Résumés
Abstract
We evaluated the vertical stratification of Colorado potato beetle (Leptinotarsa decemlineata) (CPB) populations in flight above fields of conventional potato, an insect resistant potato field expressing genes for Bacillus thuringiensis tenebrionis (Btt) toxins and a non-host clover field. Flight interception traps at 0 - 1.2, 1.2 - 2.4 and 2.4 - 3.6 m above ground were used to determine the vertical profiles of the aerial population over each crop for a growing season. The distribution of overwintered and summer CPB catches above conventional potatoes was similar with a significant effect of trap height on catch. The same vertical distribution profile was observed for overwintered CPB and a similar trend for summer CPB above the insect resistant field. Few overwintered beetles were captured over the clover field but enough summer CPBs were caught to detect a trend similar to that on resistant potatoes. Additional data show that more than 70% of CPBs fly less than 4 m in distance at a height of less than 2 m. These data suggest that the vertical flight profiles observed result from local dispersal of resident populations, or low level flights of summer beetles, but not from high level flights of dispersing overwintered adults.
Résumé
L’objectif de cette étude était de déterminer si la stratification verticale des populations du doryphore de la pomme de terre (Leptinotarsa decemlineata) en vol au-dessus de champs de pommes de terre conventionnelles est maintenue au-dessus de champs de pommes de terre exprimant les gènes pour la toxine de résistance du Bacillus thuringiensis tenebrionis (Btt) ainsi qu’au-dessus de champs non-hôtes tel un champ de trèfle. Le profil vertical de la population aérienne de doryphores au-dessus de chaque type de champ pendant une saison complète a été déterminé en utilisant des pièges à interception placés à 0 - 1.2, 1.2 - 2.4 et 2.4 - 3.6 m au-dessus du sol. La distribution des captures de doryphores de printemps et d’été au-dessus du champ de pommes de terre conventionnelles était similaire et était caractérisée par une chute significative des captures en fonction de la hauteur des pièges. La même distribution verticale fut observée pour les doryphores de printemps et une tendance similaire fut observée pour les doryphores d’été au-dessus du champ résistant à l’insecte. Très peu de doryphores de printemps furent capturés au-dessus du champ de trèfle mais suffisamment de doryphores d’été pour détecter une tendance similaire à celle observée au-dessus du champ de pommes de terre résistantes à l’insecte. Des données additionnelles obtenues au champ montrent que plus de 70 % des doryphores volent sur moins de 4 m de distance et à une hauteur de moins de 2 m. Ces données suggèrent que les profils de distribution verticale observés dans cette étude sont le résultat des vols de dispersion locale par les populations résidentes de doryphores de printemps et d’été mais ne décrivent pas les vols à plus haute altitude des doryphores de printemps en dispersion.
Corps de l’article
Introduction
The Colorado potato beetle, Leptinotarsa decemlineata (Say) [Coleoptera: Chrysomelidae] (CPB), continues to be the major defoliating pest of cultivated potatoes (Solanum tuberosum L.) in Canada and north eastern North America (Howard et al. 1994). Flight accounts for 10% to 50% of recruitment in new potato plantings each yr (Boiteau 2001) and allows adult CPB to disperse within and between fields faster than by walking (Boiteau et al. 2003). Understanding CPB flight and dispersal is important in preventing and managing infestations (Weber and Ferro 1996) and the evolution of resistance to insecticides or resistant cultivars (Alyokhin and Ferro 1999).
The horizontal dispersal by flight of CPB into, within and between potato fields and other hosts is becoming better understood (Boiteau 1986; Follet et al. 1996; French et al. 1993; Hoy et al. 1996; Noronha and Cloutier 1999; Voss and Ferro 1990a, 1990b; Weber and Ferro 1993, 1994; Weber et al. 1994, 1995; Weisz et al. 1994; Williams 1988; Zehnder and Speese 1987) but information on the vertical profile for CPB remains preliminary (Boiteau et al. 2000b).
Few studies (Boiteau et al. 1999, 2000a, 2000b) have evaluated insect vertical profiles above agricultural landscapes and the most exhaustive study on vertical distributions of insect populations in flight (Taylor 1974) was carried out above grass. The objective of this project was to determine if the vertical distribution profile of CPB in flight above the potato crop changed as beetles dispersed above CPB resistant crops and non-host crops. A secondary objective was to verify observations of Boiteau et al. (2000a) on the vertical profile of CPB. Field data on the expected positive relationship between CPB flight height and distance flown (Caprio and Grafius 1993; Voss and Ferro 1990a, 1990b) were obtained for New Bruns-wick to help interpret data on vertical profiles.
Materials and Methods
Sampling of airborne CPB
‘Harp’ style interception traps (Boiteau 2000) were used to capture CPB in flight. Harps were constructed using 1 m strands of clear fishing line (6 lb test Maxima, Santa Anna CA) hung vertically and strung 1 cm apart. Strands were attached to 1 m horizontal wooden supports top and bottom and secured with glue and an overlapping wood slat nailed in place. Harps were then mounted in metal frames, each capable of holding two harps. Finally, two lengths of 1 m plastic rain gutter were attached to both sides of the bottom support. CPB in flight impact the fishing line and fall into the plastic rain gutter. Dust from the field and a small amount of soil placed in the gutter coats the surfaces and prevents CPB from climbing out (Boiteau et al. 1994; Noronha and Clou-tier 1999). Tanglefoot (The Tanglefoot Company, Grand Rapids MI) was applied to the top and bottom supports and metal frame to trap insects alighting on the trap. Tanglefoot was also applied at the base of the frame to prevent CPB climbing into the traps. Where necessary, plants were removed from around the trap to prevent insects from climbing or falling into the gutter. Three trap heights (0 - 1.2 m, 1.2 - 2.4 m and 2.4 - 3.6 m above ground) were constructed.
Traps were arrayed in a three by three Latin square in 1 ha plots of conventional potato (cv. Russet Burbank) and insect resistant potato (cv. Russet Burbank NewLeaftm) planted on 24 May 2001, and red clover (Trifolium pra- tense L.) planted in fall 2000 at the Agri-culture and Agri-Food Canada Potato Research Centre (PRC), Fredericton, New Brunswick, Canada. Spacing between traps varied depending on plot size and shape but was set so that the distance between traps and distance from trap to edge was maximized. Traps were emptied daily between 8 a.m. and 10 a.m. from 12 June to 7 September 2001 covering the growing season from plant emergence to harvest.
All plots were located at the approximate centre of the 280 ha PRC research farm with approximately 25 ha of potatoes, ensuring the presence of a between field dispersal population of CPB. These fields as well as the clover and insect resistant potato plots were subject to typical management practices. These practices included the planting of a 0.2 ha of conventional potatoes adjacent to the insect resistant plot, as required of all growers of these type of potatoes (Caprio 2001; Kennedy and Whalon 1995). The conventional plot was managed to ensure that a population of beetles above the economic threshold for New Brunswick potato growers was present throughout the summer.
Estimating height of flight and distance flown
In the summer of 2000, observations were carried out on the height of flight and distance flown of CPB leaving a release point at the center of a potato field. Groups of 20 to 40 adult CPB were field collected and placed on a flight release apparatus consisting of a Styrofoam cone (12.5 cm diam base by 30.5 cm high) in a 2 L plastic container lined with Fluon to prevent beetles from walking out (Boiteau 2000). The flight release apparatus was elevated so that the top of the cone reached canopy height and was placed at the center of a 20 m diam observation arena. An observer (CM) was stationed at a random point 1 m from the release container. Height of flight at 1, 5 and 10 m from the cone was estimated visually and recorded (additional estimates of height of flight at 5 and 10 m from the release container were made but were not sufficient in number for analysis). The distance flown was calculated and represents the linear distance from the release point to the landing site. Flights longer than 10 m or greater than 10 m in height were recorded but were not tracked further. Observation sessions were a maximum of 2 h in duration after which the remaining beetles were returned to the field.
Statistical analysis
Two populations of adult CPB occur during the potato growing season in New Brunswick: an ‘overwintered’ population of post-diapause adults that emerge in the spring, and a ‘summer’ population that has not undergone diapause (Weber and Ferro 1996). Summer CPB in New Brunswick emerge between 25 and 30 July (Gibson et al. 1925 and personal observation) and there is little overlap in the two populations. Beetles trapped between 12 June and 29 July 2001 were classed as overwintered and those trapped after 30 July 2001 as summer CPBs. Overwintered and summer population beetles differ in their propensity for flight (Weber and Ferro 1996) and so vertical profiles were generated for both populations based on the total catch per height per d, averaged over the period in which each population was present. Our data violated assumptions of homogeneity of variance and normality of residuals. Data were therefore first ranked (PROC RANK, SAS Institute 1982) and then an analysis of variance (ANOVA) (PROC GLM, SAS Institute 1982) was performed on the ranked values, which is equivalent to a non-parametric ANOVA (Conover and Iman 1981). Pairwise comparisons between all heights were pre-planned but carried out only when a significant trend in height was found. A comparison of vertical profiles between field types was not carried out because even though it is statistically possible, the absence of field replications would prevent the attribution of causation to the type of crop.
The relation between flight height and distance flown was evaluated using linear regression (PROC REG, SAS Institute 1982).
Figure 1
Mean (± SE) total daily catch for overwintered (12 June - 29 July) and summer population (30 July - 7 Sept) Colorado potato beetles.

* denotes a significant effect of trap height on catch (ANOVA: P < 0.01). Within each graph, letters denote values which are significantly different (P < 0.01). Note y-axes differ between field types. Heights of flight interception traps are indicated by Low (0 - 1.2 m), Mid (1.2 - 2.4 m) and High (2.4 - 3.6 m), relative to ground.
Results
Vertical profile
The average number of CPB in flight decreased with the height of the traps regardless of crop type. There was a significant decrease in average total daily catch of CPB with increasing trap height above the conventional potato plot in the overwintered (F = 10.35; df = 2, 132; P < 0.01) and summer (F = 6.09; df = 2, 109; P < 0.01) populations, and above the insect resistant potato plot in the overwintered population (F = 3.90; df = 2, 135; P < 0.05) (Figs. 1a-c). A similar decrease was observed above the insect resistant potato crop and the clover crop in the summer population (Figs. 1d, f ) but almost no overwintered CPBs were trapped above clover (Fig. 1e). Comparing catches between heights revealed fewer overwintered and summer beetles were caught in high traps compared to the middle traps (overwintered: F = 13.56; df = 1, 132; P < 0.01, summer: F = 8.21; df = 1, 109; P < 0.01) and high traps compared to low traps (overwintered: F = 17.76; df = 1, 132; P < 0.01, summer: F = 9.66; df = 1, 109; P < 0.01) above the conventional potato plot, but no difference in catch was detected between low and middle traps (overwintered P = 0.59 and summer P = 0.68) (Figs. 1a, b). Fewer overwintered CPBs were caught in the high than in the low traps (F = 7.42; df = 1, 135; P < 0.01) above the insect resistant potato crop but there was no difference between high and middle (P = 0.06) and middle and low traps (P = 0.41) (Fig. 1c).
Figure 2
(a) Distance flown versus mean (± SE) height of flight of Colorado potato beetle measured at one metre from a release container, and (b) frequency of long and short distance flights.
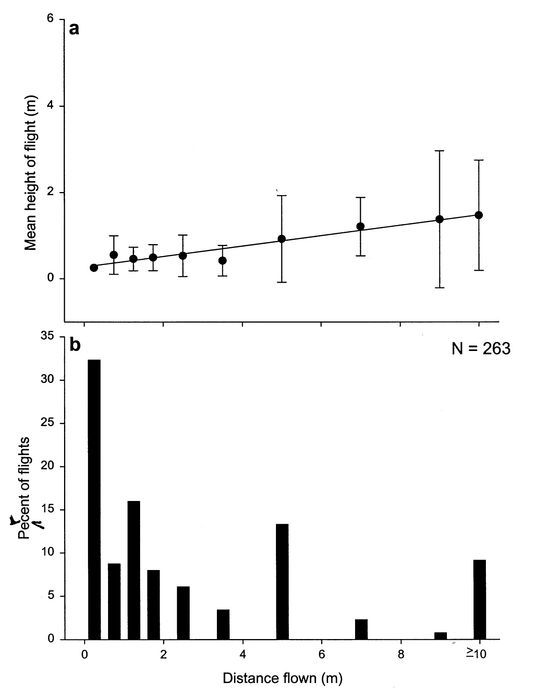
Relation between height of flight and distance flown
The height of flight of the CPB estimated at 1 m from the release point increased significantly with distance flown (r2 = 0.18, F1,156 = 34.31, P < 0.01) (Fig. 2a) within the 10 m diam arena. Although some 263 free flights were observed during 24 sessions between 26 June and 31 July 2000, most beetles made short distance flights from the cone to the ground or nearby plants, with only 10% making flights greater than or equal to 10 m (Fig. 2b).
Discussion
The CPB tends to walk, flying only when temperatures rise (Hurst 1975). This low propensity of adult CPB to take flight is characteristic of many Coleopterans (Dudley 2000) and makes it difficult to study free flight of the insect. As a consequence, research on CPB flight dispersal has been neglected, relative to walking, and is generally based on tethered flight tests (Boiteau et al. 2003).
A preliminary study described a strong, steep and negative vertical flight profile for the CPB above a potato crop, with 50% of the flights over the first 15 m above ground occurring at or below 2.3 m (Boiteau et al. 2000b). Our study using a larger number of flight records and a different style of flight interception trap, confirms the sharp drop in flight frequency at higher elevations (2.4 - 3.6 m). Also, we have established that the flight profile is qualitatively similar for the overwintered and summer populations. Seventy percent of the flights of CPB resident in a host crop were found in observations carried out in 2000 to be less than 4 m long and at a height of 2 m or less as also indicated by Voss and Ferro (1990a, 1990b) and Caprio and Grafius (1993). These flights could explain the high level of catches in the low and mid height traps.
The presence of a significant negative vertical profile above the insect resistant potato plot qualitatively similar to that above conventional potato was unexpected. There is no establishment of CPB populations in insect resistant potato fields. However, adult CPB encountering insect resistant fields can become resident for up to 31 d (Pelletier et al. 2000). It is likely that CPB from the adjacent refuge or from surrounding fields dispersed into the resistant plot and became resident. Our results suggest that during their residency, these beetles fly about in a manner similar to beetles on conventional potatoes. In the absence of a resident population, we would predict a flight activity pattern over other types of resistant potato cultivars similar to that of summer CPBs over the clover field.
Very few overwintered CPBs were caught in the clover plot perhaps because they were not abundant enough in the air layer immediately above the crop. The use of long range flights occurring above 3.6 m for between field dispersal in the search of mates and other hosts was probably another factor (Weber and Ferro 1994). The vertical distribution of summer population CPB flights obtained over clover could have resulted from a combination of walking and short flights that characterizes this insect’s dispersal at that time of the season (Weber and Ferro 1994). The relatively slow speed of this combination could have provided the residency time required to increase trap catches. Alternatively, long range and low height diapause flights proposed by Voss (Voss and Ferro 1990a) for summer beetles ready to diapause might explain the trap catches above clover. However, the number of beetles caught over clover was considerably smaller than would be expected of a mass movement to overwintering sites and more work would be required to determine the origin and destination of these beetles.
Parties annexes
Acknowledgements
D. Quiring provided valuable discussion and assistance as research supervisor to CM. R. Webster provided discussion on experimental design and comments on earlier drafts of the manuscript. P. Sandeson, H. Knight, K. Larder, I. Theriault, N. Thompson and C. Vautour provided technical assistance. This project was supported by the Matching investment initiative program of Agriculture and Agri-food Canada in collaboration with NatureMark.
References
- Alyokhin, A.V., and D.N. Ferro. 1999. Modifications in dispersal and oviposistion of Bt-resistant and Bt-susceptible Colorado potato beetle as a result of exposure to Bacillus thuringiensis subsp. tenebrionis Cry3a toxin. Entomol. Exp. Appl. 90 : 93-101.
- Boiteau, G. 1986. Effect of planting date and plant spacing on field colonization of Colorado potato beetles, Leptinotarsa decemlineata (Say), in New Brunswick. Environ. Entomol. 15 : 311-315.
- Boiteau, G. 2000. Efficiency of flight interception traps for adult Colorado potato beetles (Coleoptera: Chrysomelidae). J. Econ. Entomol. 93 : 630-635.
- Boiteau, G. 2001. Recruitment by flight and walking in a one generation Colorado potato beetle (Coleoptera: Chrysomelidae) environment. Environ. Entomol 30 : 306-317.
- Boiteau, G., Y. Pelletier, G.C. Misener, and G. Bernard. 1994. Development and evaluation of a plastic trench for protection of potato from walking adult Colorado potato beetles (Coleoptera: Chrysomelidae). J. Econ. Entomol. 87 : 1325-1331.
- Boiteau, G., Y. Bousquet, and W. Osborn. 1999. Vertical and temporal distribution of Coccinellidae (Coleoptera) in flight over an agricultural landscape. Can. Entomol. 13 : 269-277.
- Boiteau, G., Y. Bousquet, and W. Osborn. 2000a. Vertical and temporal distribution of Carabidae and Elateridae in flight above an agricultural landscape. Environ. Entomol. 29 : 1157-1163.
- Boiteau, G., W.P.L. Osborn, X. Xiong, and Y. Bousquet. 2000b. The stability of vertical distribution profiles of insects in air layers near the ground. Can. J. Zool. 78 : 2167-2173.
- Boiteau, G., A. Alyokhin, and D.N. Ferro. 2003. The Colorado potato beetle in movement. Can. Entomol. 135 : 1-22.
- Caprio, M.A. 2001. Source-sink dynamics between transgenic and non-transgenic habitats and their role in the evolution of resistance. J. Econ. Entomol. 94 : 698-705.
- Caprio, M.A., and E.J. Grafius. 1993. Movement of adult Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomeldiae), in response to isolated potato plots. Gt. Lakes Entomol. 26 : 223-231.
- Conover, W.J., and R.L. Iman. 1981. Rank transformation as a bridge between parametric and nonparametric statistics. Am. Stat. 35 : 124-129.
- Dudley, R. 2000. The biomechanics of insect flight. Princeton University Press, Prin-ceton, NJ. 476 pp.
- Follet, P.A., W.W. Cantelo, and G.K. Roderick. 1996. Local dispersal of overwintered Colorado potato beetle (Coleoptera: Chrysomelidae) determined by mark and recapture. Environ. Entomol. 25 : 1304-1311.
- French, N.M., H.P. Follet, B.A. Nault, and G.G. Kennedy. 1993. Colonization of potato fields in North Carolina by Colorado potato beetle. Entomol. Exp. Appl. 68 : 247-256.
- Gibson, A., R.P. Gorham, H.F. Hudson, and J.A. Flock. 1925. The Colorado potato beetle (Leptinotarsa decemlineata Say) in Canada. Dominion of Canada Department of Agriculture, Ottawa. 31 pp.
- Howard, R.J., J.A. Garland, and W.L. Seaman [eds.]. 1994. Diseases and pests of vegetable crops in Canada. Entomological Society of Canada/Canadian Phytopathological Society, Ottawa. 554 pp.
- Hoy, C.W., J.A. Wyman, D.A. East, and P. Kaufman. 1996. Food, ground cover and Colorado potato beetle (Coleoptera: Chrysomelidae) dispersal in late summer. J. Econ. Entomol. 89 : 963-969.
- Hurst, G.W. 1975. Meteorology and the Colorado potato beetle. Secretariat of the world meteorological organization, Geneva, Switzerland. 51 pp.
- Kennedy, G.G., and M.E. Whalon. 1995. Managing pest resistance to Bacillus thuringiensis endotoxins: Constraints and incentives to implementation. J. Econ. Entomol. 88 : 454-460.
- Noronha, C., and C. Cloutier. 1999. Ground and aerial movement of adult Colorado potato beetle (Coleoptera: Chrysome-lidae) in a univoltine population. Can. Entomol. 131 : 521-538.
- Pelletier, Y., C. Clark, G. Boiteau, and J. Feldman-Riebe. 2000. The effect of Bt-transgenic potatoes on the movement of the Colorado potato beetle [Coleoptera: Chrysomelidae]. Phytoprotection 81 : 107-114.
- SAS Institute. 1982. SAS User’s Guide: Statistics. SAS Institute, Cary, NC. 584 pp.
- Taylor, L.R. 1974. Insect migration, flight periodicity and the boundary layer. J. Anim. Ecol. 43 : 225-238.
- Voss, R.H., and D.N. Ferro. 1990a. Phenology of flight and walking by Colorado potato beetle (Coleoptera: Chrysome-lidae) adults in western Massachusetts. Environ. Entomol. 19 : 117-122.
- Voss, R.H., and D.N. Ferro. 1990b. Ecology of migrating Colorado potato beetles (Coleoptera: Chrysomelidae) in western Massachusetts. Environ. Entomol. 19 : 123-129.
- Weber, D.C., and D.N. Ferro. 1993. Distribution of overwintering Colorado potato beetle in and near Massachusetts potato fields. Entomol. Exp. Appl. 66 : 191-196.
- Weber, D.C., and D.N. Ferro. 1994. Movement of overwintered Colorado potato beetles in the field. J. Agric. Entomol. 11 : 17-27.
- Weber, D.C., and D.N. Ferro. 1996. Flight and fecundity of Colorado potato beetles (Coleoptera: Chrysomelidae) fed on different diets. Ann. Entomol. Soc. Am. 89 : 297-306.
- Weber, D.C., D.N. Ferro, J. Buonaccorsi, and R.V. Hazzard. 1994. Disrupting spring colonization of Colorado potato beetle to nonrotated potato fields. Entomol. Exp. Appl. 73 : 39-50.
- Weber, D.C., F.A. Drummond, and D.N. Ferro. 1995. Recruitment of Colorado potato beetles (Coleoptera: Chrysomelidae) to solanaceous hosts in the field. Environ. Entomol. 24 : 608-622.
- Weisz, R., Z. Smilowitz, and B. Christ. 1994. Distance, rotation and border crops affect Colorado potato beetle (Coleoptera: Chrysomelidae) colonization and population density and early blight (Alternaria solani) severity in rotated potato fields. J. Econ. Entomol. 87 : 723-729.
- Williams, C.E. 1988. Movement, dispersion, and orientation of a population of the Colorado potato beetle Leptinotarsa decemlineata (Coleoptera: Chrysomelidae), in eggplant. Gt. Lakes Entomol. 21 : 31-38.
- Zehnder, G., and J. Speese. 1987. Assessment of color response and flight activity of Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae) using window flight traps. Environ. Entomol. 16 : 1199-1202.
 10.7202/706204ar
10.7202/706204arListe des figures
Figure 1
Mean (± SE) total daily catch for overwintered (12 June - 29 July) and summer population (30 July - 7 Sept) Colorado potato beetles.
* denotes a significant effect of trap height on catch (ANOVA: P < 0.01). Within each graph, letters denote values which are significantly different (P < 0.01). Note y-axes differ between field types. Heights of flight interception traps are indicated by Low (0 - 1.2 m), Mid (1.2 - 2.4 m) and High (2.4 - 3.6 m), relative to ground.
Figure 2
(a) Distance flown versus mean (± SE) height of flight of Colorado potato beetle measured at one metre from a release container, and (b) frequency of long and short distance flights.