Corps de l’article
Qu’il devient lointain le temps du dogme central de la neurobiologie où le tissu noble, seul habilité à transporter l’information, était constitué par les réseaux de neurones, tandis que les cellules gliales étaient réduites au rôle de tissu de soutien, tout juste dignes de fournir quelques substrats énergétiques à leurs augustes voisines ! Depuis 20 ans, les travaux portant sur la macroglie, astrocytes et oligodendrocytes, ont révélé de multiples fonctions pour ces cellules, y compris des fonctions essentielles dans le domaine de la communication cellulaire [1]. La synapse, zone de contact entre deux neurones où l’information est transférée de l’un à l’autre sous forme d’amplitude de libération d’une molécule de communication, le neurotransmetteur, est devenue neuro-astrocytaire.
Astrocyte et synapse
En effet, les astrocytes jouent un rôle majeur dans l’élimination rapide des ions et des neurotransmetteurs par des systèmes de recapture puissants, permettant ainsi une régénération rapide de la capacité de transmission de l’information. L’exemple même de cette fonction est la synapse excitatrice la plus fréquemment rencontrée dans le système nerveux : la synapse glutamatergique. Les transporteurs astrocytaires, essentiellement EAAT2/GLT1 chez l’adulte, éliminent rapidement le glutamate de la synapse. Il existe de plus un couplage métabolique entre l’entrée de glutamate et la libération par l’astrocyte de glucose ou de lactate, permettant un fonctionnement harmonieux de la synapse. La perte des transporteurs, observée dans certaines maladies neurodégénératives humaines comme la sclérose latérale amyotrophique (SLA), entraîne une augmentation des concentrations de glutamate extracellulaire et une excitotoxicité qui aboutit à la perte neuronale. Il en est de même lors d’une ischémie où la modification morphologique de l’astrocyte (swelling) induit une inactivation des transporteurs du glutamate, aggravant la perte neuronale. Il est également possible d’observer des situations où les transporteurs fonctionnent à l’envers, c’est-à-dire où les astrocytes libèrent du glutamate au lieu de le recapter. Au cours des dernières années, plusieurs groupes travaillant sur un modèle de tranches d’hippocampe maintenues en survie ont mis en évidence une libération de glutamate in situ à partir des astrocytes. Cette libération provoque l’apparition de courants transitoires lents dans les neurones voisins par stimulation de récepteurs de type NMDA [2, 3].
Astrocyte et épilepsie
L’épilepsie est la pathologie neurologique la plus fréquente après la migraine. Elle se caractérise par la décharge paroxystique et hypersynchrone d’un groupe de neurones. Une des questions fondamentales de l’épileptologie réside dans la compréhension du mécanisme de synchronisation des cellules lors des crises focales. Un article du groupe de Maiden Nedergard [4] place aujourd’hui l’astrocyte au coeur du mécanisme de synchronisation. Depuis les années 60, il est connu grâce aux enregistrements EEG que la crise d’épilepsie corticale focale se caractérise par l’apparition d’une onde de dépolarisation lente de 50 à 200 millisecondes, observée dans tous les neurones au même moment et baptisée PDS (paroxysmal depolarization shift). L’analyse fine du PDS a montré qu’il était lié à un potentiel synaptique dépendant de l’activation des récepteurs AMPA, les principaux récepteurs-canaux sensibles au glutamate, et que les récepteurs NMDA, également canaux et sensibles au glutamate, jouaient un certain rôle dans la phase tardive du PDS [5]. La principale différence existant entre le PDS et les crises généralisées réside dans l’existence d’une phase inhibitrice finale lors du PDS.
Où sont nos astrocytes dans cette affaire ? Jusqu’à présent, de pauvres victimes collatérales de la pathologie neuronale. Il est effectivement bien connu que la survenue de crises comitiales provoque une modification morphologique des astrocytes avec hypertrophie du corps cellulaire et augmentation apparente de leur nombre, la gliose réactive chronique. Des enregistrements électriques lors des crises montraient également une onde de dépolarisation lente correspondant bien à la capture passive de potassium, massivement libérée à la suite de la décharge neuronale, par des cellules largement connectées grâce à des jonctions de type gap [6]. Il semble toutefois possible d’observer une gliose précédant la survenue des crises dans certains modèles. La réaction astrocytaire ne serait donc pas obligatoirement secondaire à la crise.
Le travail présenté aujourd’hui bénéficie des dernières avancées de la technologie d’imagerie cellulaire du calcium. Les auteurs ont chargé les cellules de l’hippocampe (gyrus et CA1) avec du calcium « cagé ». L’exposition à une source laser permet la libération et la brusque élévation intracellulaire du calcium. La technique utilisée, couplée à la technologie de microscopie en double photon, permet de cibler une cellule particulière et d’enregistrer les conséquences dans les cellules voisines. Lorsque le calcium est libéré au sein d’un astrocyte, on observe dans les neurones avoisinants l’apparition d’une dépolarisation typique d’un PDS. Si le calcium est libéré dans le cytoplasme d’un neurone, rien ne se passe. De façon intéressante, les auteurs ont testé 5 modèles différents de genèse des crises. Dans chaque cas, l’interférence avec la transmission neuronale n’affecte pas le PDS tandis que le blocage des vagues calciques se transmettant d’astrocytes en astrocytes ou l’inhibition de l’augmentation intracellulaire du calcium astrocytaire inhibe totalement l’apparition du PDS.
Ce travail place donc l’astrocyte au coeur du mécanisme de synchronisation de la crise. Comment s’intègrent ces données dans la véritable clinique d’une crise chez l’homme ? Au moins dans le phénomène de réentrée qui met en résonance la décharge neuronale et la réponse gliale lors de la crise. Quel est l’oeuf et quelle est la poule ? La réponse reste ouverte, mais la modulation des transporteurs astrocytaires du glutamate devient une nouvelle cible thérapeutique pour le contrôle de crises particulièrement invalidantes.
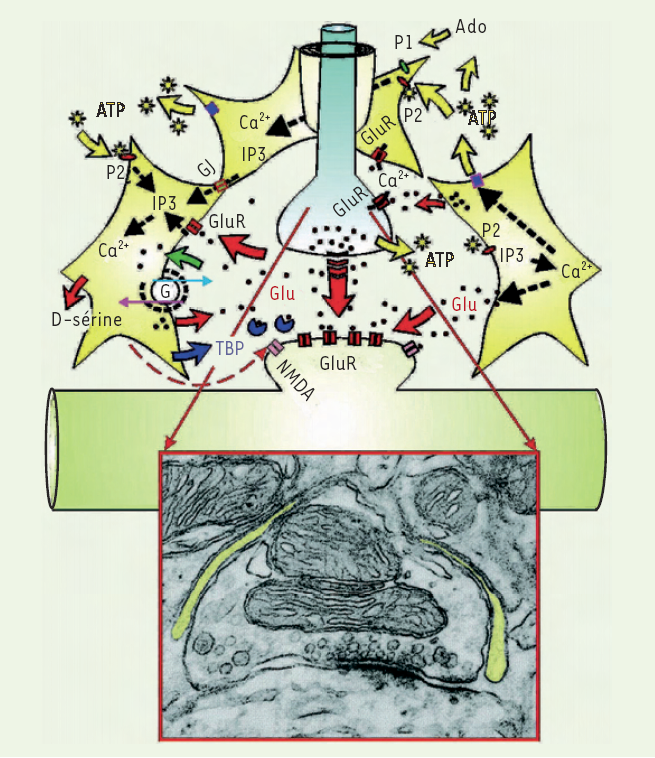
Figure 1
Les astrocytes (en jaune) participent de façon active au fonctionnement des synapses (ici glutamatergique, agrandie dans l’encart en microscopie électronique).
Parties annexes
Références
- 1. Araque A, Perea G. Glial modulation of synaptic transmission in culture. Glia 2004 ; 47 : 241-8.
- 2. Fellin T, Pascual O, Gobbo S, et al. Neuronal synchrony mediated by astrocytic glutamate through activation of extrasynaptic NMDA receptors. Neuron 2004 ; 43 : 729-3.
- 3. Angulo MC, Kozlov AS, Charpak S, Audinat E. Glutamate released from glial cells synchronizes neuronal activity in the hippocampus. J Neurosci 2004 ; 24 : 6920-7.
- 4. Tian GF, Azmi H, Takano T, et al. An astrocytic basis of epilepsy. Nat Med 2005 ; 11 : 973-81.
- 5. Dingledine R, Hynes MA, King GL. Involvement of N-methyl-D-aspartate receptors in epileptiform bursting in the rat hippocampal slice. J Physiol (Lond) 1996 ; 380 : 175-89.
- 6. Nett WJ, Oloff SH, McCarthy KD. Hippocampal astrocytes in situ exhibit calcium oscillations that occur independent of neuronal activity. J Neurophysiol 2002 ; 87 : 528-37.
Liste des figures
Figure 1
Les astrocytes (en jaune) participent de façon active au fonctionnement des synapses (ici glutamatergique, agrandie dans l’encart en microscopie électronique).