
Résumés
Abstract
Birds have been an integral part of traditional Yup’ik lifeways in the Yukon–Kuskokwim Delta, southwest Alaska, both economically and symbolically. From a subsistence point of view, the rich ethnographic record for the region highlights the importance of this resource as a critical seasonal food and a source of raw materials for clothing and tools. Little is known of bird exploitation in precontact Yup’ik society, however, as a result of limited archaeological research in the region, which thus constrains our ability to understand subsistence strategies prior to Euro-American contact. Recent excavations at the Nunalleq site (sixteenth to seventeenth century AD) have yielded a well-preserved avian assemblage that provides the opportunity to explore the use of birds during the late prehistoric period in the region. In this paper, we present the results of our preliminary zooarchaeological and technological analyses of this material. These new data demonstrate that a relatively wide range of taxa were harvested by precontact Yupiit, reflecting their multiple uses of birds. People not only exploited birds for their meat but also targeted specific taxa for the qualities of their skins for making parkas, their feathers as adornments or for fletching arrows, and their bones for making needles and other tools. Though this study shows a certain degree of continuity between precontact and historic Yup’ik subsistence practices, it also highlights a gradual decline in the non-dietary use of birds and the gradual increase in the intake of birds primarily targeted as sources of food in more recent times.
Keywords:
- Zooarchaeology,
- Alaska,
- precontact Yup’ik,
- bird exploitation,
- bone technology
Résumé
Les oiseaux ont été une partie intégrante des modes de vie traditionnels yup’ik dans le Delta du Yukon-Kuskokwim et le sud-ouest de l’Alaska, autant économiquement que symboliquement. Du point de vue de la subsistance, les riches données ethnographiques pour la région montrent l’importance de cette ressource comme un aliment critique saisonnier et une source de matériaux bruts pour la fabrication de vêtements et d’outils. L’exploitation aviaire dans la société yup’ik precontact est cependant peu connue en raison du manque de recherches archéologiques dans la région, ce qui limite notre aptitude à comprendre les stratégies de subsistance avant le contact euro-américain. Des fouilles récentes sur le site de Nunalleq (XVIe-XVIIe siècles après J.C.) ont mis au jour un assemblage aviaire bien préservé procurant l’opportunité d’explorer l’utilisation des oiseaux pendant la période préhistorique tardive dans la région. Dans cet article, nous présentons les résultats de nos analyses archéozoologique et technique préliminaires de ce matériel. Ces nouvelles données indiquent qu’une relativement grande variété d’oiseaux était exploitée par les Yupiit precontacts, reflétant ainsi l’usage diversifié qu’ils en faisaient. Les oiseaux étaient non seulement exploités pour leur viande mais certains taxons étaient également chassés pour la qualité de leurs peaux dans la fabrication de parkas, de leurs plumes comme parure ou pour l’empennage de flèches, ainsi que de leurs os pour la production d’aiguilles et autres outils. Bien que cette étude montre un certain degré de continuité entre les pratiques de subsistance des Yupiit précontact et historiques, elle souligne un déclin graduel dans l’utilisation non-carnée des oiseaux et une croissance progressive, plus récemment, de l’exploitation d’espèces principalement chassées pour la consommation.
Mots-clés:
- Archéozoologie,
- Alaska,
- yup’ik précontact,
- exploitation aviaire,
- technologie osseuse
Corps de l’article
Birds were a significant resource to precontact and historic, Arctic and subarctic communities as a source of meat, eggs, and raw materials, but they also played a central symbolic and spiritual role (Casperson 2012; Corbett 2016; Fienup-Riordan 2007; Funk 2018; Sloan 2014). As a result, bird remains are often abundant in archeological assemblages from the region (Betts 2016, 85–86) but often attract less analytical attention than other faunal remains (Casperson 2012, 20; Kristensen 2011, 293). Several zooarchaeological studies have focused on the exploitation of bird remains in Alaska (Casperson 2012; Corbett 2016; Gelvin-Reymeiller and Reuther 2010; Lefèvre and Siegel-Causey 1993; Moss 2007; Moss and Bowers 2007; Souders 1997; Sloan 2014) and in southern Greenland (Gotfredsen 1997, 2002; Gotfredsen and Møbjerg 2004). The extensive ethnographic literature can provide essential traditional knowledge on the use of birds (see Fienup-Riordan 2007; Funk 2018; Hill 2019; Hoffman 1990; Pratt 1990 for reviews) as well as an interpretative framework for the analysis of avian remains (Corbett 2016; Sloan 2014, 34).
In Southwest Alaska, excavations have been limited (Casperson 2017; Griffin 2002, 53–54; Shaw 1998; VanStone 1984; Knecht and Jones, this volume) and, as a result, few zooarchaeological studies in the region focus on avian remains, especially from the Thule era. Excavations on Nunivak Island have produced faunal assemblages of which only Souders’s (1997) study of the material from Nash Harbor (49-NI-003) is easily accessible. In the late Thule component (c. AD 1550) analyzed by Souders (1997, 15), a small avian assemblage (N = 204) represented approximately 8 per cent of the total assemblage with alcids and cormorants predominant, followed by waterfowl and gulls (Souders 1997, 41). Faunal remains were also analyzed from two Norton sites (150 BC—AD 600) by Chatters (1972, cited in Nowak 1982 and Souders 1997) and reported in an unpublished master’s thesis that we have not been able to consult, though Souders (1997, 60) and Nowak (1988) reported that anatids and gavids were predominant. In the Yukon–Kuskokwim Delta (Y–K Delta), faunal remains were recovered from relatively small-scale excavations conducted at the Manokinak site (Shaw 1982, 1985), also from the Norton period, located near the Yukon River, with the only information pertaining to birds being that caribou and birds were the most abundant in terms of raw numbers (Shaw 1985, 303–05). A limited faunal assemblage (N = 1,084) was also recovered from a small-scale excavation at the Old Togiak site (Kowta 1963) occupied between AD 1000 and 1700, Bristol Bay, with bird remains representing just under 40 per cent of the assemblage, though fish remains were not collected (Kowta 1963, 383). Cormorant and alcids predominated (Kowta 1963, 394–96). New research is being conducted at the site and will likely shed light on prehistoric subsistence as well as providing more accurate dating (Skinner and Barnett 2017). Recently, Casperson (2017) analyzed the relatively large faunal assemblage from two Norton period Summit Island sites in Bristol Bay excavated in the 1980s. She identified 1,244 bird bones to 30 taxa representing 11 families, with murres, cormorants, and eiders the most abundant birds (Casperson 2017, 200–03). This represents the largest assemblage of bird remains reported from Southwest Alaska thus far.
Recent excavations at the Nunalleq site (16–17th centuries AD) have produced an estimated thirty thousand faunal remains (Masson-MacLean et al. 2019), thus providing an unprecedented opportunity to document late precontact bird use in part due to the exceptional preservation (Britton et al. 2013, Ledger et al. 2016; Ledger et al. 2018). What role did birds play in the lifeways of the people of Nunalleq? What bird taxa are represented at the site, and how were they used by site residents—for food and to make clothes and/or bone tools? Can the study of bird remains inform us about when during the annual round people lived at Nunalleq? Were the remains (carcasses) of different bird taxa treated differently when they were discarded? How does the bird assemblage of the past compare to birds taken by Quinhagak residents today? These are some of the questions we can address through the archaeological study of the bird bones. In this paper, we report the analysis of a portion of the bird assemblage from Nunalleq combining a zooarchaeological and bone technology approach to better gauge the potential of this type of zooarchaeological study.
Background: The Yukon–Kuskokwim Delta and the Nunalleq site
The Nunalleq site is located within the Yukon–Kuskokwim Delta and more precisely on the shores of the Kuskokwim Bay near Quinhagak (Figure 1). It comprises the remains of a precontact Yup’ik sod house complex (Knecht and Jones, this volume) with several generational occupation phases dated so far between AD 1570 and 1675 (Ledger et al. 2016; Ledger et al. 2018). The Yukon–Kuskokwim Delta is one of the world’s most productive habitats for migratory birds, and the tidal flats of the coast provide extensive habitat for shorebirds. Many bird species depend on the region for much, if not all, of their nesting habitat (Darwent 2011, 45). The Yukon–Kuskokwim Delta is largely covered by water. Ponds, lakes, streams, and marshy coastal wetlands support one of the world’s largest populations of water birds, including a million ducks and half a million geese that breed there annually (US Fish and Wildlife Service 1988; Fienup-Riordan 2007, 197). The highest densities of nesting tundra swans, most of the world’s emperor geese, and 50 per cent of the world’s brant are supported by the Yukon–Kuskokwim Delta (McNab and Avers 1996). It is not surprising that birds were important resources across the region (see Hill 2019) with the Yupiit utilizing birds as sources of food (meat, eggs, fat) and of raw materials for making tools (from bones), clothing (from skins), and adornment (incorporating bones, feathers, and beaks used in ceremonial regalia, jewelry). Traditionally, a wide array of methods was used to harvest birds (nets and clubs, bolas, slings, blunt-tip or pronged arrows and pronged spears), particularly during spring (April–May) and fall (September–October) migrations and the summer moult (June–August). Migratory waterfowl arrived in the spring and were often the first visitors to the Y–K Delta, therefore providing valuable sources of food when winter supplies were low and before the food-rich summer season (Zavaleta 1999, 236). During the moult, the Yupiit would drive flightless birds on the open water with kayaks towards the shore where they were caught in nets and killed (Nelson 1899, 135). Prior to the introduction of wildlife regulations in the 1960s, this practice was common; it was economically efficient with the harvest of thousands of birds in one drive, and also contributed to the social fabric of the community as men, women, Elders, and children often took part (Fienup-Riordan 2007; Hensel and Morrow 1998). The ethnographic record and oral history also highlights the spiritual and symbolic social roles of birds. Birds were often the first prey youngsters would hunt, and a series of beliefs surrounded the gathering of eggs and nesting birds. Like Sloan (2014, 36) has documented for other Alaska Natives, Yup’ik women, “through managing the consumption of bird bodies, the use of transformative bird-bone needles and the creation of spiritually evocative bird-skin clothing,” have helped maintain spiritual connections with birds, underpinning belief systems that bridged spirituality with traditional subsistence.
Location map of the Nunalleq site

Methods and Materials
The three excavation campaigns undertaken at the Nunalleq site in 2009–2010, between 2012 and 2015, and in 2017–2018 have yielded approximately 30,000 bone specimens of which approximately 12,000 have been analyzed from the 2009–2010 (McManus-Fry 2015) and 2012–2015 (Masson-MacLean et al. 2019) field seasons. Faunal remains were hand collected and every bucket of soil was dry-screened using 1/4 inch (6.4 mm) mesh for the recovery of smaller bone fragments, including bird specimens. Bulk samples were also collected for selected contexts for wet-sieving using a 3 mm mesh but have not yet been fully analysed. The data presented in this study originate from the analysis of the 2012–2015 bird remains from the last phases of occupation at the site dating between c. AD 1620 and 1675 (Ledger et al. 2018). In total, 646 bird bones were recovered, which represents approximately 6 percent of the total number of animal bones identified from the 2012–2015 excavations. The faunal material from the 2017–2018 excavations has yet to be analyzed, including approximately 1,000 bird remains.
The Nunalleq avian material was identified using the reference collection at the Muséum National d’Histoire Naturelle in Paris (MNHN) and at the University of Aberdeen by Edouard Masson-MacLean. The osteological identifications were aided and complemented by the use of Cohen and Serjeantson (1996), combined with references in Armstrong (1995) for the geographic distribution of Alaskan birds. No attempt was made to identify bird ribs and vertebrae, and they were rare occurrences in the sample analyzed.
The faunal assemblage was recorded following established zooarchaeological methods (Driver 2011). The faunal material from Nunalleq is currently housed at the Department of Archaeology at the University of Aberdeen. After completion of the zooarchaeological analysis, the faunal material will be returned to Quinhagak under the custody of the Nunalleq Culture and Archaeology Center.
Preservation of organic materials at Nunalleq is generally excellent and the bird assemblage analyzed for this study was in very good condition with 77 per cent of specimens recorded as in good or excellent condition (n = 495) and only 5.5 per cent (n = 35) in poor condition, with exfoliation observed over 50 per cent of an individual bone specimen. Overall bird remains were in better condition than mammalian remains, of which 20 per cent were considered to be in poor condition. Carnivore teeth marks were observed on less than 1 per cent of the bones, which suggests that dog gnawing did not affect the bird assemblage; this finding contrasts with the mammalian remains, of which 15 per cent showed evidence of carnivore gnawing. The Nunalleq faunal assemblage is largely unburnt and there was no evidence of burning on the avifaunal assemblage analyzed.
Results
Taxonomic representation
From the 646 bird bones in the sample, 564 (87%) could be identified to at least the family level, comprising a minimum of 9 families and 13 genera or species. At the family level, Anatidae (swans, geese, ducks) predominated with 48% of the Number of Identified Specimens (NISP) and Laridae (gulls/terns) were also well represented (20% NISP). The other families identified included (in order of abundance) Alcidae (murres/puffins), Tetraonidae (grouses/ptarmigans), Gavidae (loons), Corvidae (ravens/crows), Phalacrocoracidae (cormorants), Strigidae (true owls), and Gruidae (cranes) (Figure 2). Ducks were the most frequent bird taxon (Table 1) both for NISP (33%) and the Minimum Number of Individuals (MNI; 23%). Large gulls—which could have included the commonly encountered Glaucous-winged gull (Larus glaucescens), Glaucous gull (L. hyperboreus), and uncommon Herring gull (L. argentus)—ranked second with 108 specimens (18%) but ranked first by MNI with 23 individuals. Swans ranked third in abundance by NISP (9%) and MNI (12%). Among anatids, although geese bones (NISP=31) were fewer than those of ducks and swans, they had an MNI of 10. Grouse/ptarmigan, murres, small loons—Red-throated loon (Gavia stellata), Pacific loon (G. pacifica), and less common Arctic loon (G. arctica)—and ravens formed non-negligible proportions of the bird assemblage. The remaining taxa were all represented by 10 or fewer specimens, including puffins, large loons (either Common loon (G. immer) or Yellow-billed loon (G. adamsii), cormorants, snowy and short-eared owls, and sandhill crane.
Bird taxonomic representation at Nunalleq

Bird taxonomic representation at Nunalleq

Bird availability and harvest data for Quinhagak and Kuskokwim Bay

Usable weights are derived from Ikuta (2016) and seasonal presence information (C=common; U=uncommon; R=rare) is from Armstrong (1995). Quinhagak = harvest estimates (Ikuta 2016); Kuskokwim Bay = harvest estimates (Wentworth 2007).
Modern bird harvests and meat weight estimates compared to bird exploitation at Nunalleq (%MNI)
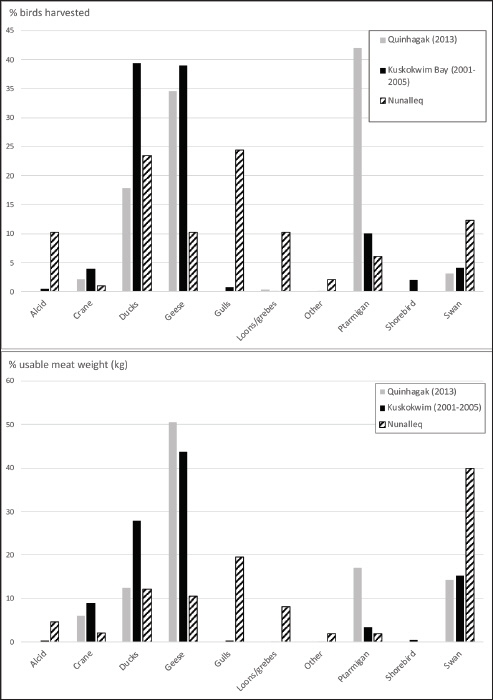
The modern data were compiled from Wentworth (2007) and Ikuta et al. (2016).
Taxonomic representation from the bird assemblage was compared, using the MNI data, to modern subsistence bird harvests by Yup’ik hunters in Quinhagak and in the wider Kuskokwim Bay region (Figure 3). The modern harvest data was compiled from Ikuta et al. (2016) for Quinhagak (2013) and Wentworth (2007) for Kuskokwim Bay (2001–2005) and presented in Table 2. Some similarities were evident between precontact and modern harvests, in particular the importance of anatids, although swans appear less frequently used, representing under 5 per cent of harvest totals, than during the precontact period where swans account for almost 12 per cent of bird MNI. Overall, an apparent difference is the greater taxonomic richness of birds that were hunted in the precontact period, which included mainly gulls (23.5%), ducks (22.5%), seabirds (21.6%), including alcids and loons, with swans (11.8%), geese (9.8%), ptarmigan (5.9%), ravens (2%), owls (2%), and cranes (1%) also exploited. By contrast, modern harvests are dominated by geese and ducks in the region though in Quinhagak ptarmigan were abundantly harvested (42%), followed by geese (34.5%) and ducks (17.9%), with swans (3.1%), cranes (2.2%), seabirds (0.3%), rare occurrences and gulls, ravens and owls not harvested. However, the eggs of seabirds, such as murres, gulls, geese, ducks, and a variety of shorebirds are still harvested in the region today (Ikuta et al. 2016, 14).
Meat weight estimates
In order to estimate the relative dietary contribution of each bird category, we used edible meat weights, a method used in zooarchaeology (Grayson 1984, 172–73) to a limited extent, but it does permit us to compare to modern subsistence studies, such as those conducted by the Subsistence Division of the Alaska Department of Fish and Game. It is calculated by multiplying the number of individuals (MNI or number of animals harvested) by the average body mass of the animal or average edible mass, often estimated at approximately 70 per cent of the total body mass for birds (Reitz and Wing 2008, 237). For this study, the usable mass for each bird species was converted from Wentworth (2007, 61) into kilograms (Table 2). Among birds at Nunalleq, swans contributed the most to the diet, providing almost 40 per cent of meat weights, followed by gulls (19.5%), ducks (12.1%), geese (10.6%), and loons (8.2%). All other birds exploited at the site would have had a minor dietary contribution (<5%). As observed for bird subsistence harvests, there are striking differences with modern subsistence meat weight estimates from Quinhagak and the region. Swans only provided approximately 15 per cent of modern avian meat weights, less than half of estimates for the precontact period. Interestingly, ptarmigan provided similar weight estimates as swans in Quinhagak (17.1%), but not in the region where the dietary contribution of ptarmigan appears much more limited (3.4%), just as it did at Nunalleq (1.8%), and these differences may reflect local ptarmigan availability. By contrast, geese contribute much more to the diet in modern times (between 40% and 50%) than estimates for the precontact period (10.6%). This difference, also observed in the harvest data, could indicate an increase in the importance of geese in Yup’ik subsistence between the precontact and modern periods.
Skeletal representation
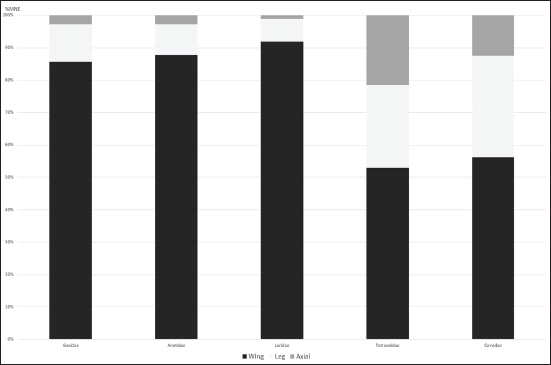
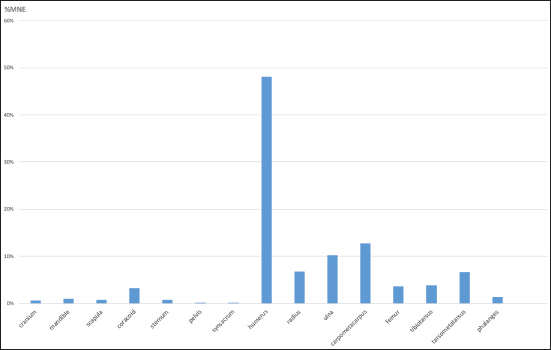
The skeletal representation of bird remains has the potential to highlight the processing of bird carcasses and cultural practises on archaeological sites (Betts 2008; Gotfredsen 1997; Lefèvre et al. 1997; Monchot et al. 2016; Moss and Bowers 2007). In this study we explored the element frequencies for anatids (swans, geese, ducks), larids (gulls), gavids (loons), ptarmigan, and raven. There are difficulties associated with the interpretation of bird skeletal representation in archaeofaunas noticeably due to the issues in understanding bone survivorship, which differs among different types of birds (see Serjeantson 2009, 155–64). As found in many assemblages (e.g., Bovy 2002, 2005), wings are often overrepresented. At Nunalleq, the humerus was by far the most frequent bird element (45.6% of identifiable elements) in the assemblage (n = 266), though differences were observed between taxonomic groups. Excellent preservation at Nunalleq is indicated by the presence of the radius in the assemblage, a fragile element with a low resistance to destructive agents (Ericson 1987) and its higher frequency than the more robust coracoid (Table 2), as well as the presence of twenty-five duck carpometacarpii, a wing element considered fragile (Serjeantson 2009, 164). This suggests that cultural processes were responsible for the representation of the various body parts observed in the avian assemblage. Very few bird vertebrae, ribs, and phalanges were present in the assemblage, and these were excluded from this analysis. Skulls were also poorly represented and the quasi-absence of these fragile elements may be related to natural attrition or represent evidence that birds were dressed elsewhere at the site given its excellent preservation overall (Serjeantson 2009, 164).
As expected, birds such as anatids, loons, and gulls were predominantly represented by elements of the wing (>85%), whereas the ground-associated ptarmigan had a higher frequency of leg and axial elements than strong flyers, though wing bones were still well represented (>40%) and the humerus was still the most frequent element among ptarmigan (Figure 4). Anatids, gulls, and loons were thus predominantly represented by the less-meatier wing elements (humerus, radius, ulna, carpometacarpus), contrasting with ptarmigan for which the meat-bearing femur and the coracoid, an element that can be associated with meat removal from the breast (Ericson 1987), represent just over 30 per cent of ptarmigan bones whereas these elements make up less than 5 per cent of anatid, gull, or loon remains (Table 3).
Element representation of selected bird taxa at Nunalleq (MNE=Minimum Number of Elements).

Body part representation of selected bird species at Nunalleq (%MNE)
Abundance of avian elements (all taxa combined) in the Nunalleq bird assemblage (%MNE).
Sex and age determination
Sex and age determination in birds can provide useful data for determining seasonality of hunting or site occupation (see Casperson 2017, 217–18; Serjeantson 2009). Bird bones only display immaturity for up to two to three months after hatching, so immature bones are good indicators of seasonality. The presence of medullary bone in breeding female bird bones can also be a good indicator of seasonality as this calcium deposit is only present shortly before and after egg laying and often occurs in spring and early summer (Serjeantson 2009, 36). There was little evidence in the avian material from Nunalleq to infer the age or sex of bird remains as bones were all mature, with the exception of one juvenile ptarmigan element, which was only relatively smaller than a fully fused humerus. Medullary bone was not observed.
Bone modifications
Small cut marks were observed on 6.3 per cent (n = 41) of bird remains, the majority of which were observed on the elements of anatids (43.9%) and gavids (41.5%). Almost all cutmarks were observed on proximal and distal humeri (n = 36) with evidence of butchery a rare occurrence on other elements, including on a raven skull and on a tarsometatarsus and tibiotarsus from a large gavid. Evidence for the use of bird bones as raw material was observed on 10.5 per cent (n = 68) of the avian remains, all humeri, mainly in the form of grooves (48.5%) and sawing (38.2%). Swan (38.2%), loons (23.5%), gulls (13.2%), and geese (10.3%) were the taxa the most commonly used with evidence of bone working also observed on puffin, murre, cormorant, and raven.
Discussion
Patterns of Abundance
The range of avian species exploited at Nunalleq during the sixteenth and seventeenth centuries is generally similar to the range of birds exploited by historic and contemporary Yupiit. Waterfowl, ptarmigan, and seabirds suggest continuity in subsistence patterns (Fienup-Riordan 2007). Some differences were observed, however, such as a greater variety of species exploited at Nunalleq when compared to modern subsistence harvests with several species no longer regularly targeted. Gulls, loons, alcids, owls, and ravens are not generally harvested today, but were still caught in the nineteenth and early twentieth centuries (Fienup-Riordan 2007). These differences appear to highlight a change in the nature of bird exploitation by the Yupiit between precontact and modern times, at least in the Quinhagak area. According to ethnographic accounts, birds were traditionally a source of food, skins, and feathers for clothing and bones for making tools (Hill 2019, 436) resulting in a variety of birds having been hunted as each bird species had its own specific qualities as a source of raw material or meat (Monchot et al. 2016). A major difference today would be that manufactured clothing is worn by most people instead of birdskin parkas. Birdskin parkas would require the labour-intensive processing and sewing of many birdskins. As Funk (2018, 155) has reflected on the Aleut use of alcids to make clothing, “Across 20 years one person would need 400–500 birds for basic clothing. A group of people living in even a small village of 20 or 30 people would need 8000 to 15,000 birds every two years.” Of course, a variety of birds can be used to make skin clothing, not just alcids. These estimates provide a sense of how different precontact life would have been when birdskins were a major source of material used to make clothing.
Traditionally, the skins of seabirds such as loons and murres were frequently used by the Yupiit to make parkas, which were also made from the skin of certain ducks and geese (Fienup-Riordan 2007, 204–07). Seabirds were particularly well represented in the avian assemblage from the site, perhaps an indication that their acquisition was driven by the desire to acquire their skins for clothing. Ethnographic and archaeological evidence documents the use of bird bones as tools in Arctic and Alaskan contexts. At Nunalleq this practice is demonstrated by the presence of longitudinal grooves on the large humeri of swans, geese, and loons related to the manufacture of bone needles (discussed below). Seabirds were a rare occurrence in modern harvests, possibly as a result of a gradual decline in the manufacture of bone needles and skin parkas in the region by the late twentieth century.
Ptarmigan represented only a small proportion of the archaeological assemblage, whereas they figured predominantly in modern harvests. Therefore, the increased importance of ptarmigan in modern Yup’ik subsistence may reflect a focus on birds as primarily a source of meat. This may also reflect bird-hunting regulations, as ptarmigan (as well as ducks and geese) are considered “game birds.” Ptarmigan at Nunalleq were also caught for food and, contrary to most species, both wing and leg elements were represented, with the femur, a meat-bearing leg bone, more prevalent among ptarmigan than for any other taxon. Caution is required as ptarmigan are renowned for mysterious changes in abundance (Weeden 2008). Although ten-year cycles have been observed in Iceland, in some parts of Alaska, ptarmigan cycles of abundance fluctuate with snowshoe hare (Merizon and Carroll 2019). Nevertheless, ptarmigan numbers can decline rapidly if the population experiences high winter losses or one or two years of poor reproduction due to a cold wet spring (Weeden 2008). It cannot be ruled out, therefore, that the lower representation of ptarmigan at Nunalleq relative to modern harvests for Quinhagak may also reflect reduced ptarmigan availability in the past as a result of cooler and wetter springs possibly experienced during the Little Ice Age.
Gulls were absent from modern harvests but ranked first (by MNI) in the Nunalleq assemblage, suggesting an abandonment of gull meat as a food source in modern times. Elders from Quinhagak regard gulls as a safety food, consumed in times of scarcity; gulls were still hunted in the earlier historic period (Andrew 2008).
Ducks were the only birds for which precontact and modern harvests appeared similar. This may reflect the importance of ducks for both clothing and food in traditional Yup’ik and Sugpiaq subsistence (Mishler 2001; Fienup-Riordan 2007). Although skin parkas were not made with the same regularity, ducks continued to be exploited as a source of food. Their bones were generally not used for manufacturing artifacts. As observed for ptarmigan, geese have been more heavily used in modern subsistence than at Nunalleq, and this may reflect the desire to acquire birds with better food returns (larger “meat packages”) today.
Ravens and owls were a rare occurrence in the faunal assemblage but were absent from modern harvests. This may reflect cultural changes in human–bird relationships. Traditionally, birds such as snowy owls were kept as pets (Fienup-Riordan 2007, 211), and as they fed on waste, ravens were not usually hunted except to make “firebath” (sauna) hats or skin bags.
After contact and the introduction of Euro-American goods, there appears to have been a gradual shift in the use of birds from serving as sources of food, feathers, skins, and bones as raw material to use as food, almost exclusively. The modern exploitation of birds is largely restricted to ptarmigan, geese, and ducks. This contrasts quite significantly with the findings from the analysis of the Nunalleq assemblage which suggest that, as observed in the ethnographic record, a range of factors influenced the selection of birds by precontact Yupiit, including the value of their non-food products.
Seasonality
The presence of the juvenile ptarmigan bone may indicate an individual caught in the late summer. Most Alaskan ptarmigan chicks hatch in late June or early July (Alaska Department Fish and Game n.d.), which more or less coincides with the start of modern subsistence hunting of willow ptarmigan in Quinhagak, a period that extends from early autumn to early spring (Wolfe et al. 1984, 316). The great majority of the birds exploited at Nunalleq, such as anatids and gulls, are seasonal residents in the Y–K Delta (Table 2), arriving in the spring and leaving late summer or early fall (Armstrong 1995).
Modern subsistence studies in the Y–K Delta (Wentworth 2007; Ikuta et al. 2016) have shown that over two-thirds of birds are taken in the spring, including migratory birds and the resident ptarmigan, most of which were hunted in the early spring between early April and mid-May. The taking of birds declines significantly in June as birds begin to nest and become less available. Bird use is at its lowest point during midsummer, when birds are raising their young and the focus of Yup’ik subsistence is salmon fishing. Few birds are taken in the summer, and bird hunting is again practiced in late August and September, when staging and the fall migrations begin, with approximately a quarter of birds harvested in the early fall.
There is some seasonal variability in the hunting of birds depending on the species exploited, though most birds are caught in the spring. The majority of geese, swans, cranes, gulls, and most seabirds, ptarmigan, and ducks were taken in the spring, though dabbling ducks were mainly hunted in the fall and were also taken during summer and spring, and ptarmigan were also hunted in the late winter “providing a much-needed source of meat during the lean late winter and early spring months” (Ikuta et al. 2016, 139). Loons were mainly hunted in the spring and summer, though the red-throated loon was almost exclusively taken in the spring and the Pacific loon was equally taken in the spring and fall (Wentworth 2007). Modern harvests also show that the south coast region of the Y–K Delta is the region where shorebirds are frequently harvested, predominantly godwits, occasionally with plovers and curlews (Naves 2009), mainly in the summer and fall, though their remains were not identified in the Nunalleq faunal assemblage.
Modern harvests echo ethnographic accounts regarding the seasonality of bird hunting in the region with the notable difference of not harvesting flightless molting birds, such as sea ducks (eiders, scoters), which molt on the ocean, during the summer months (Hill 2019 437; Andrew 2008, 159–63). This practice has since been abandoned due to wildlife regulations introduced in the latter half of the twentieth century. The avian taxonomic representation strongly suggests Nunalleq was occupied during the spring, and the presence of ptarmigan also suggests late winter occupation. Geese are also traditionally hunted in the late summer and early fall in the Y–K Delta, and their presence in the Nunalleq assemblage may also support occupation during that time of the year.
Human activities associated with bird remains
Given the relatively small size of birds compared to larger mammals, and the presence of most skeletal elements, it is likely that birds were brought back complete to the site to be processed. Body part profiles reflect on-site carcass processing. During the historic period, Yupiit consumed the meat of anatids, loons, gulls, and ptarmigan (Fienup-Riordan 2007; Wentworth 2007), and this was also likely during precontact times. The data presented here suggest that ptarmigan were primarily targeted as a source of food, whereas waterfowl, loons, and gulls must also have been highly valued for their non-food utility. The ethnographic literature depicts various technical and domestic Yup’ik usages of bird wings, such as the use of swan wings as brooms or the use of certain bird feathers in clothing or to fletch arrows (Fienup-Riordan 2007). The abundance of humeri in certain species, such as swans, geese, and large loons (Figure 5) may also reflect a desire to acquire this element as a source of raw material for making tools and evidence for worked humeri and radii have been observed in the Nunalleq faunal record and are discussed below.
Evidence of butchery was extremely rare on bird remains and included short cutmarks at the distal and proximal ends of humeri, which were probably related to the disarticulation of the wing. No cutmarks could be definitively associated with skinning, even though the use of bird skins to make parkas is widely reported in the Yup’ik ethnographic record (Barker and Barker 1993; Fienup-Riordan 2007; Hill 2019; Nelson 1899). Almost all bone modifications observed on avian remains could be associated with the osseous industry. The most common modification were longitudinal grooves, mainly observed on swan, goose, loon, cormorant, raven, and gull proximal and distal humeri fragments, and absent from all other taxa including ducks. These grooves were often combined with sawing marks and relate to the manufacture of needles; they have also been observed at various other Alaskan coastal sites (Crockford et al. 2004; Lefèvre et al. 1997). The process involves making grooves along the length of the humerus, sawing off the distal and proximal ends, and then lifting the needle (Gelvin-Reymiller and Reuther 2010; Monchot et al. 2016). This process would explain the lack of identifiable humeri shaft fragments in the assemblage. The preference for the humerus is understandable as it is a long, dense, and straight bone, especially on larger taxa such as geese and swans. Geese humeri (c. 1.5–2 mm) have a greater thickness and seem to have been preferred for the manufacture of needles (Figure 6). Swan humeri (1–1.5 mm) are slightly thinner and may have been used mostly for the manufacture of the sharpened tip used for some darts (Figure 7). The other type of modification was the polishing of split ends possibly to make a point, especially on radii. Some scrape marks on ulnae were also observed and may relate to the removal of feathers. Wing feathers were traditionally used in Yup’ik clothing and technology for decorating garments (Fienup-Riordan 2007, 48, 204) and fletching arrows, respectively (Fienup-Riordan 2007, 138, 168). Some of the preferred taxa among Yup’ik Elders for the use of their feathers in archery include cormorants, geese, cranes, and snowy owls (Fienup-Riordan 2007, 172), all of which have been identified at Nunalleq.
Cygnus sp. proximal left humerus (GDN248-13102- 37-2) showing grooves which permitted the extraction of at least four splinters

Conclusion
Compared to Western Thule and other coastal Alaska sites (Betts 2016, 85–86; Causey et al. 2005; Moss and Bowers 2007), the avian assemblage from Nunalleq is relatively small. Yet even this preliminary analysis highlights the important roles birds have played in the lives of the people who lived at Nunalleq. The ethnographic record demonstrates that birds played critical roles at specific times of the year in the Y–K Delta, especially in the early spring when stored winter supplies were low or if seals arrived later than usual in the region (Moss and Bowers 2007; Andrew 2008). In terms of total MNI, birds ranked high, suggesting that the remains of numerous birds were present in the assemblage. Birds were clearly important sources of food, but in addition, they provided the raw materials for clothing on which people relied for survival in the region. Birds may have only contributed marginally to the diet on an annual basis according to meat weight estimates and isotope analysis (Britton et al. 2018; Masson-MacLean et al. 2019), but may have supplied critical fresh meat supplies in the spring. Their economic importance to the people at Nunalleq went beyond food, to include all the body parts that served as raw materials to make other items of material culture, especially clothing and bone sewing and hunting implements. We expect that the utility of different bird taxa varied, and hope that future analyses can yield more specific identifications and specialized knowledge of different species.
Different production waste representing the initial stage of the manufacture of needles and sharpened dart tips probably used as gaming darts.

Parties annexes
Acknowledgements
This work was funded by an AHRC-LabEx award (AH/N504543/ 1) to Kate Britton, RK, Keith Dobney, and Isabelle Sidéra, and an Arts and Humanities Research Council (AH/K006029/1) grant awarded to RK, Kate Britton, and Charlotta Hillerdal (Aberdeen). The Carlsberg Foundation (Denmark) also supported this research through the Distinguished Postdoctoral Fellowship attributed to CH.
The onsite collection of samples was carried out by staff and students from the University of Aberdeen, volunteer excavators and the residents of Quinhagak. We had logistical and planning support for fieldwork by the Qanirtuuq Incorporated, Quinhagak, Alaska, and the people of Quinhagak, who we also thank for sampling permissions. Special thanks to Warren Jones, John Smith and Qanirtuuq Incorporated (especially Michael Smith and Lynn Church), and to all project team members, in Aberdeen and at other institutions. Special thanks to Christine Lefèvre (Muséum National d’Histoire Naturelle, Paris) for facilitating access to the bird reference collection at the Muséum and sharing her knowledge of avian remains during the identification process.
References
- Alaska Department of Fish and Game. n.d. “Willow Ptarmigan.” http://www.adfg.alaska.gov/index.cfm?adfg=willowptarmigan.main.
- Andrew, Frank Miisaq, Sr. 2008. Paitarkiutenka: My Legacy to You. Edited by Ann Fienup-Riordan. Seattle: University of Washington Press with Anchorage Museum Association and Calista Elders Council.
- Armstrong, Robert H. 1995. Guide to the Birds of Alaska. Portland, OR: Alaska Northwest Books.
- Barker, James H. and Robin Barker. 1993. Always Getting Ready / Upterrlainarluta: Yup’ik Eskimo Subsistence in Southwest Alaska. Seattle: University of Washington Press.
- Betts, Matthew W. 2008. Subsistence and Culture in the Western Canadian Arctic: A Multicontextual Approach. Mercury Series Archaeology Paper 169. Canadian Museum of Civilization, Gatineau, QC.
- Betts, Matthew W. 2016. “Zooarchaeology and the Reconstruction of Ancient Human–Animal Relationships in the Arctic.” In The Oxford Handbook of The Prehistoric Arctic, edited by M.T. Friesen and O.K. Mason, 81–108. New York: Oxford University Press.
- Bovy, Kristine. M. 2002. “Differential Avian Skeletal Part Distribution: Explaining the Abundance of Wings.” Journal of Archaeological Science 29, no. 9: 965–78.
- Bovy, Kristine. M. 2005. “Effects of Human Hunting, Climate Change and Tectonic Events on Waterbirds along the Pacific Northwest Coast during the Late Holocene.” PhD diss., University of Washington.
- Britton, Kate, Rick Knecht, Olaf Nehlich, Charlotta Hillerdal, Richard S. Davis, and Michael P. Richards. 2013. “Maritime Adaptations and Dietary Variation in Prehistoric Western Alaska: Stable Isotope Analysis of Permafrost-Preserved Human Hair.” American Journal of Physical Anthropology 151, no. 3: 448–61.
- Britton, Kate, Ellen McManus-Fry. Olaf Nehlich, Mike Richards, Paul M. Ledger, and Rick Knecht. 2018. “Stable Carbon, Nitrogen and Sulphur Isotope Analysis of Permafrost Preserved Human Hair from Rescue Excavations (2009, 2010) at the Precontact Site of Nunalleq, Alaska.” Journal of Archaeological Science: Reports 17 (February): 950–63.
- Casperson, Molly R. 2012. “The Importance of Birds in Ocean Bay Subsistence: Results from the Mink Island Site, Katmai National Park and Preserve, Alaska.” Arctic Anthropology 49, no 1: 18–34.
- Casperson, Molly R. 2017. “Walrus, Seal, and Seabird Faunal Remains from Summit Island in Bristol Bay, Alaska: The Subsistence Practices of Norton Peoples in an Island Environment (2740–980 Cal B.P.).” PhD diss., University of Oregon.
- Causey, Douglas et al. 2005. “The Palaeoenvironment of Humans and Marine Birds of the Aleutian Islands: Three Millennia of Change.” Fisheries Oceanography 14, no. S1: 259–76.
- Chatters, James C. 1972. “The Faunal Analysis of Sites MK2 and MK4, Nunivak, Alaska.” Master’s thesis on file. University of Washington, Seattle. (Not consulted).
- Cohen, A., and D. Serjeantson. 1996. A Manual for the Identification of Bird Bones from Archaeological Sites. London: Archetype Publications.
- Corbett, Debra. 2016. “Saĝdaĝ—To Catch Birds.” Arctic Anthropology 53, no. 2: 93–113.
- Crockford, S.J. et al. 2004. “Final Report on the Analysis of the Vertebrate Fauna from Amaknak Bridge, Unalaska, AK, UNL050.” In Amaknak Bridge Site Data Recovery Project Final Report, edited by R. Knecht and R. Davis. Report on file, Museum of the Aleutians, Unalaska, AK,
- Darwent, Christyann M. 2011. “Archaeological and Ethnographic Evidence for Indigenous Hunting and Fishing Economies in the North American Arctic and Subarctic.” In Subsistence Economies of Indigenous North American Societies: A Handbook, edited by B.D. Smith, 31–64. Lanham, MD: Smithsonian Institution Scholarly Press.
- Driver, Jonathan C. 2011. “Identification, Classification and Zooarchaeology.” Ethnobiology Letters 2:19–39. https://doi.org/10.14237/ebl.2.2011.32.
- Ericson, Per G.P. 1987. Interpretations of Archaeological Bird Remains: A Taphonomic Approach. Journal of Archaeological Science 14, no. 1: 65–75.
- Fienup-Riordan, Ann. 2007. Yuungnaqpiallerput / The Way We Genuinely Live: Masterworks of Yup’ik Science and Survival. Seattle: University of Washington Press.
- Funk, Caroline. 2018. “Ethno-Ornithology in the Rat Islands: Prehistoric Aleut Relationships with Birds in the Western Aleutians, Alaska.” Journal of Anthropological Anthropology 51: 144–58.
- Gelvin-Reymiller, Carol, and Joshua Reuther. 2010. “Birds, Needles, and Iron: Late Holocene Prehistoric Alaskan Grooving Techniques.” Alaska Journal of Anthropology 8, no. 1: 1–22.
- Gotfredsen, Anne Birgitte. 1997. “Sea Bird Exploitation on Coastal Inuit Sites, West and Southeast Greenland.” International Journal of Osteoarchaeology 7, no. 4: 271–86.
- Gotfredsen, Anne Birgitte. 2002. “Former Occurrences of Geese (Genera Anser and Branta) in Ancient West Greenland: Morphological and Biometric Approaches. Acta zoologica cracoviensia 45 (Special issue): 179–204.
- Gotfredsen, A.B., and T. Møbjerg. 2004. Society Nipisat—A Saqqaq Culture Site in Sismiut, Central West Greenland. Copenhagen: SILA and Danish Polar Center.
- Grayson, Donald K. 1984. Quantitative Zooarchaeology: Topics in the Analysis of Archaeological Faunas. Orlando, FL: Academic Press.
- Griffin, Dennis. 2002. “A History of Human Settlement on Nunivak Island, Alaska: Insights from Recent Investigations at Nash Harbor Village.” Arctic Anthropology 39, no.1–2: 51–68.
- Hensel, Chase, and Phyllis Morrow. 1998. “Co-Management and Co-Optation: Alaska Native Participation in Regulatory Processes.” Cultural Survival Quarterly 22, no. 3: 69–71.
- Hill, Erica 2019. “Humans, Birds and Burial Practices at Ipiutak, Alaska: Perspectivism in the Western Arctic.” Environmental Archaeology 24, no. 4: 434–48.
- Hoffman, Brian W. 1990. “Bird Netting, Cliff-Hanging, and Egg Gathering: Traditional Procurement Strategies on Nunivak Island.” Arctic Anthropology 27, no. 1: 66–74.
- Ikuta, Hiroko, David Michael Runfola, James J. Simon, and Marylynne L. Kostick. 2016. Subsistence Harvests in 6 Communities on the Bering Sea, in the Kuskokwim River Drainage, and on the Yukon River, 2013. Technical Paper No. 417. Alaska Department of Fish and Game, Division of Subsistence, Fairbanks.
- Knecht, Rick, and Warren Jones. 2019. “‘The Old Village’: Yup’ik Precontact Archaeology and Community-Based Research at the Nunalleq Site, Quinhagak, Alaska. ” Études Inuit Studies 43, no. 1–2, this volume.
- Kowta, Makota. 1963. “Old Togiak in Prehistory.” PhD diss., University of California.
- Kristensen, Todd J. 2011. “Seasonal Bird Exploitation by Recent Indian and Beothuk Hunter-Gatherers of Newfoundland.” Canadian Journal of Archaeology/Journal Canadien d’Archéologie 35, no. 2: 292–322.
- Ledger, Paul M., Véronique Forbes, Edouard Masson-MacLean, and Richard A. Knecht. 2016. “Dating and Digging Stratified Archaeology in Circumpolar North America: A View from Nunalleq, Southwestern Alaska.” Arctic 69, no. 4: 378–90.
- Ledger, Paul M., Véronique Forbes, Edouard Masson-MacLean, Charlotta Hillerdal, Derek W. Hamilton, Ellen McManus-Fry, Ana Jorge, Kate Britton, and Richard A. Knecht. 2018. “Three Generations under One Roof? Bayesian Modeling of Radiocarbon Data from Nunalleq, Yukon-Kuskokwim Delta, Alaska.” American Antiquity 83, no. 3: 505–24.
- Lefèvre, Christine, and Douglas Siegel-Causey. 1993. “First Report on Bird Remains from Buldir Island, Aleutian Islands, Alaska.” Archaeofauna 2: 83–96.
- Lefèvre, Christine, Debra G. Corbett, Dixie L. West, and Douglas Siegel-Causey. 1997. “A Zooarchaeological Study at Buldir Island, Western Aleutians, Alaska.” Arctic Anthropology 34, no. 2: 118–31.
- Masson-MacLean, Edouard, Claire Houmard, Rick Knecht, Isabelle Sidéra, Keith Dobney, and Kate Britton. 2019. “Pre-Contact Adaptations to the Little Ice Age in Southwest Alaska: New Evidence from the Nunalleq Site.” Quaternary International, corrected proof available online May 3, 2019. https://doi.org/10.1016/j.quaint.2019.05.003.
- McManus-Fry, Ellen Teresa. 2015. “Pre-Contact Ecology, Subsistence and Diet on the Yukon-Kuskokwim Delta: An Integrated Ecosystem Approach to Pre-Contact Arctic Lifeways Using Zooarchaeological Analysis and Stable Isotope Techniques. PhD diss., University of Aberdeen.
- McNab, W.H., and P.E. Avers. 1996. Ecological Subregions of the United States. USDA Forest Service, Washington Office. Accessed December 5, 2018. https://www.fs.fed.us/land/pubs/ecoregions/.
- Merizon, Richard A.J., and Cameron J. Carroll. 2019. Status of Grouse, Ptarmigan, and Hare in Alaska, 2017 and 2018. Alaska Department of Fish and Game, Wildlife Management Report ADF&G/DWC/WMR-2019-2, Juneau.
- Mishler, Craig. 2001. Black Ducks and Salmon Bellies: An Ethnography of Old Harbor and Ouzinkie. Anchorage, AK: Donning Company Publishers.
- Monchot, Hervé, Andrea Thompson, Benjamin Patenaude, and Claire Houmard. 2016. “The Role of Birds in the Subsistence Practices of the Dorset Peoples of Nunavik.” Arctic Anthropology 53, no. 1: 69–83.
- Moss, Madonna L. 2007. “Haida and Tlingit Use of Seabirds from the Forrester Islands, Southeast Alaska.” Journal of Ethnobiology 27 (Summer): 28–45.
- Moss, Madonna L., and Peter M. Bowers. 2007. “Migratory Bird Harvest in Northwestern Alaska: A Zooarchaeological Analysis of Ipiutak and Thule Occupations from the Deering Archaeological District.” Arctic Anthropology 44, no. 1: 37–50.
- Naves, Liliana C. 2009. Alaska Migratory Bird Subsistence Harvest Estimates, 2004–2007, Alaska Migratory Bird Co-Management Council. Anchorage: Alaska Department of Fish and Game, Division of Subsistence.
- Nelson, Edward W. 1899 The Eskimo about Bering Strait. Washington, DC: Government Printing Office.
- Nowak, Michael. 1988. “The Late Prehistoric Development of Alaska’s Native People Post-Norton Nunivak: A Study in Coastal Adaptation.” In The Late Prehistoric Development of Alaska’s Native People, edited by Robert D. Shaw, Roger K. Harritt, and Don E. Dumond, 149–67. Anchorage: Alaska Anthropological Association.
- Pratt, Kenneth L. 1990. “Economic and Social Aspects of Nunivak Eskimo ‘Cliff-Hanging.’” Arctic Anthropology 27, no. 1: 75–86.
- Reitz, Elizabeth J., and Elizabeth S. Wing. 2008. Zooarchaeology. New York: Cambridge University Press.
- Serjeantson, Dale. 2009. Birds. Cambridge: Cambridge University Press.
- Shaw, Robert D. 1982. “The Expansion and Survival of the Norton Tradition on the Yukon-Kuskokwim Delta.” Arctic Anthropology 19, no. 2: 59–73.
- Shaw, Robert D. 1985. “The Archaeology of the Manokinak Site: A Study of the Cultural Transition between Late Norton Tradition and Historic Eskimo.” PhD diss., Washington State University.
- Shaw, Robert D. 1998. “An Archaeology of the Central Yupik: A Regional Overview for the Yukon-Kuskokwim Delta, Northern Bristol Bay, and Nunivak Island.” Arctic Anthropology 35, no. 1: 234–46.
- Skinner, Dougless, and Kristen D. Barnett. 2017. “Togiak Archaeological and Paleoecological Project: Exploring Relationships and Ecology at the Old Togiak Village.” Proceedings from Presented at the 81st Annual Meeting of the Society for American Archaeology, Vancouver, British Columbia. tDAR id: 430534. https://core.tdar.org/document/430534/togiak-archaeological-and-paleoecological-project-exploring-relationships-and-ecology-at-the-old-togiak-village.
- Sloan, Anna C. 2014. “Spirituality and the Seamstress: Birds in Ipiutak and Western Thule Lifeways at Deering, Alaska.” Arctic Anthropology 51, no. 2: 35–59.
- Souders, Paul. 1997. Ellikarrmiut Economy: Animal Resource Use at Nash Harbor (49-NI-003), Nunivak Island, Alaska. Unpublished MSc thesis, University of Oregon.
- US Fish and Wildlife Service. 1988. Yukon Delta National Wildlife Refuge: Comprehensive Conservation Plan, Environmental Impact Statement, Wilderness Review and Wild River Plan. Anchorage: US Fish and Wildlife Service.
- VanStone, James M. 1984. “Mainland Southwest Alaska Eskimo.” In Handbook of North American Indians, vol. 5, Arctic, edited by D. Damas, 224–42. Washington, DC: Smithsonian Institution Press.
- Weeden, Robert B. 2008. “Ptarmigan.” Alaska Department of Fish and Game. http://www.adfg.alaska.gov/static/education/wns/ptarmigan.pdf.
- Wentworth, Cynthia. 2007. Subsistence Migratory Bird Harvest Survey: Yukon-Kuskokwim Delta 2001–2005 with 1985–2005 Species Tables. Anchorage: US Fish and Wildlife Service.
- Wolfe, Robert J., Joseph J. Gross, Steven J. Langdon, John M. Wright, George K. Sherrod, Linda J. Ellanna, Valerie Sumida, and Peter J. Usher. 1984. Subsistence-Based Economies in Coastal Communities of Southwest Alaska. Technical Paper No. 89. Juneau: Alaska Department of Fish and Game.
- Zavaleta, Erika. 1999. “The Emergence of Waterfowl Conservation among Yup’ik Hunters in the Yukon-Kuskokwim Delta, Alaska.” Human Ecology 27, no. 2: 231–66.
Liste des figures
Location map of the Nunalleq site
Bird taxonomic representation at Nunalleq
Modern bird harvests and meat weight estimates compared to bird exploitation at Nunalleq (%MNI)
The modern data were compiled from Wentworth (2007) and Ikuta et al. (2016).
Body part representation of selected bird species at Nunalleq (%MNE)
Abundance of avian elements (all taxa combined) in the Nunalleq bird assemblage (%MNE).
Cygnus sp. proximal left humerus (GDN248-13102- 37-2) showing grooves which permitted the extraction of at least four splinters
Different production waste representing the initial stage of the manufacture of needles and sharpened dart tips probably used as gaming darts.
Liste des tableaux
Bird taxonomic representation at Nunalleq
Bird availability and harvest data for Quinhagak and Kuskokwim Bay
Usable weights are derived from Ikuta (2016) and seasonal presence information (C=common; U=uncommon; R=rare) is from Armstrong (1995). Quinhagak = harvest estimates (Ikuta 2016); Kuskokwim Bay = harvest estimates (Wentworth 2007).
Element representation of selected bird taxa at Nunalleq (MNE=Minimum Number of Elements).