dégrader la performance à la tâche en cours (Dolcos, 2013). La clarification de ces questions est d’une importance primordiale pour arriver à mieux comprendre les mécanismes qui sous-tendent les interactions entre émotion et cognition à la fois dans le fonctionnement sain et dans le cas des troubles émotionnels.
Parties annexes
Bibliographie
- Arnsten, A. F. (2000). Stress impairs prefrontal cortical function in rats and monkeys : Role of dopamine D1 and norepinephrine alpha-1 receptor mechanisms. Progress in Brain Research, 126, 183-192.
- Arnsten, A. F. (2009). Stress signalling pathways that impair prefrontal cortex structure and function. Nature Reviews Neuroscience, 10(6), 410-422.
- Buetti, S. & Lleras, A. (2012). Perceiving control over aversive and fearful events can alter how we experience those events : An investigation of time perception in spider-fearful individuals. Frontiers in Psychology, 3.
- Chiu, Y. C., Dolcos, F., Gonsalves, B. D. & Cohen, N. J. (2013). On opposing effects of emotion on contextual or relational memory. Frontiers in Psychology, 4, 103. doi : 10.3389/fpsyg.2013.00103
- Christianson, S. A. (1992). Emotional stress and eyewitness memory : A critical review. Psychological Bulletin, 112(2), 284-309.
- Cohen, N. J., Ryan, J., Hunt, C., Romine, L., Wszalek, T. & Nash, C. (1999). Hippocampal system and declarative (relational) memory : Summarizing the data from functional neuroimaging studies. Hippocampus, 9(1), 83-98.
- Corbetta, M. & Shulman, G. L. (2002). Control of goal-directed and stimulus-driven attention in the brain. Nature Reviews Neuroscience, 3(3), 201-215.
- Davachi, L., Mitchell, J. P. & Wagner, A. D. (2003). Multiple routes to memory : Dis-tinct medial temporal lobe processes build item and source memories. Proceedings of the National Academy of Sciences of the United States of America, 100(4), 2157-2162.
- Diamond, D. M., Campbell, A. M., Park, C. R., Halonen, J. & Zoladz, P. R. (2007). The temporal dynamics model of emotional memory processing : A synthesis on the neurobiological basis of stress-induced amnesia, flashbulb and traumatic memories, and the Yerkes-Dodson law. Neural Plasticity, 33, 60803.
- Dillon, D. G., Ritchey, M., Johnson, B. D. & LaBar, K. S. (2007). Dissociable effects of conscious emotion regulation strategies on explicit and implicit memory. Emotion, 7(2), 354-365.
- Dolcos, F. (2013). Linking enhancing and impairing effects of emotion – the case of PTSD. Frontiers in Integrative Neuroscience, 7, 26.
- Dolcos, F. & Denkova, E. (2008). Neural correlates of encoding emotional memories : A review of functional neuroimaging evidence. Cell Science Reviews, 5(2), 78-122.
- Dolcos, F., Denkova, E. & Dolcos, S. (2012). Neural correlates of emotional memories : A review of evidence from brain imaging studies. Psychologia, 55, 80-111.
- Dolcos, F., Diaz-Granados, P., Wang, L. & McCarthy, G. (2008). Opposing influences of emotional and non-emotional distracters upon sustained prefrontal cortex activity during a delayed-response working memory task. Neuropsychologia, 46(1), 326-335.
- Dolcos, F., Iordan, A., Kragel, J., Stokes, J., Campbell, R., McCarthy, G. & Cabeza, R. (2013). Neural correlates of opposing effects of emotional distraction on working memory and episodic memory : An event related fMRI investigation. Frontiers in Psychology, 4.
- Dolcos, F., LaBar, K. S. & Cabeza, R. (2006). The memory-enhancing effect of emotion : Functional neuroimaging evidence. In B. Uttl, N. Ohta & A. L. Siegenthaler (Eds.), Memory and emotion : Interdisciplinary perspectives (pp. 107-133). Malden, MA : Blackwell Publishing.
- Dolcos, F. & McCarthy, G. (2006). Brain systems mediating cognitive interference by emotional distraction. Journal of Neuroscience, 26(7), 2072-2079.
- Dolcos, F., Miller, B., Kragel, P., Jha, A. & McCarthy, G. (2007). Regional brain differences in the effect of distraction during the delay interval of a working memory task. Brain Research, 1152, 171-181.
- Dougal, S., Phelps, E. A. & Davachi, L. (2007). The role of medial temporal lobe in item recognition and source recollection of emotional stimuli. Cognitive, Affective, & Behavioral Neuroscience, 7(3), 233-242.
- Eichenbaum, H. & Cohen, N.J. (2001). From conditioning to conscious recollection : Memory systems of the brain. New York, NY : Oxford University Press.
- Ezzyat, Y. & Davachi, L. (2014). Similarity breeds proximity : Pattern similarity within and across contexts is related to later mnemonic judgments of temporal proximity. Neuron, 81(5), 1179-1189.
- Glascher, J. & Adolphs, R. (2003). Processing of the arousal of subliminal and supraliminal emotional stimuli by the human amygdala. Journal of Neuroscience, 23(32), 10274-10282.
- Graf, P. & Schacter, D. L. (1989). Unitization and grouping mediate dissociations in memory for new associations. Journal of Experimental Psychology Learning, 15, 930-940.
- Hains, A. B. & Arnsten, A. F. (2008). Molecular mechanisms of stress-induced pre-frontal cortical impairment : Implications for mental illness. Learning & Memory, 15(8), 551-564.
- Henckens, M. J., van Wingen, G. A., Joëls, M. & Fernández, G. (2012). Time-dependent effects of cortisol on selective attention and emotional interference : a functional MRI study. Frontiers in Integrative Neuroscience, 6, 66. doi : 10.3389/fnint.2012.00066
- Henderson, R. K., Snyder, H. R., Gupta, T. & Banich, M. T. (2012). When does stress help or harm ? The effects of stress controllability and subjective stress response on Stroop performance. Frontiers in Psychology, 3, 179.
- Joels, M. & Baram, T. Z. (2009). The neuro-symphony of stress. Nature ReviewsNeuroscience, 10(6), 459-466.
- Keightley, M. L., Winocur, G., Graham, S. J., Mayberg, H. S., Hevenor, S. J. & Grady, C. L. (2003). An fMRI study investigating cognitive modulation of brain regions associated with emotional processing of visual stimuli. Neuropsychologia, 41(5), 585-596.
- Kensinger, E. A. (2009). Remembering the details : Effects of emotion. EmotionReview, 1(2), 99-113.
- Kensinger, E. A., Garoff-Eaton, R. J. & Schacter, D. L. (2007a). Effects of emotion on memory specificity : Memory trade-offs elicited by negative visually arousing stimuli. Journal of Memory and Language, 56, 575-591.
- Kensinger, E. A., Garoff-Eaton, R. J. & Schacter, D. L. (2007b). How negative emotion enhances the visual specificity of a memory. Journal of Cognitive Neuroscience, 19(11), 1872-1887.
- Kensinger, E. A. & Schacter, D. L. (2006). Amygdala activity is associated with the successful encoding of item, but not source, information for positive and negative stimuli. Journal ofNeuroscience, 26(9), 2564-2570.
- Kerr, D. L., McLaren, D. G., Mathy, R. M. & Nitschke, J. B. (2012). Controllability modulates the anticipatory response in the human ventromedial prefrontal cortex. Frontiers in Psychology, 3, 557.
- Lee, T. H., Itti, L. & Mather, M. (2012). Evidence for arousal-biased competition in perceptual learning. Frontiers in Psychology, 3, 241.
- Lee, T. H., Sakaki, M., Cheng, R., Velasco, R. & Mather, M. (2014). Emotional arousal amplifies the effects of biased competition in the brain. Social Cognitive & Affective Neuroscience.
- Levine, L. J. & Edelstein, R. S. (2009). Emotion and memory narrowing : A review and goal-relevance approach. Cognition & Emotion, 23(5), 833-875.
- Loftus, E. F., Loftus, G. R. & Messo, J. (1987). Some facts about “weapon focus”. Law and Human Behavior, 11, 55-62.
- Luck, D., Leclerc, M. E. & Lepage, M (2014). The Potentiation of Associative Memory by Emotions : An Event-Related FMRI Study. Advances in Neuroscience.
- Lupien, S. J., Maheu, F., Tu, M., Fiocco, A. & Schramek, T. E. (2007). The effects of stress and stress hormones on human cognition : Implications for the field of brain and cognition. Brain and Cognition, 65(3), 209-237.
- Lupien, S. J., McEwen, B. S., Gunnar, M. R. & Heim, C. (2009). Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nature Reviews Neuroscience, 10(6), 434-445.
- Mather, M. (2007). Emotional arousal and memory binding : An object-based frame-work. Perspectives on Psychological Science, 2, 33-52.
- Mather, M. & Sutherland, M. R. (2011). Arousal-biased competition in perception and memory. Perspectives on Psychological Science, 6(2), 114-133.
- McEwen, B. S.(2007). Physiology and neurobiology of stress and adaptation : Central role of the brain. Physiological Reviews, 87(3), 873-904.
- McGaugh, J. L. (2000). Memory-a century of consolidation. Science, 287(5451), 248-251.
- McGaugh, J. L. (2004). The amygdala modulates the consolidation of memories of emotionally arousing experiences. Annual Review of Neuroscience, 27, 1-28.
- Mereu, S. & Lleras, A. (2013). Feelings of control restore distorted time perception of emotionally charged events. Consciousness and Cognition, 22(1), 306-314.
- Nadel, L. & Jacobs, W. J. (1998). Traumatic memory is special. Current Directions in Psychological Science, 7, 154-157.
- Park, C. R., Campbell, A. M., Woodson, J. C., Smith, T. P., Fleshner, M. & Diamond, D. M. (2006). Permissive influence of stress in the expression of a U-shaped relationship between serum corticosterone levels and spatial memory errors in rats. Dose Response, 4(1), 55-74.
- Phelps, E. A., O’Connor, K. J., Gatenby, J. C., Gore, J. C., Grillon, C. & Davis, M. (2001). Activation of the left amygdala to a cognitive representation of fear. Nature Neuroscience, 4(4), 437-441.
- Ranganath, C., Cohen, M. X., Dam, C. & D’Esposito, M. (2004). Inferior temporal, prefrontal, and hippocampal contributions to visual working memory maintenance and associative memory retrieval. Journal of Neuroscience, 24(16), 3917-3925.
- Roozendaal, B., McEwen, B. S. & Chattarji, S. (2009). Stress, memory and the amygdala. Nature Reviews Neuroscience, 10(6), 423-433.
- Sakaki, M., Fryer, K. & Mather, M. (2014). Emotion strengthens high-priority memory traces but weakens low-priority memory traces. Psychological Science, 25(2), 387-395.
- Sakaki, M., Ycaza-Herrera, A. E. & Mather, M. (2014). Association learning for emotional harbinger cues : When do previous emotional associations impair and when do they facilitate subsequent learning of new associations ? Emotion, 14(1),115-129.
- Sander, D., Grandjean, D. & Scherer, K. R. (2005). A systems approach to appraisal mechanisms in emotion. Neural Networks, 18(4), 317-352.
- Sandi, C. & Pinelo-Nava, M. T. (2007). Stress and memory : Behavioral effects and neurobiological mechanisms. Neural Plasticity, 78970.
- Scheuerecker, J., Frodl, T., Koutsouleris, N., Zetzsche, T., Wiesmann, M., Kleemann, A. M. … Meisenzahl, E. M. (2007). Cerebral differences in explicit and implicit emotional processing – an fMRI study. Neuropsychobiology, 56(1), 32-39.
- Shafer, A. T. & Dolcos, F. (2012). Neural correlates of opposing effects of emotional distraction on perception and episodic memory : An event-related fMRI investigation. Frontiers in Integrative Neuroscience, 6, 70.
- Staresina, B. P. & Davachi, L. (2010). Object unitization and associative memory formation are supported by distinct brain regions. Journal of Neuroscience, 30(29), 9890-9897.
- Tubridy, S. & Davachi, L. (2011). Medial temporal lobe contributions to episodic sequence encoding. Cerebral Cortex, 21(2), 272-280.
- Wang, L., Paul, N., Stanton, S. J., Greeson, J. M. & Smoski, M. (2013). Loss of sustained activity in the ventromedial prefrontal cortex in response to repeated stress in individuals with early – life emotional abuse : Implications for depression vulnerability. Frontiers in Psychology, 4, 320.
- Waring, J. D., Payne, J. D., Schacter, D. L. & Kensinger, E.A. (2010). Impact of individual differences upon emotion-induced memory trade-offs. Cognition & Emotion, 24(1), 150-167.
Liste des figures
Figure 1
Taux de reconnaissance correcte pour l’item central et le contexte (l’arrière-plan) de l’image démontrant le compromis entre éléments centraux et éléments périphériques. Alors que les items négatifs ont été mieux rappelés que les items neutres, le contexte associé avec un item émotionnel a été moins bien rappelé que le contexte associé avec un item neutre.
Figure 2
L’effet de priorisation au niveau de la performance mnésique lors d’un test de mémoire où les participants doivent retenir des images d’objets en mémoire en portant leur attention soit sur un objet encadré, soit sur l’objet qui le précède. Comparées à des images neutres, les images à charge émotionnelle négative améliorent la mémoire pour l’image précédente neutre (Objet -1) quand celle-ci est privilégiée lors de l’encodage (Objet -1 prioritaire), et non pas quand elle n’est pas privilégiée (Objet prioritaire).
Figure 3
Dissociation entre effet immédiat et effet à long terme de la distraction émotionnelle : (a) Vue coronale de l’amygdale illustrant l’activation bilatérale liée à l’effet altérant immédiat (en gris foncé) et l’activation gauche liée à l’effet facilitant à long terme (en blanc) ; à noter que l’activation d’une sous-région de l’amygdale gauche est positivement corrélée avec la performance mnésique ; (b) Vue en 3-D du cerveau illustrant l’implication du cortex préfrontal médian dans l’effet altérant immédiat (en gris foncé) et l’implication du cortex pariétal dans l’effet facilitant à long terme (en blanc). D = hémisphère droit, G = hémisphère gauche.
Figure 4
Les activations cérébrales liées à la détérioration de la mémoire de travail (MT) et/ou l’amélioration de la mémoire épisodique (ME) par la distraction émotionnelle : (a) Désactivation plus prononcée dans le cortex préfrontal (CPF) dorso-latéral (en bleu) ; (b) Activation plus prononcée dans l’amygdale (AMY, en rouge) et l’hippocampe (HC, en vert) liée à une mémoire de travail détériorée et une mémoire épisodique améliorée ; (c) Activation plus prononcée dans le cortex préfrontal ventro-latéral (en rouge) liée à l’amélioration de la mémoire de travail et mémoire épisodique. Il est intéressant de noter que des sous-régions de l’amygdale et du cortex préfrontal ventro-latéral (en blanc) contribuent différemment à l’effet de la distraction émotionnelle sur la mémoire de travail, l’amygdale étant liée à une performance détériorée (corrélation négative) et le cortex préfrontal ventro-latéral étant lié à une performance améliorée (corrélation positive). D = hémisphère droit, G = hémisphère gauche.
Figure 5
L’interférence au test de Stroop en fonction de la contrôlabilité et le ressenti subjectif du stress : (a) Dans le groupe avec stress contrôlable (SC) – les niveaux de stress subjectivement perçus comme étant moyens sont liés à une meilleure performance (interférence réduite), alors que les niveaux de stress subjectivement perçus comme étant trop bas ou trop élevés sont liés à une performance détériorée ; (b) Dans le groupe avec stress incontrôlable (SI) – les niveaux de stress subjectif ne sont pas liés à la performance au Stroop.