Résumés
Résumé
Cette étude a pour but la caractérisation hydrobiologique d’une lagune méditerranéenne (lagune de Bizerte, nord de la Tunisie) sur la base de dix campagnes de mesures réalisées à une fréquence mensuelle durant l’année 2004, de janvier à décembre. Deux parmi ces dix campagnes ont fait l’objet d’un maillage fin avec 118 stations en mai et 40 stations en septembre. Le reste n’a couvert que 10 stations seulement. Pendant ces campagnes, des paramètres physicochimiques ont été mesurés (Température, Salinité, Oxygène dissous). Des mesures de paramètres chimiques et biologiques ont été effectuées au laboratoire (Ammonium, Nitrates, Phosphates, Azote total, Phosphore total, Chlorophylle a et Phéopigment).
Une analyse multivariée des données collectées a permis de distinguer deux principaux modes de fonctionnement de la lagune. Le premier mode, expliquant 26,3 % de la variance totale, montre un contraste nord-sud et reflète l’effet relatif de la Méditerranée et du bassin versant de la lagune, alors que le second mode, expliquant 18,2 % de la variance totale, montre un contraste est-ouest qui traduit l’effet du lac Ichkeul. Les paramètres qui contribuent significativement au premier mode sont la température, la salinité, la transparence absolue et relative. Ceux qui contribuent au second mode sont la transparence absolue et relative ainsi que la chlorophylle a et le phéopigment. Cependant, les paramètres dont les poids moyens sont non significatifs ont montré des oscillations spatiales importantes en fonction des saisons qui sont en liaison avec les rejets terrestres.
Mots-clés :
- Lagune de Bizerte,
- Hydrobiologie,
- Concentration des nutriments,
- Analyse multivariée
Abstract
The purpose of this study was the hydrobiological characterization of a Mediterranean lagoon (Bizerte Lagoon, North Tunisia) based on ten sampling campaigns carried out during the year 2004, from January to December, on a roughly monthly frequency. Two of these ten campaigns were subject to a high spatial coverage with 118 stations in May and 40 stations in September; the eight remaining campaigns covered only ten stations. During these campaigns, physicochemical parameters were measured (temperature, salinity, dissolved oxygen) and laboratory analyses were performed for chemical and biological parameters (ammonium, nitrate, phosphate, total nitrogen, total phosphorus, chlorophyll-a and pheopigments). A multivariate analysis distinguished two principal functioning hydrobiologic modes. The first mode, explaining 26.3% of the total variance, shows a North-South contrast reflecting the relative effect of the Mediterranean Sea and the catchment area. The second mode, explaining 18.2% of the total variance, shows an East-West contrast that reflects the effect of Lake Ichkeul. The parameters that contribute significantly to the first mode are temperature, salinity, absolute and relative transparency, whereas those that contribute to the second mode are absolute and relative transparency as well as chlorophyll-a and pheopigments. However, parameters for which the average weights were not significant showed important spatial oscillations according to seasons, in relation with terrestrial discharges.
Keywords:
- Bizerte Lagoon,
- Hydrobiology,
- Nutrient concentration,
- Multivariate analysis
Corps de l’article
1. Introduction
Les lagunes méditerranéennes sont des écosystèmes particulièrement sensibles à l'eutrophisation. Leurs conditions hydrobiologiques sont sous l’influence directe de la quantité et de la qualité d’effluents qui s’y déversent. L’aménagement de ces plans d’eau de haute valeur socio-économique nécessite des études hydrobiologiques approfondies afin de comprendre leur fonctionnement et d’estimer leur situation trophique.
Par ailleurs, les proliférations récurrentes de microalgues nuisibles indiquent une dégradation biologique de ces écosystèmes côtiers. À cet effet, au niveau des côtes méditerranéennes de l’Espagne, ce sont les proliférations d'Alexandrium taylori qui sont signalées de manière récursive et ont été attribuées au degré de confinement hydrodynamique des eaux plutôt qu'à un enrichissement en nutriments (GARCES et al., 1999). En Tunisie, les phénomènes d’eaux colorées associées à des proliférations d’algues nuisibles en milieux lagunaires (Dinophysis spp. et Prorocentrum lima), ont fait l’objet de plusieurs études reliant les facteurs climatiques et trophiques (BENRAJEB-JENHANI et ROMDHANE, 2002; DALY YAHIA et al., 2005; DALY YAHIA-KEFI et DALY YAHIA, 1997; TURKI et EL ABED, 1999).
La lagune de Bizerte, située au nord de la Tunisie, constitue un réceptacle qui draine toute l’eau du bassin versant en amont. Les apparitions d’efflorescences phytoplanctoniques, parfois toxiques, y sont devenues de plus en plus régulières en réponse aux multiples rejets qui s’y déversent (MAERH, 2003; SAKKA HLAILI et al., 2006). Plusieurs études lui ont été consacrées depuis les années 1950 (AZOUZ, 1966; CHAUMONT, 1956; FRISONI et al., 1986; GIMAZANE, 1981; ZAOUALI, 1974 ; ZAOUALI, 1979). Toutefois, ce n’est qu’à partir des années 1980 que ce milieu lagunaire a fait l’objet de travaux de recherche pluridisciplinaire visant à définir son fonctionnement hydrologique, sédimentologique et biologique (BELKHIR et al., 1985; SOUSSI, 1983).
Ainsi, l’objectif de la présente étude est d’étudier la variabilité spatio-temporelle des paramètres hydrobiologiques dans la lagune de Bizerte et de comprendre, par l’application d’une méthode objective, son mode de fonctionnement hydrobiologique en se basant sur les mesures mensuelles effectuées durant l’année 2004.
Notons que les modes de variabilité des paramètres hydrobiologiques dans cette lagune ont fait l’objet d’une étude antérieure (Béjaouiet al., 2008) dont l’objectif était de valider une nouvelle variante de l’analyse EOF (Empirical Orthogonal Function) sur les mesures hydrobiologiques réalisées en 2004. Dans cette approche, l’information concernant la variation spatio-temporelle des paramètres dans le système SI était manquante du fait de la normalisation des paramètres. La présente étude complète cette dernière en détaillant la variation spatio-temporelle des paramètres (variation saisonnière des paramètres hydrobiologiques avec leurs unités et modes de variabilité saisonnière).
2. Présentation de la lagune de bizerte
La lagune de Bizerte est située au nord de la Tunisie (latitude : 37°8’-37°14’ N, longitude : 9°46’-9°56’ E). Elle s’étend sur une superficie de 128 km2 (la largeur maximale est de 11 km et la longueur maximale est de 13 km) avec une profondeur moyenne de 7 m. Elle communique avec la mer par un goulet de 6 km de longueur et 12 m de profondeur. La lagune draine un bassin versant de l’ordre de 380 km2. Elle est reliée dans sa partie ouest à un second plan d’eau, le lac Ichkeul, par l’intermédiaire du canal Tinja d’environ 5 km de long et de quelques mètres de profondeur (3 mètres en période de crue) (Figure 1). Le canal est actuellement équipé d’écluses permettant de gérer les échanges entre le lac Ichkeul et la lagune de Bizerte. Avant la construction de ces ouvrages, les apports à travers le canal Tinja vers la lagune de Bizerte ont été estimés pour une année normale à 165 Mm3•an-1 (ANPE, 2007). Après la mise en service des barrages (Joumine, Ghezala et Sejnene) et l’installation des écluses sur le canal Tinja en 1989, les apports d’eau du lac Ichkeul dans la lagune ont diminué.
Figure 1
Situation géographique et délimitation du bassin versant de la lagune de Bizerte.
Location and delimitation of the catchment area of the Bizerte Lagoon.

Des villes en expansion urbaine et industrielle (Bizerte, Menzel Jemil, Menzel Abderrahmen et Menzel Bourguiba) jonchent le pourtour de la lagune (Figure 2).
Figure 2
Carte bathymétrique de la lagune de Bizerte et répartition des stations de prélèvement.
Bathymetric map of the Bizerte Lagoon and distribution of the sampling stations.
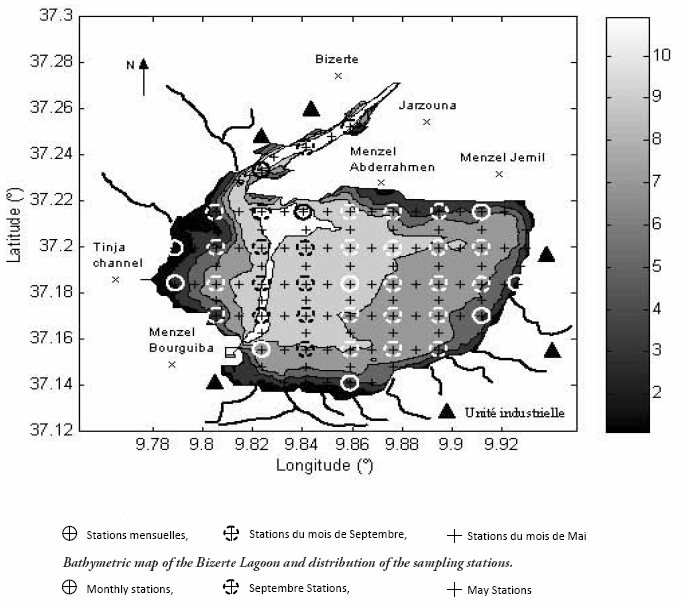
L’appartenance de cette lagune à une région d’effondrement entourée d’une série de petites montagnes lui confère le caractère d’un réceptacle d’un réseau hydrographique développé composé de plusieurs cours d’eau qui débouchent principalement aux niveaux des bordures sud et ouest de la lagune et qui charrient une quantité d’eau variable en fonction des précipitations et des apports provenant du lac Ichkeul.
À cette double influence marine et continentale, s’ajoute une troisième influence d’ordres urbain et industriel (MAERH, 2003). Néanmoins, deux principales stations d’épuration d’eaux usées (STEPs de Menzel Bourguiba et de Bizerte) ont été récemment installées autour de la lagune. Depuis la mise en service des stations d’épuration des villes de Bizerte (en octobre 1997) et de Menzel Bourguiba (en janvier 1998), les eaux usées rejetées dans la lagune sont en majorité épurées (MAERH, 2003). D’autre part, la lagune est exploitée par une activité conchylicole au niveau de cinq secteurs.
3. Cadre climatique
Le climat de la région de Bizerte est de type méditerranéen à nuance sub-humide où l’été est chaud et sec et l’hiver est doux et pluvieux (KALLEL, 1989).
Les vents soufflent généralement du secteur nord-ouest. Les vents d’ouest sont les plus fréquents et soufflent à des vitesses comprises entre 5 et 9 m•s-1 pouvant atteindre 15 m•s-1 (HIDROTECNICA PORTUGUESA, 1995). La température de l’air dans la région de Bizerte varie entre 10 °C au mois de janvier et 27 °C au mois d’août avec une moyenne annuelle de 19 °C. La précipitation moyenne annuelle est de l’ordre de 74 Mm3•an-1 (ANPE, 1990). Environ 65 % de cette quantité a été enregistrée pendant la saison hivernale (ANPE, 1990). Les fortes évaporations ont été enregistrées aux mois de juillet et août alors que les faibles évaporations ont été enregistrées aux mois de janvier et décembre. Le bilan précipitation-évaporation est estimé à ‑83 Mm3•an‑1 en moyenne (ANPE, 1990).
4. Materiel et methodes
4.1 Analyse des échantillons
Dix campagnes mensuelles de prélèvement d’eau de surface ont été effectuées au cours de l’année 2004. Deux parmi celles-ci (mai et septembre) ont fait l’objet de prélèvements à maillage fin (118 stations au mois de mai et 40 stations au mois de septembre) alors que les autres ont porté sur 10 stations seulement couvrant toute son étendue (Figure 2). Les échantillons d’eau ont fait l’objet de mesures in situ (température, salinité, concentration en oxygène dissous et transparence) et d’analyses en laboratoire (matières en suspension, nutriments, azote total, phosphore total, chlorophylle a et phéopigments).
La température de l’eau (T) a été mesurée à environ 10 cm de la surface à l'aide d'un thermomètre électronique. La salinité de l’eau (S) et la teneur en oxygène dissous (O2) ont été mesurées, respectivement, à l’aide d’un conductimètre Thermo Orion de type 115Aplus et d'un oxymètre Orion de type 810Aplus préalablement étalonnés. La transparence de l'eau (Tr) a été évaluée à l'aide du disque de Secchi (HOLMES, 1970). La transparence relative (Tr%) est définie comme la transparence de Secchi rapportée à la profondeur d’eau.
Au laboratoire, les dosages des nitrites (NO2-), nitrates (NO3-), ammonium (NH4+) et phosphates (PO43-) ont été réalisés à l’aide d’un auto-analyseur (TREGUER et LE CORRE, 1975). L’azote total (NT) et le phosphore total (PT) ont été dosés après oxydation des composés organiques. Le dosage des pigments chlorophylliens (Chlorophylle a et phéopigments) a été réalisé par spectrophotométrie (RODIER et al., 1996). La concentration des matières en suspension (MES) a été déterminée par double pesée avant et après passage à l’étuve à 105 °C du filtrat solide. Le filtrat a été recueilli par filtration d’un litre d’eau avec des filtres millipores 0,45 µm.
4.2 Méthode d’analyse multivariée
Les données hydrobiologiques ont été interpolées linéairement sur une grille curviligne orthogonale (52 x 61 mailles) en utilisant les distances comme facteurs de pondération. La dimension de la grille selon la longitude (∆x) varie entre 75 et 495 m alors que la dimension selon la longitude (∆y) varie entre 218 et 686 m. La concentration moyenne de chaque paramètre sur l’ensemble du plan d’eau a été ensuite calculée pour chaque campagne mensuelle. Une matrice de données brutes xb (1 783 x 111) a été construite en remplissant les colonnes par les paramètres mesurés pendant toutes les campagnes. Ainsi, chaque colonne de cette matrice représente un paramètre mesuré (111 paramètres pour toutes les campagnes) et chaque ligne représente un point de grille (1 783 points de grille). Certains paramètres tels que la température et la salinité sont représentés dix fois dans la matrice car mesurés à chaque campagne alors que d’autres sont représentés en fonction des mesures réalisées des paramètres pendant les dix campagnes : l’oxygène dissous est représenté cinq fois (mesuré seulement cinq fois pendant les dix campagnes) et la Chl a six fois (mesurées six fois pendant les dix campagnes).
Les données de cette matrice ont été ensuite standardisées par rapport aux variations spatiales. Cette technique a permis de déterminer les anomalies et de définir la structure moyenne permanente de l’ensemble des paramètres. Une analyse multivariée a été ensuite appliquée sur les données hydrobiologiques transformées (ESCOFIER et PAGES, 1990; SCHRUM et al., 2006).
La matrice de données centrées réduites xs est donnée par :
où i représente la ie station et j le je paramètre. ![]() représente la moyenne spatiale du je paramètre et
représente la moyenne spatiale du je paramètre et ![]() l’écart-type spatial relatif. La matrice des corrélations Cs (111 x 111) est ensuite construite. Les valeurs propres λsk et les vecteurs propres vsjk de cette matrice sont alors calculés. L’indice k représente le ke mode relatif à la ke valeur propre et au ke vecteur propre. La projection de la matrice xs sur les vecteurs propres relatifs donne la matrice des composantes principales ys.
l’écart-type spatial relatif. La matrice des corrélations Cs (111 x 111) est ensuite construite. Les valeurs propres λsk et les vecteurs propres vsjk de cette matrice sont alors calculés. L’indice k représente le ke mode relatif à la ke valeur propre et au ke vecteur propre. La projection de la matrice xs sur les vecteurs propres relatifs donne la matrice des composantes principales ys.
La part la plus importante de l’information contenue dans les données se trouve alors sur l’axe factoriel associé à la plus grande valeur propre.
5. Résultats
Les variations saisonnières des concentrations moyennes des paramètres hydrobiologiques sont présentées sous la forme d’histogrammes sur les figures 3 et 4. Ces moyennes sont réalisées sur les données brutes. Les écarts-types indiquant l’importance de la variabilité spatiale des paramètres sont aussi représentés sur les mêmes figures.
Figure 3
Variation mensuelle des paramètres physico-chimiques : température (A), salinité (B), oxygène dissous (C), matières en suspension (D) et la transparence (E). Les barres indiquent l’importance du gradient spatial du paramètre.
Monthly variation of physico-chemical parameters: temperature (A), salinity (B), dissolved oxygen (C), suspended matter (D) and transparency (E). The bars indicate the importance of the spatial gradient of the parameter.

Figure 4
Variation mensuelle de la concentration de l’ammonium (A), nitrates (B), phosphates (C), azote total (D), phosphore total (E) et de la chlorophylle a (F). Les barres indiquent l’importance du gradient spatial du paramètre.
Monthly variation of the concentration of ammonia (A), nitrate (B), phosphate (C), total nitrogen (D), total phosphorus (E) and chlorophyll-a (F). The bars indicate the importance of the spatial gradient of the parameter.

Afin de tester l’importance de la variabilité saisonnière, une analyse de la variance (ANOVA) à un facteur a été réalisée. Les paramètres ont été pris un à un puisque le nombre de stations est variable en fonction des mois.
Pour compléter cette analyse de la variance, le test de comparaison multiple des moyennes de Scheffé a été appliqué. Ce test a permis de tester si les moyennes mensuelles des paramètres (variables indépendantes) sont significatives.
Les résultats de cette analyse (ANOVA et test de Scheffé) sont présentés dans le tableau 1.
Tableau 1
Résultats du test ANOVA à un facteur (Scheffé F test) appliqué aux variables hydrobiologiques. Le test a été appliqué aux variables une à une, après une analyse de la variance (ANOVA). Une cellule grise indique que la différence (à un seuil de 5 %) entre les moyennes des variables est significative. Une cellule blanche indique que la différence n’est pas significative. Par exemple, la différence au niveau de 5 % des moyennes mensuelles de la température est significative sauf pour les mois de (février – décembre) et (juillet –septembre). La transparence n’est pas présentée dans ce tableau parce que les moyennes mensuelles sont significatives pour tous les mois.
Results of one factor ANOVA (Scheffé F test) applied to hydrobiological variables. The test was applied to variables taken one by one, after an analysis of variance (ANOVA). A grey cell indicates that the difference (in a 5% threshold) between the averages of variables is significant. A white cell indicates that the difference is not significant. For example, the difference at the level of 5% of the monthly averages of the temperature is significant except for (February-December) and (July-September) months. The transparency is not presented in this tableau because the monthly means are significant for all months.

5.1 Les paramètres physico-chimiques
La température moyenne de l’eau montre une variation saisonnière avec un maximum au mois d’août (29 °C) et un minimum au mois de janvier (11 °C) (Figure 3A).
Il en va de même pour la salinité moyenne qui montre une évolution sinusoïdale bien nette avec de fortes valeurs en été (35,8 psu) et de faibles valeurs en hiver (33,9 psu) (Figure 3B).
À l’opposé, l’oxygène dissous (Figure 3C) montre des concentrations moyennes élevées en hiver (8 mg•L-1) et faibles en été (au mois de juillet) (4 mg•L-1).
La variation saisonnière de la concentration des matières en suspension montre deux périodes contrastées : une période hivernale où les eaux sont chargées en MES avec une concentration maximale égale à 35 mg•L-1 et une période estivale où les eaux sont peu chargées en MES avec des concentrations inférieures à 20 mg•L-1 (Figure 3D).
La variation saisonnière de la transparence montre que les valeurs les plus élevées ont été généralement observées pendant le printemps et l’été. Les valeurs les plus faibles observées en novembre, décembre et janvier résultent d’une forte turbidité pendant la saison hivernale. Le gradient spatial (écarts-types élevés) observé entre les différentes stations est principalement dû à la vitesse du courant et à la bathymétrie. Pour la plupart des mesures, une corrélation négative entre la transparence et la teneur en matière en suspension en surface a été observée (Figure 3D).
5.2 Les paramètres chimiques et biologiques
La concentration de l’ammonium à l’échelle de la lagune de Bizerte montre des variations saisonnières importantes comprises entre 0,7 µmol•L-1 (au mois de mai) et 2,3 µmol•L-1 (au mois de septembre) (Figure 4A). Les teneurs en nitrates augmentent en hiver (2,3 µmol•L-1) et diminuent en été (0,8 µmol•L-1) (Figure 4B). Quant aux phosphates, leurs variations saisonnières semblent être inverses par rapport à celles de l’ammonium et des nitrates, avec des concentrations faibles au mois de septembre (0,1 µmol•L-1) et fortes au mois d’août (0,4 µmol•L-1) (Figure 4C). L’augmentation de la concentration en phosphates observée au mois d’août peut être expliquée par un relargage de cet élément à partir des sédiments du fond en présence de conditions réductrices (HASNAOUI et al., 2001, IFREMER, 2004) et/ou par une augmentation de la concentration de cet élément dans les eaux usées avec l’élévation de la température estivale (NARASIAH et al., 1988). La lagune de Bizerte est caractérisée par de faibles concentrations en azote total (10,7 ‑ 14,3 µmol•L‑1) (Figure 4D) et en phosphore total (1,1 ‑ 1,8 µmol•L‑1). Les variations saisonnières de la concentration en phosphore total présentent deux épisodes différents (Figure 4E) : une période hivernale caractérisée par des teneurs élevées (1,6 ‑ 1,9 µmol•L‑1) et une période estivale caractérisée par des teneurs relativement faibles (1,1 ‑ 1,3 µmol•L‑1).
Les variations saisonnières de la concentration moyenne de la chlorophylle a montre deux pics : le premier apparaît au mois d’avril (5,2 µg•L‑1) et le deuxième au mois d’août (6,5 µg•L-1) (Figure 4F). Au moment du premier pic, la distribution spatiale de la chlorophylle a (Figure 5A) montre que les concentrations les plus élevées (5,2 - 6,5 µg•L-1) se trouvent dans les eaux du goulet ainsi que les berges de la lagune qui sont caractérisées par une abondance en nutriments. Les faibles concentrations ont été observées dans la zone qui s’étend du secteur sud-ouest (en face de Menzel Bourguiba) allant vers le centre de la lagune. La distribution spatiale des phéopigments à la même période (Figure 5B) est quasiment l’inverse de celle de la chlorophylle a. Au moment du deuxième pic, les distributions spatiales de la biomasse chlorophyllienne (Figure 5C) et des phéopigments (Figure 5D) sont similaires à celles observées au printemps mais les concentrations sont plus élevées pour la chlorophylle a et sont du même ordre de grandeur pour les phéopigments. Le goulet présente des concentrations en chlorophylle a de l’ordre de 8,5 µg•L‑1 alors que le secteur sud-ouest, en face de la zone industrielle de Menzel Bourguiba, présente des concentrations plus faibles. Au niveau de la région nord-est de la lagune (zone conchylicole de Menzel Jemil), les concentrations en chlorophylle a sont relativement élevées (8 µg•L-1).
Figure 5
Distributions spatiales des concentrations en chlorophylle a et en phéopigments en avril (A et B) et en soût (C et D).
Spatial distributions of chlorophyll-a and pheopigment concentrations in April (A and B) and in August (C and D).

5.3 Analyse multivariée
La décomposition des données en modes orthogonaux a montré que les deux premiers modes expliquent la part la plus importante de la variance totale avec 26,3 % et 18,2 %, respectivement. Le troisième mode explique 13,6 % seulement de la variance totale. De ce fait, seuls les deux premiers modes sont retenus. L’ensemble des deux premiers modes représente 45 % de la variance totale. Il reste 55 % de la variance totale répartis sur 109 modes (111-2), soit approximativement 0,5 % de la variance totale. Ce pourcentage est clairement inférieur à ceux des deux premiers modes. D’autre part, la représentation graphique du troisième et du quatrième modes présente des structures très hétérogènes qui ne permettent pas d’extraire des informations précises.
Les résultats de la décomposition en modes orthogonaux sont présentés sous forme de séries temporelles et de distributions spatiales. Le poids moyen de chaque paramètre est indiqué par une ligne horizontale. Un test de Student au seuil de 5 % a été appliqué pour vérifier l’hypothèse de la moyenne nulle. Les poids des paramètres qui sont significativement différents de zéro sont indiqués par un carré noir, ceux des autres par un carré blanc.
Les poids mensuels des paramètres hydrobiologiques ainsi que la distribution spatiale du premier mode sont présentés respectivement sur les figures 6A et B. Les paramètres montrant des poids moyens différents de zéro sont T, S, Tr et Tr%. Les signes de S et de Tr sont opposés à ceux de T et de Tr%. La distribution spatiale de ce mode montre une opposition nord-sud qui met en évidence la contribution des zones sous influence marine et celles au sud sous influence continentale. La température de l’eau est relativement élevée dans les régions peu profondes du sud et sud-est de la lagune. A l’opposé, le centre et le nord-ouest de la lagune montrent des eaux plus fraîches et transparentes reflétant les caractéristiques des eaux marines. L’analyse des poids mensuels (Figure 6A) ainsi que la distribution spatiale permanente des poids de la salinité (Figure 6B) montrent que les faibles salinités sont observées au sud de la lagune alors que celles observées au nord sont plus élevées, reflétant l’effet de la Méditerranée. La transparence relative élevée au sud et faible au centre et au nord-est de la lagune est attribuée à l'effet de la profondeur de la colonne d’eau. La contribution des paramètres chimiques et biologiques n’est pas significative dans ce premier mode.
Figure 6
Variation temporelle (A) et distribution spatiale (B) des poids des paramètres hydro-biologiques du premier mode. L’axe des abscisse de la carte (A) représente les paramètres : Température (T), Salinité (S), Transparence (Tr), Matières en suspension (MES), Transparence relative (Tr%), Oxygène (O2), Nitrites (NO2), Nitrates (NO3), Ammonium (NH4), Azote total (NT), Phosphates (PO4), Phosphore (PT), Chlorophylle a (Chl a) et Phéopigments (Pheo). L’axe des ordonnées représente les poids des paramètres. Les poids mensuels de la température (T) et de la salinité (S) sont de même signe alors que les signes des poids mensuels des autres paramètres sont variables en fonction des mois. Ceci montre que la variabilité spatiale des paramètres est due entre autres aux effets oscillants de la Méditerranée et du bassin versant. Par exemple, les poids mensuels (carte B) de la température sont tous positifs, indiquant que la température des eaux au sud de la lagune est souvent (pour tous les mois) supérieure à celle au nord conséquence des rejets chaudes au sud de la lagune.
Series weights (A) and loadings (B) of hydrobiological parameters of the first mode. The x-axis of the chart (A) represents the parameters: Temperature (T), Salinity (S), Transparency (Tr), Suspended matter (MES), Relative transparency (Tr%), Oxygen (O2), Nitrite (NO2), Nitrate (NO3), Ammonia (NH4), Total nitrogen (NT), Phosphorus (PO4), Total phosphorus (PT), Chlorophyll-a (Chl-a) and Pheopigments (Pheo). The y-axis represents the parameter weights. The monthly weights of the temperature (T) and salinity (S) are of the same sign whereas the signs of the monthly weights of the other parameters are variable according to months. This shows that the spatial variability of the parameters is due to the oscillating effects of the Mediterranean and the catchment area. For example, the monthly weights (chart B) of the temperature are all positive indicating that the water temperature in the south of the lagoon is often higher than that of the north, a result of the thermal discharges in the south of the lagoon.

La description présentée plus haut correspond aux paramètres dont les poids moyens sont significativement différents de zéro selon le test de Student (BEJAOUI et al., 2008). Cependant, l’amplitude et le signe des poids temporels des autres paramètres varient considérablement en fonction des saisons, ils méritent par conséquent d’être étudiés.
Pour le cas de la MES, et si nous admettons qu’un poids mensuel est significatif s’il est supérieur à un seuil de 0,5, nous pouvons alors en déduire que le contraste est plus prononcé au mois de mai (poids = -1,3) (Figure 6A) avec des concentrations importantes au sud de la lagune en comparaison avec le secteur nord. Cette structure est encore valable pour le mois de décembre mais le contraste nord-sud est moins important. Notons que ce contraste s’inverse pour les mois d’avril et juillet où la concentration en MES devient plus importante au nord qu’au sud. Cette inversion du sens du contraste est due aux apports des rivières qui se déversent en majorité dans le secteur sud. Les eaux montrent une oxygénation plus élevée au sud qu’au nord. En hiver, le sens du contraste s’inverse et les eaux deviennent plus oxygénées au nord en liaison avec les échanges avec la Méditerranée. Pour les nitrates et l’ammonium, le contraste nord-sud le plus prononcé est observé au mois de juillet avec des teneurs élevées au sud. En ce qui concerne l’azote total, ses teneurs sont toujours plus élevées au sud alors que les teneurs en nitrites sont plus élevées au nord. Le contraste des phosphates est plus important aux mois de mai, août et février. Pour le phosphore total, les contrastes les plus importants sont observés aux mois de novembre et décembre mais avec des orientations différentes. Pour la Chl a, nous observons un changement du sens du contraste alors que le phéopigment garde presque des concentrations plus élevées au sud de la lagune, en liaison avec les conditions défavorables qui y règnent, en particulier le secteur sud-ouest. Le changement des structures des paramètres, en particulier ceux chimiques, est lié, d’une part, aux rejets urbains et industriels qui ont un caractère intermittent au cours du temps au point de vue quantité et qualité et, d’autre part, à l’échange d’eau entre la lagune et la Méditerranée. Les contrastes nord-sud de la température, la salinité, la transparence et la transparence relative gardent la même orientation au cours du temps, leur intensité est fonction des apports des rivières et des échanges avec la Méditerranée (BEJAOUI et al., 2008).
Les poids mensuels des paramètres hydrobiologiques ainsi que la distribution spatiale du deuxième mode sont présentés respectivement sur les figures 7A et B. La transparence (Tr) et la transparence relative (Tr%) mais également la chlorophylle a (Chl a) et les phéopigments (Pheo) ont des valeurs moyennes sensiblement différentes de zéro. Ce mode montre l'effet local du canal Tinja à l’ouest de la lagune en comparaison avec la partie centrale et la région sud-ouest en face de la zone industrielle de Menzel Bourguiba. Au niveau du canal Tinja, la Tr et le Pheo sont faibles alors que Tr% et Chl a sont élevées. Le secteur sud-ouest est caractérisé par des eaux transparentes, des concentrations élevées en Pheo et faibles en Chl a.
Figure 7
Variation temporelle (A) et distribution spatiale (B) des poids des paramètres hydro-biologiques du deuxième mode. Pour chaque paramètre, les poids mensuels (+) et moyen (carré) sont présentés.
Series weights (A) and loadings (B) of hydrobiological parameters of the second mode. For each parameter monthly weight (+) and mean (square) are presented.

Les poids mensuels du deuxième mode montrent des oscillations est-ouest relativement importantes. La température au centre de la lagune, en particulier le secteur sud-est, est généralement supérieure à la température observée à l’embouchure du canal Tinja. Cependant, en plein hiver, la situation s’inverse. Au niveau de ces secteurs (centre et le secteur sud-ouest), la salinité en période estivale est supérieure à celle observée à l’embouchure du canal Tinja et le goulet. La concentration en matières en suspension présente des oscillations mensuelles est-ouest importantes. En hiver, sa concentration à l’embouchure du canal Tinja est nettement supérieure à celles du reste de la lagune. Ces concentrations élevées sont dues aux apports du canal Tinja chargés en matières en suspension. La transparence au niveau du canal Tinja est faible en comparaison au reste de la lagune alors que la transparence relative exhibe des oscillations inverses. L’oxygène dissous présente une oscillation est-ouest remarquable en saisons hivernale et printanière. En ce qui concerne les nitrates, le contraste le plus prononcé est observé au mois de janvier où les teneurs sont élevées au centre et le secteur sud-est de la lagune et faible à l’embouchure du canal Tinja. L’ammonium, l’azote total, les phosphates et le phosphore total présentent des contrastes mensuels est-ouest variables en fonction des apports urbains et industriels. Pour la concentration en Chl a, on observe aussi des oscillations est-ouest variables en fonction des mois mais avec la même orientation. Les concentrations à l’est et au niveau du secteur sud-ouest sont toujours plus élevées que celles à l’embouchure du canal Tinja. Les oscillations du phéopigment sont quasiment l’inverse.
6. Discussion
L’étude hydrobiologique entreprise pendant un cycle annuel dans la lagune de Bizerte a permis de définir les grands traits de son fonctionnement hydrobiologique. Cette étude a aussi permis de mettre en évidence l’effet important de la Méditerranée et du canal Tinja sur la variabilité saisonnière des paramètres hydrobiologiques. En effet, avant la construction des barrages sur les principaux cours d’eau qui alimentent l’Ichkeul (les barrages de Joumine en 1983, Ghezala en 1984 et Sejnane en 1994) et la mise en service de l’Écluse installée sur le canal Tinja en 1996, les quantités d’eau qui arrivent du lac Ichkeul à la lagune de Bizerte étaient de l’ordre de 165 Mm3•an-1 alors qu’après la réalisation de ces aménagements, les quantités sont estimées à 20 Mm3•an-1 (HARZALLAH, 2002).
Il est à signaler que la lagune de Bizerte fonctionne après l’installation des ouvrages avec un flux saisonnier de quatre mois/six mois, pendant les mois d’août, septembre, octobre et novembre, les eaux de la lagune passent vers le lac Ichkeul.
La diminution des apports en eau provenant de l’Ichkeul a favorisé l’entrée des eaux marines dans la lagune. Les débits mesurés sous le pont de Bizerte peuvent atteindre 3 000 m3•s-1 (équivalant à 26,3 Mm3•an-1) (HARZALLAH et KOUTITONSKY, 2001).
Le gradient moyen entre les salinités moyennes hivernale et estivale est de l’ordre de 1,9 psu. Ce gradient relativement faible montre la « marinisation » de la lagune. La salinité saisonnière dans la lagune de Thau varie entre 36 et 40 psu (IFREMER, 2006). Ces quantités importantes d’eaux marines ont influencé la variabilité spatio-temporelle de la plupart des paramètres hydrobiologiques.
La salinité moyenne des eaux de la lagune montre des valeurs rappelant celles rencontrées dans des eaux marines. HARZALLAH (2002) a montré que la salinité moyenne des eaux a augmenté de 4 psu environ après la réalisation des aménagements dans le bassin versant du lac Ichkeul. Par ailleurs, la température des eaux de la lagune reste toujours inférieure à 30 °C durant toute l’année grâce aux échanges importants d’eau avec la mer.
Les fortes concentrations en MES mesurées pendant l’hiver sont entre autres dues à un fort hydrodynamisme qui provoque la remise en suspension des sédiments de surface et un apport en sédiment par le canal Tinja qui peut atteindre des valeurs de l’ordre de 24 mg•L-1 (OUAKAD, 2007). La concentration moyenne hivernale en MES rencontrée dans la lagune de Nador (en Méditerranée, Maroc) est égale à 53 mg•L-1 (BLOUNDI, 2005) alors que la concentration moyenne en MES dans la lagune de Thau est de 9,4 mg•L-1 en 1990-1991 (GANGNERY et al., 2003). En comparant ces concentrations à celle mesurée dans la lagune de Bizerte, celle-ci peut être considérée assez chargée en matières en suspension.
Les variations saisonnières des concentrations en oxygène dissous montrent que la lagune ne présente pas de problème d’oxygénation puisque les concentrations moyennes sont souvent supérieures à 4 mg•L-1 en période estivale. Une étude plus récente (GRAMI, 2008) a montré que les teneurs en oxygène dissous en période estivale (mois de juillet et août) dans les eaux de surface sont comprises entre 5,3 et 6,5 mg•L-1.
Les teneurs en nutriments mesurées au cours de la présente étude montrent une amélioration de la qualité de ses eaux. Les teneurs maximales observées en ammonium et en phosphates sont de l’ordre de 2,3 µmol•L-1 et 0,4 µmol•L-1, respectivement. Notons que les teneurs en ammonium et en phosphates ont atteint, avant l’assainissement de la lagune, respectivement des teneurs de l’ordre de 14 mmol•L-1 (BELKHIR et al., 1985) et 7,5 mmol•L-1 (MASTOURI, 1997). Néanmoins, même si les conditions hydrobiologiques se sont améliorées, des excès d’azote et/ou du phosphore en provenance du bassin versant entraînent un phénomène d’eutrophisation et ce dysfonctionnement de l’écosystème se traduit par un déséquilibre du rapport N/P (HAMDI, 2002; HAMDI et al., 2002).
La comparaison des paramètres hydrobiologiques dans les eaux de surface de la lagune de Bizerte avec d’autres lagunes méditerranéennes montre des valeurs moyennement élevées pour l’ammonium, les nitrates et les nitrites (Tableau 2). Les teneurs importantes en ammonium sont liées à l’urbanisation du bassin versant et à la capacité épuratoire des STEPs. La présence en quantités relativement élevées du nitrate pourrait être attribuée à l’étendue assez importante du bassin versant de la lagune. Les concentrations des phosphates sont par contre de même ordre de grandeurs. Les concentrations de la chlorophylle a dans la lagune de Bizerte sont similaires à celles enregistrées dans l’étang de Thau (Tableau 2).
Tableau 2
Concentrations moyennes (min - max) de quelques paramètres hydrobiologiques dans des lagunes méditerranéennes.
Mean concentrations (min - max) of some hydrobiological parameters in Mediterranean lagoons.

La situation trophique marquée par deux pics de la biomasse chlorophyllienne est typique des milieux paraliques semi-fermés (ALONSO-LAITA et al., 2005; AZOUZ, 1966; HARTMANN et al., 2006; HERRERA-SILVEIRA, 1994). Les concentrations moyennes de la chlorophylle a montrent que les eaux de la lagune sont plus riches en phytoplancton que celles de la baie de Bizerte, considérées oligotrophe. La biomasse phytoplanctonique provenant de la mer Méditerranée augmente au fur et à mesure du déplacement des microalgues transportées par les courants giratoires dans la lagune (HARZALLAH, 2002) bénéficiant ainsi des conditions hydrologiques et trophiques du milieu. Les distributions spatiales de la chlorophylle a (Figures 5A et C) et des phéopigments (Figures 5B et D), au moment des pics, confirment que les eaux de la Méditerranée contiennent une biomasse phytoplanctonique moins dégradée par rapport à l’intérieur de la lagune qui présente une population microalgale vieillissante. En effet, leurs distributions spatiales montrent que le secteur sud-ouest, au niveau de la zone industrielle de Menzel Bourguiba, présente des conditions défavorables sous l’influence directe des rejets industriels.
Par ailleurs, l’analyse du rapport des éléments minéraux azotés et phosphorés montre que l’azote minéral compte parmi les facteurs limitants la croissance du phytoplancton dans la lagune presque durant toute l’année. Ce rapport (Tableau 3) est bien en deçà de l’optimum de Redfield (GOLDMAN et al., 1979; KARL et al., 1993; REDFIELD et al., 1963) sauf pour les mois de février et novembre où il est légèrement supérieur à 16.
Tableau 3
Facteurs limitants la croissance du phytoplancton. (-) Mesures manquantes.
Limiting factors of phytoplankton growth. (-) Missing measurements.

A l’opposé, SAKKA-HLAILI et al. (2006) ont montré que les facteurs qui limitent la croissance du phytoplancton dans la lagune sont l’azote et le phosphore. Cette différence peut être attribuée aux approches expérimentales suivies. En effet, la présente étude est basée sur l’analyse des paramètres hydrobiologiques in situ en tenant compte implicitement de l’effet des conditions du milieu (prédation, turbidité, dynamique, etc.) alors que celle de SAKKA-HLAILI et al. (2006) est basée sur l’étude de la croissance in vitro du phytoplancton dans des bouteilles enrichies en nutriments.
Ainsi, l’analyse multivariée appliquée aux mesures hydrobiologiques de la lagune de Bizerte nous a permis d’extraire les principaux modes de fonctionnement hydrobiologiques de cet écosystème. Ces deux modes caractérisent, d’une part, un premier contraste nord-sud attribué à l’effet relatif de la mer Méditerranée et du bassin versant et, d’autre part, un second contraste est-ouest attribué à l’effet du canal Tinja.
Les paramètres qui contribuent significativement à la structure nord-sud sont la température, la salinité, la transparence absolue et relative alors que ceux qui contribuent à la structure est-ouest sont la transparence absolue et relative, la chlorophylle a et le phéopigment. La contribution moyenne des éléments nutritifs à ces deux modes est faible, toutefois leurs contributions mensuelles s’avèrent importantes. Ces fluctuations sont liées directement aux variations des rejets terrestres.
Les contrastes observés dans les deux modes sont plus prononcés avant l’aménagement réalisé dans le bassin versant, en particulier les constructions des barrages. Nous supposons que ces contrastes diminuent de plus en plus que les apports en eau douce se réduisent, par conséquent, la lagune devient plus marinisée. En effet, le contraste du premier mode est essentiellement lié à trois paramètres essentiels : la température, la salinité et la transparence des eaux et, en conséquence, aux apports du bassin versant et aux échanges avec la mer. D’autre part, les paramètres qui maintiennent le contraste ouest-est sont la transparence, la chlorophylle a et le phéopigment. Une diminution des apports en eau de l’Ichkeul entraînera une réduction de la concentration en chlorophylle a dans la lagune alors que celle du phéopigment reste liée aux facteurs environnementaux.
Dans ce contexte, les études effectuées par SAKKA-HLAILI et al. (2007, 2008) distinguent deux chaînes trophiques différentes dans les secteurs nord et sud de la lagune de Bizerte. Ces dernières études confirment le mode de fonctionnement nord-sud de la lagune démontré dans la présente étude.
7. Conclusions
Cette étude a permis de mieux comprendre le fonctionnement hydrobiologique de la lagune de Bizerte. Ce fonctionnement hydrobiologique est en étroite relation avec la Méditerranée, le bassin versant de la lagune et le canal Tinja. Pour toute caractérisation hydrobiologique de la lagune, il est alors indispensable d’intégrer ces trois entités.
Parties annexes
Remerciements
Cette étude a été réalisée dans le cadre du Projet National de Recherche EL BIHERA pour la période allant de 2002 à 2006. Les auteurs remercient les réviseurs pour leurs remarques constructives. Ils remercient également Dr Anouar Hermassi pour sa contribution au niveau du traitement statistique des données.
Références bibliographiques
- ALONSO-LAITA P., N. NAVARRO, C.M. DUARTE et S. AGUSTI (2005). Seasonality of pico-phytoplankton abundance and cell death in a Mediterranean bay (Bay of Palma, Majorca Island). Vie et Milieu, 55, 177-184.
- ANPE (1990). Étude préliminaire de l’écologie du lac de Bizerte. Rapport, Agence Nationale de Protection de l’Environnement, Tunisie, 100 p.
- ANPE (2007). Rapport sur le suivi scientifique au Parc National de l'Ichkeul année 2005-2006. Rapport, Agence Nationale de Protection de l’Environnement, Tunisie, 64 p.
- AZOUZ A. (1966). Étude des peuplements et des possibilités d’Ostréiculture du lac de Bizerte. Bull. Inst. Nat. Sci. Tech. Oceanogr. Pech. Salammbô, 15, 1-69.
- BEJAOUI B., A. HARZALLAH, M. MOUSSA, A. CHAPELLE et C. SOLIDORO (2008). Analysis of hydrobiological pattern in the Bizerte lagoon (Tunisia). Est. Coast. Shelf Sci., 80, 121-129.
- BELKHIR M., H. GHABI, K. DRIDI, H. AMARA, A. KSOURI et M. HADJ ALI SALEM (1985). Le lac de Bizerte, étude physico-chimique et micro-biologique de l'écosystème lagunaire. Rapports et documents, Institut National des Sciences et Technologies de la Mer, Salammbô, Tunisie, 25 p.
- BENRAJEB-JENHANI A. et M.S. ROMDHANE (2002). Impact des perturbations anthropiques sur l’évolution du phytoplancton de la lagune de Boughrara, (Tunisie), Bull. Inst. Nat. Sci. Tech. Mer Salammbô, 29, 65-75.
- BLOUNDI M.K. (2005). Étude géochimique de la lagune de Nador (Maroc oriental). Impacts des facteurs anthropiques. Thèse de doctorat en Sciences de la Terre et de l'Univers, Faculté des Sciences de Rabat, Maroc, 210 p.
- CHAUMONT M. (1956). Hydrologie du lac Ichkeul et de ses affluents. Rapport, Archives Ministère de l’Agriculture, Tunisie, 83 p.
- DALY YAHIA M.N., O. DALY YAHIA-KEFI, S. SOUISSI, F. MAAMOURI et P. AISSA (2005). Associations Tintinnides (Ciliophora, Tintinnina)-Dinoflagellés (Dinophyceae) autotrophes potentiellement nuisibles au niveau de la baie de Tunis et de deux lagunes associées : Ghar El Melh et Tunis Sud. La Mer, 43, 19-32.
- DALY YAHIA-KEFI et O. M.N. DALY YAHIA (1997). Le phytoplancton toxique dans trois milieux lagunaires tunisiens (Ghar El Melh, Lac Sud de Tunis et Bou Ghrara). Dans : Actes Séminaire National sur la Gestion et la Conservation des Zones Humides Tunisiennes, Sousse, Tunisie, pp. 27-29.
- ESCOFIER B. et J. PAGES (1990). Analyses factorielles simples et multiples. Objectifs, méthodes et interprétation.Dunod (Éditeur), Paris, 287 p.
- FRISONI G.F., O. GUELORGET et J.P. PERTHUISOT (1986). Évaluation des potentialités aquacoles du lac de Bizerte. Projet régional méditerranéen de développement de l’aquaculture. Rapport, Institut National des Sciences et Technologies de la Mer, Salammbô, Tunisie, 44 p.
- GANGNERY A., J.M. Chabirand, F. Lagarde, P. Le Gall, J. Oheix, C. Bacher et D. Buestel (2003). Growth model of the Pacific oyster, Crassostrea gigas, cultured in Thau Lagoon (Mediterranée, France). Rev. Sci. Eau, 215, 267-290.
- GARCES E., M. MASO et J. CAMP (1999). A recurrent and localized dinoflagellate bloom in a Mediterranean beach. J. Plankton Res., 21, 2373-2391.
- GIMAZANE J.P. (1981). La production de la moule au lac de Bizerte. Bull. Off. Nat. Pech., Tunis, 5, 128-129.
- GOLDMAN J.C., J.J. McCARTHY J.J. et D.G. PEAVEY (1979). Growth rate influence on the chemical composition of phytoplankton in oceanic waters. Nature, 279, 210-215.
- GRAMI E. (2008). Variation saisonnière des nutriments dans la lagune de Bizerte – Effet de l’apport atmosphérique en nutriments. Mémoire de mastère, Hydrobiologie, Faculté des Sciences de Bizerte, Univ. 7 Novembre de Carthage, Tunisie, 59 p.
- HAMDI H. (2002). Contribution à l'étude du milieu et des copépodes pélagiques de la lagune de Bizerte (systématique et écologie numérique). Mémoire de DEA, Sciences de l’Environnement, Faculté des Sciences de Bizerte, Univ. 7 Novembre de Carthage, Tunisie, 106 p.
- HAMDI H., R. GHRIBI, N. OUERYEMI et M.N. DALY YAHIA (2002). Fluctuations printanières et estivales des communautés de copépodes pélagiques dans la lagune de Bizerte. Rev. FSB, Tunisie 1, 200-207.
- HARTMANN J.H., H. MONTAINE et L. BOURCEREAU (2006). Analyse de la succession saisonnière et de la productivité du plancton dans le bassin de Marennes-Oléron : État de l’art. Dans : Colloque Développement récents de la recherche en environnement côtier, Nantes, France, pp. 26-28.
- HARZALLAH A. (2002). État actuel et évolution de l’exploitation halieutique et aquacole des lagunes - Cas de la lagune de Bizerte. Rapport, Institut National des Sciences et Technologies de la Mer, Salammbô, Tunisie, 128 p.
- HARZALLAH A. et V.G. KOUTITONSKY (2001). Observations and numerical simulations of hydrodynamics of the Bizerte Lagoon. Dans : Proceedings of the 5th International Conference on the Mediterranean Coastal Environment, MEDCOAST 01, Hammamet, Tunisia, pp. 1101-1110.
- HASNAOUI M., J. KASSILA, M. LOUDIKI, M. DROUSSI, G. BALVAY et G. BARROIN (2001). Relargage du phosphore à l’interface eau-sédiment dans des étangs de pisciculture de la station Deroua (Béni Mellal, Maroc). Rev. Sci. Eau, 14, 307-322.
- HERRERA-SILVEIRA J.A. (1994). Phytoplankton productivity and submerged macrophyte biomass variation in a tropical coastal lagoon with groundwater discharge. Vie et Milieu, 44, 257-266.
- HIDROTECNICA PORTUGUESA (1995). Étude générale pour la protection du littoral tunisien contre l’érosion marine. Ministère de l’Équipement et de l’Habitat, Tunisie.
- HOLMES R.W. (1970). The secchi disk in turbid coastal water. Limn. Oceanogr., 15, 688–694.
- IFREMER, 2006. Réseau de suivi lagunaire du Languedoc-Roussillon : Bilan des résultats 2005. Rapport RSL-06/2006, 450 p.
- IFREMER, 2004. Réseau de suivi lagunaire du Languedoc-Roussillon : Bilan des Résultats 2005. Rapport RSL-04/2004, 523 p.
- KALLEL M.R. (1989). Hydrologie du lac de Bizerte. Rapport, DRE (Direction des Ressources en Eau)). Tunisie, 41 p.
- KARL D.M., G. TIEN, J. DORE et C.D. WINN (1993). Total dissolved nitrogen and phosphorus concentrations at US-JGOFS station Aloha: Redfield reconciliation. Mar. Chem., 41, 203-208.
- MAERH (2003). Étude sur la dépollution industrielle dans le bassin versant du lac de Bizerte. Rapport, Ministère de l’Agriculture de l’Environnement et des Ressources Hydrauliques, Tunisie, 182 p.
- MASTOURI A. (1997). Étude de l’hydrologie du lac de Bizerte. Rapport, Institut National des Sciences et Technologies de la Mer, Salammbô, Tunisie, 10 p.
- NARASIAH S., J. SHOIRY et C. MORASSE (1988). Effets des variations des températures saisonnières sur les modifications des phosphates dans des eaux usées. Rev. Sci. Eau, 1, 305-320.
- OUAKAD M. (2007). Genèse et évolution des milieux laguno-lacustres du nord-est de la Tunisie (Garaet, Lagunaires de Bizerte et Ghar et Melh). Thèse de doctorat de géologie de l’environnement, option Géologie de l’Envionnement. Faculté des Sciences de Tunis, 461 p.
- REDFIELD A.C., B.H. KETCHUM et F.A. RICHARDS (1963). The influence of organisms on the composition of sea water. Dans : Composition of sea water, comparative and descriptive oceanography. The sea: ideas and observations on progress in the study of the seas, Hill M.N. (Éditeur), Interscience Publishers, USA, NY., pp. 26-77.
- RODIER J., C. BAZIN, J.P. BROUTIN, P. CHAMBON, H. CHAMPSAUR et L. RODI (1996). L’analyse de l’eau. Eaux naturelles, eaux résiduaires, eau de mer. Dunod (Éditeur), Paris, 1383 p.
- SAKKA HLAILI A., B. GRAMI, H. HADJ MABROUK, M. GOSSELIN et D. HAMEL (2007). Phytoplankton growth and microzooplankton grazing rates in a restricted Mediterranean lagoon (Bizerte Lagoon, Tunisia). Mar. Biol., 151, 767-783.
- SAKKA HLAILI A., B. EL GRAMI, N. NIQUIL, M. GOSSELIN, D. HAMEL, M. TROUSSELLIER et H. HADJ MABROUK (2008). The planktonic food web of the Bizerte lagoon (south-western Mediterranean) during summer: I. Spatial distribution under different anthropogenic pressures. Est. Coast. Shelf Sci., 78, 61-77.
- SAKKA HLAILI A., M.A. CHIKHAOUI, B. EL GRAMI et H. HADJ MABROUK (2006). Effects of N and P supply on phytoplankton in Bizerte Lagoon (western Mediterranean). J. Exp. Mar. Biol. Ecol., 333, 79-96.
- SCHRUM C., M. ST. JOHN et I. ALEKSEEVA (2006). ECOSMO a coupled ecosystem model of the North Sea and Baltic Sea: Part II. Spatial-seasonal characteristics in the North Sea as revealed by EOF analysis. J. Mar. Sys., 61, 100-113.
- SOUSSI N. (1983). Répartition des résidus d’hydrocarbures dans les sédiments de la lagune de Bizerte (Tunisie septentrionale). Arch. Inst. Past., Tunis, 60, 21-28.
- Treguer P. et P. Le Corre (1975). Manuel d’analyse des sels nutritifs dans l’eau de mer. Laboratoire d’océanologie chimique, Bretagne, 5109 p.
- TURKI S. et A. EL ABED (1999). Sur la présence de Dinophysis spp dans les milieux lagunaires du nord de la Tunisie. Bull. Inst. Nat. Sci. Tech. Mer Salammbô, 26, 165-173.
- ZAOUALI J. (1974). Les peuplements malacologiques dans les biocoenoses lagunaires tunisiennes. Étude de l’espèce pionnière, Cerastoderma glaucum. Poiret, 1789. Thèse de Doctorat, Univ. de Caen, Caen, France, 345 p.
- ZAOUALI J. (1979). Étude écologique du lac de Bizerte. Bull. Off. Nat. Pech., Tunis, 3, 107-140.
Liste des figures
Figure 1
Situation géographique et délimitation du bassin versant de la lagune de Bizerte.
Location and delimitation of the catchment area of the Bizerte Lagoon.
Figure 2
Carte bathymétrique de la lagune de Bizerte et répartition des stations de prélèvement.
Bathymetric map of the Bizerte Lagoon and distribution of the sampling stations.
Figure 3
Variation mensuelle des paramètres physico-chimiques : température (A), salinité (B), oxygène dissous (C), matières en suspension (D) et la transparence (E). Les barres indiquent l’importance du gradient spatial du paramètre.
Monthly variation of physico-chemical parameters: temperature (A), salinity (B), dissolved oxygen (C), suspended matter (D) and transparency (E). The bars indicate the importance of the spatial gradient of the parameter.
Figure 4
Variation mensuelle de la concentration de l’ammonium (A), nitrates (B), phosphates (C), azote total (D), phosphore total (E) et de la chlorophylle a (F). Les barres indiquent l’importance du gradient spatial du paramètre.
Monthly variation of the concentration of ammonia (A), nitrate (B), phosphate (C), total nitrogen (D), total phosphorus (E) and chlorophyll-a (F). The bars indicate the importance of the spatial gradient of the parameter.
Figure 5
Distributions spatiales des concentrations en chlorophylle a et en phéopigments en avril (A et B) et en soût (C et D).
Spatial distributions of chlorophyll-a and pheopigment concentrations in April (A and B) and in August (C and D).
Figure 6
Variation temporelle (A) et distribution spatiale (B) des poids des paramètres hydro-biologiques du premier mode. L’axe des abscisse de la carte (A) représente les paramètres : Température (T), Salinité (S), Transparence (Tr), Matières en suspension (MES), Transparence relative (Tr%), Oxygène (O2), Nitrites (NO2), Nitrates (NO3), Ammonium (NH4), Azote total (NT), Phosphates (PO4), Phosphore (PT), Chlorophylle a (Chl a) et Phéopigments (Pheo). L’axe des ordonnées représente les poids des paramètres. Les poids mensuels de la température (T) et de la salinité (S) sont de même signe alors que les signes des poids mensuels des autres paramètres sont variables en fonction des mois. Ceci montre que la variabilité spatiale des paramètres est due entre autres aux effets oscillants de la Méditerranée et du bassin versant. Par exemple, les poids mensuels (carte B) de la température sont tous positifs, indiquant que la température des eaux au sud de la lagune est souvent (pour tous les mois) supérieure à celle au nord conséquence des rejets chaudes au sud de la lagune.
Series weights (A) and loadings (B) of hydrobiological parameters of the first mode. The x-axis of the chart (A) represents the parameters: Temperature (T), Salinity (S), Transparency (Tr), Suspended matter (MES), Relative transparency (Tr%), Oxygen (O2), Nitrite (NO2), Nitrate (NO3), Ammonia (NH4), Total nitrogen (NT), Phosphorus (PO4), Total phosphorus (PT), Chlorophyll-a (Chl-a) and Pheopigments (Pheo). The y-axis represents the parameter weights. The monthly weights of the temperature (T) and salinity (S) are of the same sign whereas the signs of the monthly weights of the other parameters are variable according to months. This shows that the spatial variability of the parameters is due to the oscillating effects of the Mediterranean and the catchment area. For example, the monthly weights (chart B) of the temperature are all positive indicating that the water temperature in the south of the lagoon is often higher than that of the north, a result of the thermal discharges in the south of the lagoon.
Figure 7
Variation temporelle (A) et distribution spatiale (B) des poids des paramètres hydro-biologiques du deuxième mode. Pour chaque paramètre, les poids mensuels (+) et moyen (carré) sont présentés.
Series weights (A) and loadings (B) of hydrobiological parameters of the second mode. For each parameter monthly weight (+) and mean (square) are presented.
Liste des tableaux
Tableau 1
Résultats du test ANOVA à un facteur (Scheffé F test) appliqué aux variables hydrobiologiques. Le test a été appliqué aux variables une à une, après une analyse de la variance (ANOVA). Une cellule grise indique que la différence (à un seuil de 5 %) entre les moyennes des variables est significative. Une cellule blanche indique que la différence n’est pas significative. Par exemple, la différence au niveau de 5 % des moyennes mensuelles de la température est significative sauf pour les mois de (février – décembre) et (juillet –septembre). La transparence n’est pas présentée dans ce tableau parce que les moyennes mensuelles sont significatives pour tous les mois.
Results of one factor ANOVA (Scheffé F test) applied to hydrobiological variables. The test was applied to variables taken one by one, after an analysis of variance (ANOVA). A grey cell indicates that the difference (in a 5% threshold) between the averages of variables is significant. A white cell indicates that the difference is not significant. For example, the difference at the level of 5% of the monthly averages of the temperature is significant except for (February-December) and (July-September) months. The transparency is not presented in this tableau because the monthly means are significant for all months.
Tableau 2
Concentrations moyennes (min - max) de quelques paramètres hydrobiologiques dans des lagunes méditerranéennes.
Mean concentrations (min - max) of some hydrobiological parameters in Mediterranean lagoons.
Tableau 3
Facteurs limitants la croissance du phytoplancton. (-) Mesures manquantes.
Limiting factors of phytoplankton growth. (-) Missing measurements.