Résumés
Résumé
Dans un contexte de peuplement, les premiers immigrants sont habituellement les plus représentés dans le bassin génétique de la population. La contribution différentielle des immigrants fondateurs de la population de l’Isle-aux-Coudres est évaluée ici à l’aide du Registre de la population et du recensement ethnologique de 1967; le rôle de la parenté biologique ainsi que du comportement matrimonial et reproducteur des descendants de ces fondateurs est étudié à l’aide des mesures du coefficient de parenté biologique, de la probabilité d’origine des gènes et des fins d’information généalogique. L’immigration, de type familial au début, a été accompagnée puis suivie de migration individuelle. Les générations ultérieures ont puisé leurs gènes dans ce bassin initial d’immigrants et dans les ajouts subséquents. Par ailleurs, ces ajouts n’ont pas nécessairement contribué à diversifier le patrimoine génétique de la population puisqu’on a pu constater qu’un bon nombre d’unions exogames étaient célébrées entre apparentés provenant majoritairement de Charlevoix. Les unions impliquant des apparentés ont eu une fécondité exprimée dans l’île plus importante, ce qui a contribué à l’homogénéisation du pool génique; les individus en âge de se reproduire en 1967 descendent d’un noyau restreint de fondateurs.
Abstract
In a context of land settlement, the first immigrants are generally most strongly represented in the population’s gene pool. This study uses the Registre de la population et du recensement ethnologique de 1967 (1967 ethnological census and population register) to evaluate the differential contribution of the immigrant founders of the population of Isle-aux-Coudres (an island in the St. Lawrence River east of Quebec City); it examines the role of kinship and of the marital and reproductive behaviours of the descendants of these founders based on measures of kinship coefficient, probability of gene origin and genealogical information concerning the first known ancestor. The early colonisation period was initially characterised by familial immigration, followed by individual immigration. Subsequent generations have drawn their genes from this initial pool of immigrants and from later arrivals. Moreover, these later arrivals did not necessarily help to diversify the population’s genetic heritage: the authors found a large number of exogamous unions with related individuals, most of whom were from the Charlevoix region. On the island, unions involving inbreeding proved to be more fertile, which contributed to homogenization of the gene pool; individuals of reproductive age in 1967 descend from quite a small core of founders.
Corps de l’article
Les études de génétique des populations ou de démographie génétique et historique ont montré que, dans un contexte de peuplement, quelle que soit la période considérée, les premiers immigrants, qui sont à l’origine de l’établissement initial, sont habituellement les plus représentés dans le bassin génétique de la population lorsque celle-ci est examinée en termes de contribution différentielle de ses fondateurs [1].
La mesure de la probabilité d’origine des gènes (Jacquard, 1972) est un bon outil pour évaluer cette représentation différentielle des fondateurs dans des pools géniques successifs ou à un moment unique. Bien qu’ils aient une probabilité beaucoup plus faible d’être représentés (1/2n) dans une population compte tenu du nombre de générations (n) les séparant de leurs descendants, les fondateurs les plus éloignés se retrouvent, de façon contradictoire, dans l’ascendance de presque tous les individus composant la population, et ils sont parfois représentés chez certains avec des probabilités assez élevées. Dans un contexte de peuplement, les premiers arrivants migrent souvent en famille, avec des enfants adultes en âge de s’établir dans le nouveau milieu, donc de se marier et de se reproduire. Ceux qui arrivent seuls contractent des unions avec des membres de la population déjà établis, si bien que chaque nouvelle naissance contribue à la représentation biologique, à la fois du nouvel arrivant et de ceux qui l’ont précédé en termes de migration (les parents de son conjoint). Les arrivants plus récents ont, par ailleurs, une probabilité plus élevée d’être représentés dans la population. Une fécondité différentielle des immigrants peut aussi infléchir l’évolution du patrimoine génétique (Chapman et Jacquard, 1971). Le mode de peuplement (couples immigrants, individus seuls, apparentés ou non apparenté aux premiers), la taille des effectifs impliqués, le degré d’ouverture de la population, les choix préférentiels de conjoint ou encore l’absence de choix sont autant de facteurs qui modulent la représentation biologique des fondateurs. Par exemple, l’existence et le maintien de liens entre une population donnée et la population d’origine peuvent renforcer la représentation génétique des fondateurs et contribuer à homogénéiser le bassin génétique de la population observée au lieu de favoriser l’introduction de variants génétiques.
Le peuplement initial ou la migration subséquente d’une paroisse ou d’une région se situent dans un contexte historique, économique et culturel qui recouvre plusieurs niveaux de réalité; il faut interpréter l’évolution biologique de la population en tenant compte de ces facteurs et de l’échelle à laquelle ils opèrent. À ce jour, les études de démographie historique ou génétique sur le Québec ont concerné la population du Québec dans son ensemble ou encore la région du Saguenay. Elles ont porté, notamment, sur l’identification des mécanismes qui pourraient expliquer la constitution de pools géniques régionaux et (par exemple) les effets fondateurs en relation avec la distribution régionale différentielle des pathologies héréditaires (Tremblay et al., 2001; Gagnon et Heyer, 2001; Gagnon et al., 1998; Bouchard et al., 1995; Heyer et Tremblay, 1995). Dans tous ces travaux, c’est un ensemble divisé en régions qui était observé, alors que le présent travail porte sur une petite population; l’observation est donc faite à une échelle beaucoup plus réduite, et la perspective est différente et complémentaire. Une étude intragénérationnelle des alliances conjugales et plus particulièrement de l’endogamie et des unions consanguines à partir de huit micro-régions du Saguenay a mis en relief l’utilité d’une analyse plus fine à l’échelle des familles et de la paroisse pour faire apparaître des variations qui sont silencieuses à cause de l’échelle utilisée (Bouchard, 1989). Au terme d’un examen de l’endogamie intergénérationnelle où ils abordent la question des effets à long terme de l’endogamie régionale sur la diversification des pools géniques de la population du Saguenay (de nouveau), les auteurs s’interrogent sur le rôle éventuel de l’endogamie familiale et sur son impact sur la consanguinité régionale (Tremblay et al., 2000). Un travail plus récent (Vézina et al., 2004) sur les mesures de l’apparentement et de la consanguinité au Saguenay-Lac-Saint-Jean précise qu’il faudrait, pour l’étude de certains phénomènes, travailler à une échelle plus fine, comme celle des sous-régions ou même des villages ou des localités.
Objectifs
C’est en vue de cerner la contribution différentielle des immigrants fondateurs d’une population que nous évaluons le rôle de la parenté biologique dans le mouvement de peuplement, afin de voir si celle-ci ne vient pas ajouter un poids non négligeable à leur représentation biologique. Plus spécifiquement, il s’agit de vérifier si le comportement matrimonial et reproducteur des descendants des fondateurs dilue ou intensifie la représentation biologique de ces derniers au fil des générations, à quel moment le maintien des liens avec les régions d’origine s’atténue et quelle est la dynamique plus fine qui nourrit ces échanges. Encore au milieu du 19e siècle, des liens privilégiés existaient toujours entre les habitants de l’Isle-aux-Coudres et ceux de la région de Charlevoix (Mayer et Boisvert, 2005). Des études sur le peuplement de la région du Saguenay-Lac-Saint-Jean ont montré l’existence de liens de parenté entre les émigrants en partance de la région de Charlevoix (voir en particulier Jetté et al., 1991); à une échelle plus petite, le même phénomène a été observé, notamment, à Bois-Vert au Saguenay (Gomila et Guyon, 1969) et à La Serpentine dans les Cantons de l’Est (Mayer, 1977).
Population étudiée
L’Isle-aux-Coudres, dans la région de Charlevoix, a fait l’objet de recherches dans diverses disciplines, dont celle de l’anthropologie biologique (Laberge, 1967; Philippe et Gomila, 1971, 1972; Boisvert, 1992; Boisvert et Mayer, 1994), et son histoire est assez bien connue et documentée (Lalancette, l980; Dépatie et al., 1987; Desjardins-MacGregor, 1992). Connue pour son endogamie, l’île appartient, de plus, à une région où la consanguinité et la parenté sont relativement élevées si on tient compte des liens éloignés (Vézina et al., 2004). Le coefficient moyen de parenté à l’Isle-aux-Coudres a été estimé à 0,012 et le coefficient moyen de consanguinité à 0,015 (Philippe, 1969). Ces valeurs sont un peu plus élevées que celles qui ont été estimées pour la région de Charlevoix, malgré une profondeur généalogique légèrement moindre, et en dépit de l’évitement de mariages entre proches apparentés : on compte 12 mariages entre cousins germains sur un total de 1351 mariages célébrés à l’île (Boisvert et Mayer, 1994).
Le peuplement initial de l’Isle-aux-Coudres
Située dans l’estuaire du fleuve Saint-Laurent, au nord-est de Québec et à moins de trois kilomètres de la côte charlevoisienne, face à Baie-Saint-Paul et aux Éboulements, l’Isle-aux-Coudres couvre une superficie de cinq kilomètres sur onze et compte trois côtes : la Côte à la Branche (ouest), la Côte de la Baleine, au sud-est, et une troisième, septentrionale, incluant la Côte des Roches. Martin (1957) et Lalancette (1980) distinguent trois phases de peuplement de l’île :
une phase de peuplement par immigration (1720-1770), au cours de laquelle la population passe de 30 à 213 habitants; cette phase peut être scindée en deux périodes : 1720-1728 (peuplement de type familial) et 1729-1770 (peuplement à caractère individuel);
une phase de peuplement autonome (1770-1790), caractérisée par une baisse de l’immigration et le maintien d’un fort taux d’accroissement naturel, durant laquelle la population s’élève à 566 habitants;
enfin, une troisième phase (1790-1870), de stabilisation des effectifs à un peu plus de 600 habitants (tableau 1). Au début de cette troisième phase, les mariages endogames constituent 35 pour cent des mariages célébrés dans l’île, alors qu’ils en forment 75 pour cent en 1870. En 1960, leur part atteint 82 pour cent (Philippe, 1969).
Lalancette a retracé, grâce à diverses sources (recensements, aveux et dénombrements, papiers-terriers, actes notariés, registres paroissiaux, ouvrages historiques...), l’évolution de l’occupation du sol et de la distribution des terres. Il conclut de son étude que le noyau initial de peuplement est constitué d’apparentés ou d’affins qui consolident leurs liens de telle manière que peu d’étrangers réussissent à s’intégrer à la communauté et à accéder à la propriété, sinon par le biais de mariages avec des insulaires. Ces mariages exogames deviennent inévitables dans ce contexte et constituent le seul type d’immigration observé (Lalancette, 1980).
Tableau 1
Évolution de la population de l’Isle-aux-Coudres (1721-1967)

Endogamie et choix de conjoint
Le choix limité de conjoints, compte tenu du faible effectif de mariables, a amené les insulaires, au début du peuplement, à épouser des conjoints venus de l’extérieur, d’autant plus que l’Église décourageait la célébration d’unions entre proches parents. Avec l’accroissement naturel de la population, les unions endogames sont devenues majoritaires et, dès 1890, la moitié d’entre elles étaient consanguines; durant la période 1910-1929, leur part passe de 75 pour cent à 98 pour cent. La consanguinité serait principalement due à la dérive génétique [2], et la plupart des unions auraient été célébrées au hasard, avec un évitement pour les unions entre proches cousins (Philippe, 1969). Quant aux unions exogames, elles se sont surtout produites dans les décennies qui ont suivi le peuplement initial. L’existence de dispenses religieuses pour un certain nombre de ces unions indiquerait un certain choix préférentiel dans la parenté, choix que nous ne pouvons estimer que partiellement, puisque les unions célébrées dans la paroisse de l’épouse nous échappent. Il peut aussi s’agir d’un apparentement correspondant à l’apparentement moyen que les gens de la côte et de l’île partagent, vu leur origine commune, et qui peut nécessiter une dispense de l’Église [3].
L’approche de population comporte donc une limite qui ne peut qu’en partie être compensée par la consultation de bases de données régionales et la reconstitution généalogique, et elle mène fort probablement à une sous-estimation de l’apparentement biologique réel entre les individus de la communauté ou encore de leur consanguinité. Étant donné l’existence d’actes de mariages célébrés à l’île dans lesquels l’épouse est résidente de l’île et qui inscrivent une dispense religieuse, on peut penser, pour le moins, qu’un certain nombre des unions célébrées dans la paroisse de l’épouse exogame ont nécessité une dispense, et que nos résultats d’analyse présentent donc une estimation minimum de la perte de variabilité du bassin génétique. Par contre, notre approche permet de suivre à une échelle plus fine l’évolution démographique et biologique de la communauté et aide à mieux comprendre comment les différents pools géniques se sont formés au Québec.
Registre de population et enquête ethnologique
Cette étude utilise le Registre de la population de l’Isle-aux-Coudres, constitué à partir du dépouillement des actes d’état civil (baptêmes, mariages et sépultures) et de leur jumelage selon la méthode de reconstitution des familles (Fleury et Henry, 1965); il couvre la période qui s’étend du début du peuplement à 1973 (Philippe, 1971; Boisvert, 1992). Le Registre comprend 8382 individus et 2001 unions, identifiés grâce au rapprochement des mentions nominatives contenues dans les 6740 actes de naissance, 1351 actes de mariage et 2487 actes de décès enregistrés à l’état civil. Est aussi utilisé le recensement ethnologique de 1967 (Philippe, 1969), qui sert de base pour définir le pool génique le plus récent à partir duquel seront observées les contributions génétiques des fondateurs chez leurs descendants.
Limites et avantages du Registre
Le travail de Lalancette confirme l’existence de liens très étroits entre les premiers concessionnaires et habitants de l’île. Le Registre de population nous permet, en partie, de retracer cette information pour ceux-ci et les autres qui ont migré par la suite à l’île. On ne peut connaître directement l’apparentement des couples immigrants déjà mariés, en remontant les ascendances ou en consultant les dispenses, puisque les données sur leur mariage sont absentes du Registre; il est cependant connu indirectement par les dispenses de parenté octroyées lors du mariage de leurs descendants à l’île, mais de façon incomplète, puisque certains apparentements concernent en partie des branches de la famille extérieures à l’île, donc non inscrites dans le fichier de population. Par définition le Registre de la population ne fournit d’information que sur l’ensemble des individus qui, à un moment ou l’autre de leur vie, ont été sujets d’acte d’état civil ou mentionnés dans un acte en raison d’un lien conjugal ou de procréation avec le sujet. Il permet de remonter, par liens générationnels, à toutes les fins d’information généalogique, qui concernent par définition des couples. Fait plus intéressant, le Registre permet de distinguer, parmi l’ensemble des immigrants ou conjoints exogames, ceux qui ont fait souche (les immigrants fondateurs) de ceux qui n’ont pas eu de descendants à l’île ou n’en ont eu que pour une période non continue. Il permet aussi, pour les unions célébrées à l’île, d’estimer à l’aide des dispenses ou de calculer à l’aide des généalogies le coefficient de parenté entre les conjoints.
Ce fichier permet aussi de retracer des bribes de mouvements d’aller-retour à l’île ou des passages probablement brefs, qui montrent que le Registre n’est pas parfait dès qu’il s’agit d’étudier la migration. La consultation des recensements (Boisvert, 1992) a confirmé cette lacune et mis en évidence que les recensements peuvent aider à cerner de plus près le phénomène tout en étant une source lacunaire aussi sur ce point. Étant donné les liens organiques entretenus par les résidents de l’île avec ceux de la région de Charlevoix (Collard, 1985, 1999; Mayer et Boisvert, 2005), nous avons essayé de repousser les limites du Registre en prenant systématiquement en considération les mariages célébrés dans Charlevoix qui impliquaient au moins un résident de l’île [4]. Cela nous a permis de retracer 46 mariages, dont 11 ont nécessité une dispense de l’Église, c’est-à-dire près du quart, et pour des liens de type 3-4 et 4-4 [5]. Ces unions ne sont pas prises en considération ici, étant donné la démarche méthodologique adoptée pour la présente étude.
Définitions, concepts et mesures
Fin d’information généalogique et fondateur
Un certain nombre de concepts et de mesures doivent être présentés puisqu’ils sous-tendent la structuration des données et leur analyse. En outre, certains concepts ou termes connus, tels « fondateur », « fin d’information généalogique » et « contribution génétique », peuvent, selon les disciplines, correspondre à des notions différentes. Dans une approche de population, les limites de l’observation sont à la fois imposées par le Registre et définies par le chercheur. Dans les paroisses de peuplement, les limites inhérentes au Registre sont, d’une part, l’ouverture du registre paroissial, qui suit de peu l’installation des pionniers, et, d’autre part, l’arrêt du dépouillement des registres d’état civil à une date définie, plus ou moins récente. En recherche, les « fins d’information généalogique » identifient les fondateurs de la population, dont la contribution génétique sera suivie le long des descendances pour être observée à un moment précis dans le temps. L’intervalle d’observation pourra varier, par ailleurs, selon les fins des analyses; ainsi, on utilisera un découpage par regroupement de généalogies, pour éviter le plus possible leur chevauchement (Jacquard, 1972), ou un découpage transversal selon des critères démographiques prédéfinis (Mayer, 1977), par exemple en recourant à un recensement nominatif (Mayer, 1977; Charbonneau et al., 1987; Asselin, 2003); ou encore on reconstituera, à l’aide des généalogies, des pools géniques successifs (Gagnon et al., 2004).
En génétique des populations, le fondateur constitue donc la fin de l’information généalogique (Jacquard, 1972). « Les fondateurs sont les ancêtres dont l’ascendance est inconnue; ils représentent pour l’historien les extrémités des généalogies qu’il a pu reconstituer, pour le généticien les sources des gènes présents dans le groupe » (Jacquard, 1977). En histoire, la reconstitution des généalogies peut dépasser largement les limites de la population étudiée afin de répondre à diverses problématiques de recherche; le travail de Dessureault (2001) sur le village de Lavaltrie en est une illustration. On peut aussi considérer comme fondateurs d’un pays, d’une région, d’un village, les individus qui y ont immigré sans pour autant contribuer à la reproduction biologique de la population. Pour le généticien, ce sont les limites mêmes de la population qui déterminent le cadre de sa recherche puisqu’elles définissent l’unité de reproduction au sein de laquelle se fera l’évolution des structures génétiques. Les fondateurs « représentent la source, multiple, des gènes actuels, telle que notre connaissance des filiations permet de la décrire » (Jacquard, 1977 : 76). « Ils peuvent appartenir au petit groupe initial qui a effectivement fondé la population ou faire partie des immigrés qui sont venus s’y joindre ultérieurement » (Jacquard, 1972). Dans l’étude sur les Français établis au Canada au 17e siècle, les fondateurs pionniers sont aussi appelés « tête de lignée » (Charbonneau et al., 1987 : 120). Selon les disciplines, le terme « fondateur » peut donc être interprété de plusieurs façons; dans le présent travail, le terme « fin d’information généalogique » a été préféré à celui de « fondateur » puisqu’il renvoie sans ambiguïté à la méthode utilisée pour identifier l’individu dont la contribution génétique sera mesurée dans sa descendance.
Estimations probabilistes de la parenté biologique et de la consanguinité
En génétique des populations, les coefficients de parenté biologique et de consanguinité permettent de mesurer de façon probabiliste la perte de variabilité génétique de la population.
Le coefficient de parenté entre deux individus A et B est défini par Jacquard comme étant « la probabilité pour qu’un gène désigné au hasard chez un individu et un gène désigné au hasard, au même locus, chez l’autre, soient identiques ». La formule utilisée est celle de Malécot (1948) :
Le coefficient de consanguinité d’un individu Z est défini comme étant « la probabilité pour que les deux gènes qu’un individu possède en un locus soient identiques, c’est-à-dire soient la copie d’un même gène ancêtre » (Jacquard, 1974). Il peut être calculé selon la formule de Wright (1922) :
La valeur du coefficient de consanguinité d’un individu est égale à la valeur du coefficient de parenté de son père et de sa mère.
La dispense religieuse et la mesure de parenté
Lorsque la profondeur généalogique est insuffisante pour permettre un calcul des coefficients de parenté entre les conjoints, les dispenses religieuses offrent la possibilité d’identifier des liens de parenté. Il faut, par ailleurs, tenir compte du fait que l’interdit religieux a progressivement diminué en importance, de telle sorte que les empêchements ont été abaissés au quatrième degré égal (consanguinité 4-4) lors du concile de Latran, en 1215, puis au troisième degré égal (consanguinité 3-3) en vertu du Code de droit canonique promulgué en mai 1917 et entré en vigueur en mai 1918, et enfin au deuxième degré (consanguinité 2-2, c’est-à-dire entre cousins germains) en vertu du Code de droit canonique révisé en 1983. On calcule le degré de consanguinité en comptant le nombre de chaînons qui séparent chacun des conjoints de l’ancêtre qu’ils ont en commun. Ainsi, une consanguinité 4-4 signifie que 4 chaînons séparent chacun des conjoints de l’ancêtre commun, et ainsi de suite. La dispense peut donc être transformée en valeur de coefficient de parenté. Nous avons vérifié la fiabilité des dispenses inscrites dans les registres de l’île en confrontant les valeurs estimées à partir des dispenses et les valeurs calculées des coefficients de parenté, pour les couples dont la dispense indiquait une parenté biologique et pour lesquels nous étions en mesure de calculer le coefficient de parenté (Boisvert, 1992; Boisvert et Mayer, 1994).
La probabilité d’origine des gènes
La probabilité d’origine des gènes (Jacquard, 1972) met l’accent sur la variation génétique puisqu’elle mesure les contributions des fondateurs de la population aux différents pools géniques. Jacquard explique qu’en reconstituant ainsi les généalogies par leurs bases, avec la fin de l’information généalogique, on retrace les fondateurs du groupe; en fait, selon Jacquard (1974), l’intérêt de remonter les généalogies n’existe que pour la reconstitution de l’évolution du patrimoine biologique d’une population. Ainsi, en partant, par exemple, d’un recensement nominatif, on remonte les ascendances de tous les individus recensés jusqu’aux fondateurs des lignées; il s’agit ensuite de calculer la contribution génétique de ces derniers dans la population observée en prenant en compte le nombre de descendants, les liens qui relient les parents de ces individus à chaque fondateur et la profondeur généalogique à laquelle ce dernier se situe.
La probabilité d’origine des gènes (POG) est « la probabilité qu’un gène pris au hasard chez un fondateur se retrouve chez les individus de la population étudiée » (Jacquard, 1972). Elle peut être calculée selon la formule suivante (Heyer, 1999) :
Comme il s’agit d’un calcul de probabilité, le total des contributions ne peut dépasser l’unité.
Ajoutons, à la suite de Chapman et Jacquard (1971 : 178), que les « probabilités d’origine des gènes ainsi calculées ne représentent pas la même réalité que la notion, souvent évoquée, de “nombre de descendants” de divers fondateurs ». En effet, les « réseaux d’ascendance paternel et maternel d’un individu ont naturellement, si l’on remonte assez loin dans le passé, des parties communes; un même ancêtre peut être atteint par plusieurs voies : pour calculer les probabilités d’origine le concernant, il suffit alors d’additionner les probabilités affectées à chacune des voies qui aboutissent à lui » (Jacquard, 1972 : 790). Cette mesure de la contribution des fondateurs au patrimoine génétique de la population tient donc compte à la fois du nombre de descendants, du nombre de voies rattachant les descendants à chaque fondateur et finalement du nombre de générations qui les sépare. Elle est plus exacte, par exemple, que la mesure utilisée pour l’étude de la contribution génétique des pionniers français établis au Canada au 17e siècle (Charbonneau et al., 1987), qui, elle, ne tient compte que du nombre de descendants par génération, pondéré par la probabilité de transmission à chaque génération, et laisse de côté la multiplication des liens qui relient les descendants à l’ancêtre fondateur à travers divers ancêtres situés dans l’ascendance; cette méthode ne donne pas une idée juste de la véritable contribution probabiliste des fondateurs. Plus la population est endogame, de petite taille et observée sur un grand nombre de générations, plus les liens ont le temps de se complexifier et moins on peut négliger de prendre en compte, dans le calcul de la contribution génétique, le nombre de chemins reliant un individu à un ancêtre fondateur. Ainsi, plus les descendants d’un fondateur se reproduiront à l’intérieur de la même lignée, plus les liens reliant l’ancêtre à ses descendants seront nombreux et augmenteront, du moins en probabilité, la représentation biologique de l’ancêtre chez ses descendants. Ces liens témoigneront éventuellement de la subdivision de la population en plus petites unités endogames (Mayer et al., 1985).
Les couples « fin d’information généalogique »
Définition
En génétique des populations, une « fin d’information généalogique » coïncide obligatoirement avec un fondateur qui est un géniteur. Les fins d’information généalogique sont par définition des couples repérés dans la base de données selon différents événements démographiques de manière à ce qu’on puisse les dater, minimalement dans certains cas, et surtout les classer selon leur statut d’entrée à l’île. Ainsi, dans un Registre de population, à quelques rares exceptions près, la fin d’information généalogique en ascendance est toujours le couple : au baptême, au décès ou au mariage d’un enfant, les parents sont nommés. Au décès d’un conjoint, c’est directement le couple qui est identifié, comme dans le cas de certains mariages, par exemple celui d’un veuf et d’une veuve.
Repérage chronologique
L’événement qui situe le mieux chronologiquement le couple est son mariage. Nous avons distribué toutes les fins d’information en fonction de la présence ou de l’absence d’une date complète de mariage à l’île le caractérisant; ces deux classes de fin d’information ont ensuite été subdivisées selon un autre événement démographique : première naissance ou premier décès d’un enfant à l’île, ou premier mariage d’un enfant célébré à l’île ou, en dernier ressort, décès d’un membre du couple. Ensuite, nous avons examiné la fiche de famille de chaque couple pour mieux documenter les cas complexes et, ultérieurement, classer chaque couple fin d’information selon son statut d’entrée dans la population.
Unions sans date de mariage
Les unions sans date de mariage et sans aucune autre date d’événement démographique les concernant sont presque exclusivement des premières unions de veufs ou de veuves qui figurent dans le Registre à cause de remariages à l’île. Les unions sans date de mariage et repérées grâce à une première naissance d’enfant à l’île concernent des couples venus à l’île avec une famille habituellement en partie constituée. Il peut aussi s’agir de couples dont un des membres est un résident de l’île (habituellement l’homme) et dont le mariage a été célébré hors de l’île, c’est-à-dire dans la paroisse de l’épouse, selon la coutume. Le couple réside à l’île et y élève sa famille (exogamie avec gain). Les couples identifiés par un décès d’enfant n’ont habituellement pas eu un autre enfant à l’île sauf quelques exceptions. Dans près de la moitié des cas, il s’agit de décès de célibataires adultes dont les parents ne semblent pas avoir résidé à l’Isle-aux-Coudres puisque aucun autre événement démographique ne les caractérise. Les couples identifiés par mariage d’enfants sont habituellement des couples parents d’un enfant venu se marier dans la paroisse de son épouse; le couple peut s’établir à l’île ou résider ailleurs. Les couples identifiés par le décès d’un des conjoints sont des couples dont un membre ou les deux décèdent à l’île; ils sont habituellement âgés et sans liens de filiation ou de procréation. Il peut s’agir de parents d’un individu ayant immigré déjà marié, ou de parents de célibataires n’étant pas nés ou décédés à l’île.
Dans cette catégorie, certaines unions ne peuvent donc être datées alors que les autres le sont grâce à des événements démographiques entre lesquels peut exister un écart équivalent à une génération (par exemple, naissance et mariage d’enfant). Le hiatus entre deux générations est réel et pourrait, en partie, être confirmé par les données sur les unions célébrées en dehors des deux paroisses de l’île.
Unions avec date complète de mariage
Bon nombre de couples d’abord repérés dans le Registre de population par la date de leur mariage le sont ensuite par une date de naissance d’un premier enfant né à l’île ou, parfois, de décès d’un enfant (dans ce dernier cas, c’est souvent le seul indice de la présence des parents durant un bref séjour à l’île). L’autre groupe, dont les effectifs sont encore plus nombreux, est représenté par les unions exogames avec perte et, dans ce cas, aucun autre événement démographique ne suit celui du mariage. Lorsque les deux époux sont nés hors de l’île, ils appartiennent souvent à une famille migrante avec laquelle ils sont venus à l’île. Les autres catégories sont peu fréquentes.
Individus sans union
À quelques rares exceptions près, comme nous l’avons mentionné précédemment, la fin d’information généalogique en ascendance est le couple. On repère les rares exceptions en examinant les individus sans mention de parent et sans union. Sur 8382 individus répertoriés dans le Registre de population, 1100 figurent sans mention de parents et seulement 68 d’entre eux ne sont pas sujets d’union. Il s’agit de religieux, d’enfants adoptifs, d’individus noyés sans mention de parents ou encore d’individus qui sont décédés durant une période d’épidémie et dont l’acte de sépulture est très sommaire. D’autres sont connus exclusivement par l’enquête ethnologique menée par Pierre Philippe en 1967 : il s’agit d’immigrants qui ont pu s’établir ultérieurement dans la population mais échappent à notre observation.
Types de « fin d’information » généalogique : couple immigrant, couple non immigrant
L’examen des types de couples selon les différentes catégories de repérage chronologique par premier événement démographique nous a permis de réaliser qu’il n’y a pas nécessairement adéquation entre couple « fin d’information » et couple immigrant, et que chaque union devait faire l’objet d’un examen attentif. Certains couples « fin d’information » pouvaient être assimilés à des immigrants constitués en couple venu s’établir à l’île dans le cadre d’une migration familiale, ou venus individuellement s’y établir, s’y marier et y résider ou non. Par contre, un enfant ayant immigré avec sa famille et s’étant marié subséquemment à l’île ne représente pas une « fin d’information » puisque ce sont ses parents qui le sont, mais il demeure un immigrant. Pour complexifier notre observation, ajoutons que la personne qu’il épouse est née à l’île de parents immigrants (figure 1). Enfin, d’autres couples « fin d’information » ne peuvent absolument pas être considérés comme des immigrants puisqu’ils ne sont connus que par leur mention lors du mariage de leur enfant à l’île (non représentés à la figure 1).
Statut à l’entrée dans la population
Pour les couples « fin d’information » généalogique qui sont aussi immigrants (fondateurs), nous avons créé trois statuts à l’entrée dans la population, selon que le couple ou l’homme ou la femme était un immigrant fin d’information généalogique. Ces statuts caractérisent le couple et non pas l’individu. C’est l’examen de la fiche de famille, extraite du Registre de la population, combiné à celui des types de fin d’information décrits plus haut qui a permis de confirmer le statut des couples à l’entrée.
Les parents d’un individu venu se marier à à l’Isle-aux-Coudres mais qui eux-mêmes n’y ont jamais résidé sont exclus de l’analyse. Ils demeurent cependant dans la base de données puisque c’est leur mention qui contribue à déterminer le statut d’immigrant fondateur de leur enfant.
Analyses et résultats
Distribution des unions selon le statut de fondateur
Sur un total de 563 couples comprenant soit deux soit un conjoint fondateur, 37 pour cent (208) sont des couples dont les deux membres sont des fondateurs, 47 pour cent (264) des couples dont l’homme est un fondateur et 16 pour cent (91) des couples dont la femme est une fondatrice (tableau 2, section A). Les couples à deux fondateurs sont plus nombreux dans les premières décennies qui suivent le début du peuplement de l’île; à partir de 1780 les couples dont l’homme est un fondateur sont plus nombreux, et ce jusqu’à la fin de l’observation, sauf entre 1820 et 1899, où ces deux catégories sont en nombre à peu près égal (figure 2).
Figure 1
Couples fin d'information généalogique (FIG), couples immigrants (I)
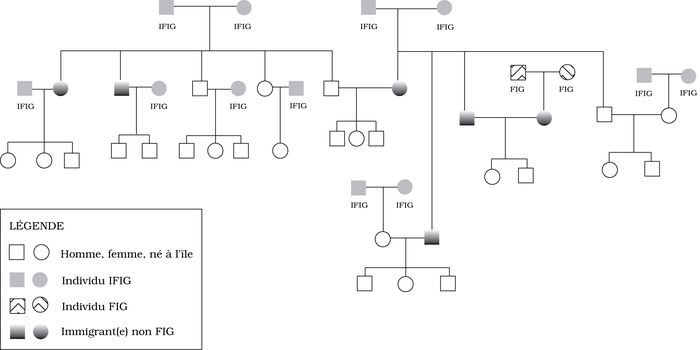
Il est intéressant de constater qu’il y a eu des entrées par migration de couples fondateurs à toutes les périodes de l’histoire de la population de l’Isle-aux-Coudres, et plus particulièrement du début du peuplement au début du 20e siècle. Il faudra examiner les facteurs qui ont joué pour amener des familles et des individus à immigrer à l’île de façon relativement constante. Si un seul des membres du couple est immigrant, on peut légitimement faire l’hypothèse qu’il a pu s’établir à l’île par mariage, ou que le mariage a pu suivre une immigration préalable. Le recensement ethnologique de 1967 a mis en évidence des mouvements d’aller-retour à l’île au niveau d’une génération. Un individu né à Montréal de parents nés à l’île se marie à Montréal et s’établit par la suite avec sa famille dans l’île. L’enquête (seule) a permis de faire ressortir le lien entre l’individu et ses parents et illustré des allées et venues insoupçonnées. On peut faire l’hypothèse que les liens biologiques étaient encore susceptibles d’inciter à la migration au 20e siècle.
Tableau 2
Distribution, par période de 20 ans, des couples fin d’information (A) ou issus de fin d’information (B), selon divers critères de repérage dans le Registre de population, et selon que les conjoints sont nés ou non dans la population a

FIG : fin d’information généalogique.
IFIG : immigrant fin d’information généalogique = fondateur.
Figure 2
Distribution, par période de 20 ans, des couples fin d'information ou issus de fin d'information selon divers critères de repérage dans le Registre de population (conjoints nés ou non dans la population)
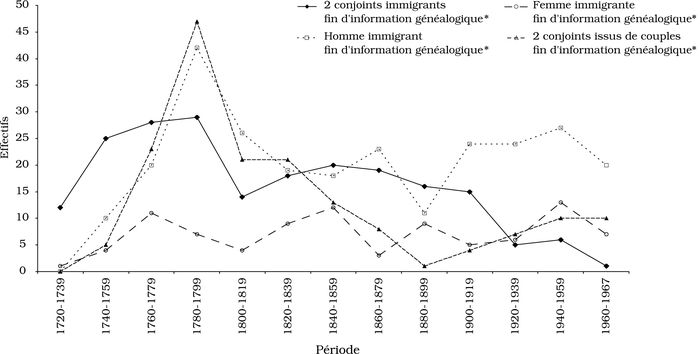
* Immigrant fin d'information généalogique : fondateur.
Sur les 264 unions où l’homme est exogame, la majorité (248/264) a été célébrée à l’île (données non présentées). Il faut faire la part des choses en ce qui concerne l’accroissement du nombre d’unions où l’homme est extérieur à la communauté. Puisque la coutume ecclésiastique exige que l’on se marie dans la paroisse de la jeune fille (il faut demander une autorisation de son curé pour y déroger), plus de la moitié de ces mariages célébrés à l’île (148/248 : 59,7 pour cent) n’a pas été suivie de l’établissement du couple à l’île. La tendance semble aller vers une augmentation de ce type de mariage avec le temps; il est réaliste de penser que souvent, durant la période initiale de peuplement, les couples exogames s’établissaient dans l’île parce que le mariage était la seule façon d’acquérir les terres, qui se transmettaient préférentiellement au sein des familles (Lalancette, 1980).
Les unions où l’épouse est née hors de l’île nous sont connues dans la moitié des cas (42/91) par le fait que les couples mariés dans la paroisse de l’épouse ont résidé à l’Isle-aux-Coudres et y ont élevé leur famille; les 49 autres unions ont été célébrées à l’île. Sur les 51 unions de la période allant du début du peuplement à 1860 environ, 43 impliquent des femmes venues se marier à l’île, mais seulement 6 des 40 unions de la période qui suit ont été célébrées à l’île.
Distribution des unions impliquant les descendants immédiats des couples fondateurs
Le tableau 2 (section B) présente la distribution des unions conclues entre des conjoints qui n’étaient pas des fondateurs mais étaient tous deux issus de parents fondateurs. Précisons qu’un des deux conjoints ou les deux a été immigrant à l’île avec sa famille ou est né de parents immigrants. Ces couples issus de parents fondateurs sont plus nombreux durant la période de peuplement autonome (1770-1790) et un peu après, ce qui les rattache en majorité aux familles immigrantes de la période de peuplement initial (figure 2).
Apparentement et fécondité
La caractérisation des couples en termes d’information généalogique et de migration et l’identification des fondateurs permettent de mesurer l’apparentement biologique des couples fondateurs et des couples constitués par leurs descendants. Cet apparentement biologique peut avoir eu un impact sur l’homogénéisation du patrimoine génétique de la population dans la mesure où la fécondité [6] s’est exprimée à l’île.
Apparentement des couples fondateurs
La mesure d’apparentement provient des dispenses de l’Église accordées pour parenté biologique aux couples désireux de se marier : logiquement, l’absence de profondeur généalogique ne permet pas de connaître les liens entre les conjoints fondateurs. Les dispenses religieuses n’ont pu être relevées que pour les couples dont le mariage a été célébré à l’Isle-aux-Coudres. Il est toutefois intéressant de constater que pour l’ensemble de la période d’observation, 22 pour cent des couples sont apparentés biologiquement; en outre, la proportion va croissant (tableau 3), des couples où les deux conjoints sont immigrants (13 pour cent) à ceux où l’homme est immigrant (22 pour cent) puis à ceux où c’est la femme qui immigre (27 pour cent). Peut-on voir là l’expression de choix préférentiels de conjoint se concrétisant par des mariages avec des apparentés qui ne sont pas de l’Isle-aux-Coudres ? L’absence de conjoint potentiel a pu prévenir l’endogamie au début du peuplement, et la croissance de la population élargir le marché matrimonial local dans les périodes ultérieures, mais il demeure que près du quart des unions exogames célébrées à l’île ont nécessité une dispense.
Apparentement des couples issus de parents fondateurs
Les couples formés de conjoints non fondateurs mais qui sont venus à l’île avec leur famille ou qui sont nés de ces couples immigrants fondateurs sont beaucoup plus souvent apparentés (58 pour cent). Les chiffres confirment l’existence d’un comportement matrimonial endogame dès la génération issue des premières familles immigrantes, comportement qui s’est maintenu au cours de l’histoire de l’île (Philippe, 1969, 1971). La parenté semble être un moteur d’intégration sociale extrêmement important dans cette population. Les études ethnologiques de Collard (1985, 1991, 1999) sur une population de la côte de Charlevoix, face à l’Isle-aux-Coudres, mettent bien en évidence l’importance de la parenté comme institution sociale majeure et facteur d’intégration et d’exclusion, de même que les échanges entre les grandes familles patronymiques.
Dans le cadre d’une étude sur les individus déclarés non membres de la famille au recensement de l’Isle-aux-Coudres de 1851 (Mayer et Boisvert, 2005), nous avons observé qu’un certain nombre de jeunes adultes résidant dans des ménages de l’île étaient en fait apparentés au chef de ménage ou à son épouse et que certains d’entre eux ont par la suite pris conjoint dans la population et y ont élevé leur famille. Un certain nombre de ces jeunes adultes, bien que nés à l’extérieur de l’île, avaient des parents ou des grands-parents originaires de l’Isle-aux-Coudres. Parfois les liens de parenté sont un peu plus éloignés. La consultation du Registre de la population ainsi que des recensements et des registres de leur paroisse d’origine a permis, en reconstituant sur une courte période l’ascendance de ces individus déclarés non membres, de le vérifier.
Individus fondateurs
Les 563 couples fondateurs (tableau 2) comprennent en fait 771 personnes fondatrices (208 couples à deux fondateurs; 264 + 91 couples à un fondateur), et ce pour l’ensemble de la période qui va de 1720 à 1967. Certaines personnes (155) sont venues se marier à l’île sans s’y établir (unions exogames avec perte). Le départ, par mariage, de l’individu né à l’île ou immigré avec sa famille diminue la représentation génétique de ses parents dans le pool génique de la population tout en privant ce même pool génique d’une nouvelle contribution, celle de son conjoint.
Unions célébrées à l’île et comportement reproducteur
On peut examiner le comportement reproducteur des fondateurs efficaces (qui ont effectivement contribué au pool génique) et de leurs descendants, dans un premier temps, en comparant la distribution des mariages célébrés à l’île (tableau 4) avec celle de ces unions qui ont été fécondes dans la population selon le critère d’au moins une naissance d’enfant à l’île (tableau 5). Le nombre de couples mariés à l’île dont les deux conjoints sont nés à l’extérieur est plus élevé au début de la période de peuplement à l’île, et 50,6 pour cent (44/87) d’entre eux ont été féconds à un moment ou l’autre dans la population. Les unions exogames, plus nombreuses à partir de 1760, maintiennent leurs effectifs relativement élevés, surtout dans le cas où c’est l’époux qui est né à l’extérieur. C’est 36,8 pour cent (102/177) de ces mariages exogames où l’homme est né à l’extérieur de l’île qui ont été féconds, alors que 77,8 pour cent (63/81) des unions où l’épouse est née à l’extérieur l’ont été. Les unions entre deux conjoints nés à l’île voient leur nombre augmenter; ces unions endogames sont à 80 pour cent fécondes dans la population (726/906).
Tableau 3
Répartition des unions célébrées à l’Isle-aux-Coudres selon que les conjoints sont apparentés a ou non et selon que les conjoints sont nés ou non dans la communauté

Apparentement calculé ou déclaré (dispenses religieuses).
Unions célébrées à l’extérieur et comportement reproducteur
Il est intéressant de prendre aussi en considération les unions qui n’ont pas été célébrées à l’île mais qui impliquent une résidence à l’île confirmée dans la base de données par la naissance d’enfants. Ces unions sont donc repérées parce que fécondes et nous ne pouvons pas présentement connaître le dénominateur sur lequel les rapporter. Elles ont été distribuées selon les mêmes critères de naissance du conjoint dans l’île (tableau 6). En fait ce sont les couples féconds à l’île dont les deux membres sont nés à l’extérieur de l’île qui sont les plus représentés dans ce groupe (66,5 pour cent : 131/197).
Tableau 4
Distribution, par période de 20 ans, des mariages célébrés à l’Isle-aux-Coudres (1720-1967), selon que les conjoints sont nés ou non dans la communauté

En termes biologiques, ce sont ces couples ainsi que ceux dont un des membres est né hors de l’île qui représentent théoriquement une entrée, dans la population, de gènes provenant de l’extérieur susceptibles de différer en termes alléliques de ceux qui sont déjà en place, quelle que soit la période d’entrée dans la population.
Il faut rappeler que les immigrants féconds qui se sont établis au début du peuplement ont une longueur d’avance sur les immigrants arrivés plus tardivement dans la représentation de leurs gènes aux générations successives. Si de plus leurs descendants se marient entre eux, ou encore s’ils choisissent de se marier dans la parenté, même si elle n’est pas de l’île, le pool génique devient alors restreint du point de vue de la variation allélique. Cette perte de variabilité peut être mesurée par le degré de consanguinité de la population, estimé par l’évolution du degré d’apparentement entre les conjoints au cours du temps. La question qu’il faut maintenant se poser est celle de savoir si, sur l’ensemble des unions fécondes à l’île, ce sont celles qui impliquent des apparentés qui sont les plus fréquentes, ou le contraire.
Tableau 5
Distribution, par période de 20 ans, des unions fécondes célébrées à l’Isle-aux-Coudres (1720-1967), selon que les conjoints sont nés ou non dans la communauté

Le Registre nous limite à l’étude des couples féconds mariés à l’île puisque les dispenses religieuses des couples non mariés à l’île nous échappent.
Apparentement et comportement reproducteur
La mesure probabiliste de l’apparentement biologique (coefficient de Wright, 1922) est calculée à partir des généalogies ascendantes reconstituées. Cet apparentement peut être proche ou éloigné ou les deux à la fois. Lorsque la profondeur généalogique est absente, onn retient la dispense religieuse pour son estimation. Cependant, nous avons basé la comparaison des couples féconds apparentés et non apparentés sur le fait qu’il y avait ou non présence d’un apparentement, quelle qu’en soit la valeur.
Tableau 6
Distribution, par période de 20 ans, des unions sans date de mariage connues par naissance d’enfant, selon que les conjoints sont nés ou non dans la communauté

Apparentement
Quand on examine la présence de parenté biologique entre les couples mariés à l’île, tant pour l’ensemble des unions que pour les seules unions fécondes, on voit que 67 pour cent (908/1351) et 75 pour cent (701/935) concernent des mariages entre apparentés (tableaux 4 et 7 ainsi que tableaux 5 et 8). Selon que les deux conjoints sont nés à l’extérieur ou seulement l’époux ou l’épouse, la fréquence de l’apparentement est respectivement, pour l’ensemble des unions, de 29 pour cent, 26 pour cent et 44 pour cent, et il en est de même pour les unions fécondes : 27 pour cent, 34 pour cent et 46 pour cent. Quand les deux individus sont nés à l’île, que l’on considère l’ensemble des unions célébrées à l’île ou celles qui ont été fécondes, l’apparentement est le même : 86 pour cent (776/906 et 625/726 respectivement).
Ajoutons que ces unions entre apparentés (totales et fécondes) augmentent avec le temps et que ce sont les unions endogames qui expliquent les distributions. Ces résultats appuient les études antérieures (Philippe et Gomila, 1971; Philippe, 1971; Boisvert et Mayer, 1994).
Comportement reproducteur
Lorsque l’on considère, d’une part, la fécondité des couples mariés à l’île (tableaux 4 et 5, résumés au tableau 9A) et, d’autre part, celle des couples apparentés mariés à l’île (tableaux 7 et 8, résumés au tableau 9B), on constate que le pourcentage d’unions fécondes chez les uns et les autres varie selon la composition du couple. Par ailleurs, si on compare ces mêmes couples selon leur composition, on observe les faits suivants : lorsque les deux conjoints sont nés à l’extérieur, le pourcentage d’unions fécondes est à peu près le même dans les deux groupes (50,6 pour cent et 48,0 pour cent). Dans le cas des unions exogames où l’homme est né à l’extérieur de l’île, les unions entre apparentés sont plus fécondes (49,3 pour cent comparativement à 36,8 pour cent). Lorsque l’épouse est née à l’extérieur les valeurs sont comparables : 80,6 pour cent pour les apparentés et 77,8 pour cent pour l’ensemble des unions. Les unions endogames, c’est-à-dire entre deux conjoints nés à l’île, sont à 80 pour cent fécondes dans les deux cas, ce qui ne surprend pas, étant donné que la majorité des unions endogames sont entre apparentés.
Ces résultats montrent que la parenté biologique a été un paramètre important dans la composition des unions même lorsque l’un des conjoints ou les deux ne sont pas nés à l’île. Dans le cas des unions endogames, il faudra faire la part des parentés proches, qui peuvent être objets de stratégies d’alliance, et de celles qui sont éloignées et moins assujetties à des stratégies, si on ne tient pas compte des pressions sociales du groupe, incitant à se marier dans l’île ou à l’extérieur de l’île par exemple, ou bien dans la parentèle ou hors de celle-ci, comme le démontre Collard pour le village de Rivière Frémiote (1999). La fécondité des unions, quant à elle, varie selon la composition du couple; elle est plus importante dans le cas des unions endogames et des unions où l’épouse n’est pas originaire de l’île. Parmi les unions exogames où l’époux est né à l’extérieur, ce sont les unions célébrées entre deux apparentés qui semblent plus fécondes. On peut imaginer que la parenté avec l’épouse favorise l’établissement du couple à l’île plutôt que dans la paroisse de l’époux.
Tableau 7
Distribution, par période de 20 ans, des mariages entre apparentés célébrés à l’Isle-aux-Coudres (1720-1967), selon que les conjoints sont nés ou non dans la communauté

Tableau 8
Distribution, par période de 20 ans, des unions fécondes entre apparentés célébrées à l’Isle-aux-Coudres (1720-1967), selon que les conjoints sont nés ou non dans la communauté

Représentation différentielle des fondateurs dans le pool génique de la population en 1967
Comment la fécondité des unions a-t-elle contribué à la représentation différentielle des fondateurs ? Parmi les immigrants fondateurs de la population, combien ont contribué aux générations successives et combien sont encore représentés dans le pool génique de la population actuelle ? Quels sont ceux qui ont « imprégné » le pool génique de manière telle que nous les retrouvons invariablement dans l’ascendance de tout individu qui naît dans la population ? Quels sont ceux qui, par leur fécondité et celle de leurs descendants, se sont assuré une plus forte représentation ? C’est la mesure de la probabilité d’origine des gènes (Jacquard, 1972) qui permet de répondre, du moins en partie, à ces questions.
Tableau 9
Pourcentage d’unions fécondes chez les couples mariés (A) et chez les couples apparentés (B) mariés à l’Isle-aux-Coudres (1720-1967), selon que les conjoints sont nés ou non dans la communauté

L’examen du pool génique de la population de l’île en 1967 chez les 15-44 ans, c’est-à-dire ceux qui sont en âge de se reproduire et de contribuer à la génération suivante, permet de confirmer que ce ne sont pas tous les immigrants « fin d’information généalogique » qui sont fondateurs biologiques de la population au moment où s’arrête notre observation.
Pour les 720 individus en âge de se reproduire en 1967, nous remontons à 97 couples fondateurs dont la majorité (79,4 pour cent) est constituée de deux fondateurs. Parmi les 182 individus fondateurs, dont l’arrivée s’échelonne du début du 18e siècle jusqu’à la fin de la période d’observation, 13 sont recensés en 1967. Plus de la moitié sont arrivés avant le 19e siècle (54/97 : 55,6 pour cent), 30,9 pour cent au cours du 19e siècle et 13,4 pour cent après ce siècle.
Ces 97 couples fondateurs appartiennent au groupe initial des 563 unions (tableau 2) comprenant deux ou un immigrant « fin d’information généalogique ». Si on élimine du groupe initial les unions exogames avec perte (563 – 155), ces couples représentent 23,8 pour cent des couples fondateurs immigrants. Si on calcule par nombre d’individus fondateurs, ce sont 182 fondateurs sur 616 (771 – 155) qui constituent les fins d’information « contributrices » au pool génique de 1967 (29,5 pour cent). Il est intéressant de constater que seulement un peu plus du quart des immigrants identifiés dans le Registre de population contribue au patrimoine génétique de la population en âge de se reproduire en 1967.
Sur un total de 1,0 (100 pour cent), représentant l’ensemble des contributions au pool génique, les plus fortes contributions reviennent à une fondatrice (contribution de 0,032) et à un fondateur (contribution de 0,021) (voir le petit tableau de la page suivante). Les contributions individuelles se situent en majorité entre 0,001 et 0,0099 (118/182).
Les contributions les plus élevées sont toutes relatives à des immigrants arrivés au 18e siècle (32/33), sauf pour un immigrant arrivé au tout début du 19e siècle. Sur les 182 fondateurs, 110 appartiennent au 18e siècle (60,4 pour cent). L’ancienneté dans la population, le nombre de descendants féconds, l’âge au mariage et les unions endogames sont autant de facteurs combinés qui déterminent la valeur de la contribution du fondateur.

Les deux exemples qui suivent (tableau 10) illustrent bien le jeu des différentes composantes de la valeur de la contribution génétique d’un immigrant fondateur. Ainsi, Madeleine Boissonneault, fondatrice, a épousé vers 1750 Gabriel Dufour, fils de fondateurs. Sa contribution génétique en 1967 est la plus élevée, suivie de celle de Germain Desgagnés, fondateur ayant épousé, autour de 1800, la fille d’un couple de fondateurs.
Pour un nombre égal d’enfants (11), la descendance de la fondatrice est 1,86 fois plus élevée que celle du fondateur (4184 comparativement à 2240); la proportion d’enfants qui se sont reproduits est à peu près la même (25,8 pour cent : 1081/4184, et 23,2 pour cent : 520/2240); une plus grande profondeur généalogique (9 générations comparativement à 7) permet un enchevêtrement plus important des chemins généalogiques entre la fondatrice et ses descendants dans la population, ce qui fait augmenter la représentation génétique de la première chez les seconds.
Dans le pool génique de 1967, sur 720 individus en âge de se reproduire (données non présentées), 657 ont Madeleine Boissonneault comme ancêtre fondatrice et ils se rattachent à elle par 2781 chaînes d’ascendance différentes (on compte en moyenne 4,23 chaînes par individu), alors que 454 individus se rattachent à Germain Desgagnés par 621 chaînes différentes (soit 1,37 par individu). Le nombre de chaînes rattachant les descendants à l’ancêtre fondateur est une composante importante de la valeur de la contribution génétique de celui-ci; ces chaînes sont le résultat des comportements matrimoniaux des descendants au fil des générations et ne sont pas négligeables lorsqu’il s’agit de documenter l’homogénéisation du bassin génétique d’une population.
Par ailleurs, l’existence d’une corrélation positive entre la représentation génétique des fondateurs et le nombre d’enfants, ainsi qu’un rapport élevé de masculinité à la naissance dans les familles qui ont composé la descendance des fondateurs, donne à penser qu’un facteur migratoire différentiel a pu jouer un rôle dans la contribution ancestrale (Asselin, 2003). Les nombreux arrangements résidentiels complexes comprenant un chef de ménage marié vivant avec un et même deux fils mariés laissent croire que des stratégies socioéconomiques et culturelles ont pu, au cours des générations, contribuer à moduler la représentation des fondateurs.
Tableau 10
Variables dans la valeur de la contribution génétique de deux fondateurs chez les 720 individus en âge de se reproduire en 1967

Conclusion
L’utilisation des fins d’information généalogique a permis de suivre l’arrivée des fondateurs biologiques de la population de l’Isle-aux-Coudres et ce, au cours de son histoire. L’immigration a été de type familial au début, accompagnée puis suivie d’une migration individuelle que l’on a pu repérer par la formation des unions et la naissance d’enfants dans la population. Les générations ultérieures ont puisé leurs gènes dans ce bassin initial et dans les ajouts subséquents. Mais ces ajouts n’ont pas nécessairement contribué à diversifier le patrimoine génétique de la population, car un bon nombre d’unions exogames étaient célébrées entre apparentés provenant majoritairement de Charlevoix. De plus, les unions impliquant des apparentés semblent avoir eu une fécondité exprimée dans l’île plus importante que les unions entre non apparentés, ce qui a contribué à l’homogénéisation du pool génique. La probabilité d’origine des gènes, tout en laissant voir une représentation différentielle des fondateurs, met bien en évidence que les individus en âge de se reproduire en 1967 descendent d’un noyau restreint de fondateurs.
Parties annexes
Remerciements
Cette recherche a été rendue possible par une subvention du Conseil de recherches en sciences humaines du Canada (1999-2005), octroyée à F. M. Mayer dans le cadre du programme de recherche « Parenté biologique et reproduction différentielle des immigrants fondateurs de la population de l’Isle-aux-Coudres (Québec) aux 18e et 19e siècles ». Nous tenons à remercier les évaluateurs externes pour leurs remarques stimulantes.
Notes
-
[1]
« Fondateur : tout individu situé à la fin d’une information généalogique ascendante, quel que soit le moment de son introduction dans la population » (Jacquard, 1972).
-
[2]
« La dérive génétique : définit les fluctuations aléatoires qui agitent les fréquences alléliques, particulièrement dans les petites populations, et qui sont le résultat d’un échantillonnage aléatoire parmi les gamètes » (Hartl, 1994 : 69-70). « Les générations successives constituent des échantillons aléatoires du pool génique parental […] Les fréquences alléliques peuvent, par hasard, être différentes de celles de la génération précédente. Cette variation résulte de ce que les allèles qui constituent le pool génique d’une génération constituent un échantillon aléatoire du pool génique de la génération parentale » (Ridley, 1997 : 133). « La vitesse à laquelle la dérive génique modifie les fréquences alléliques dépend de l’effectif de la population; plus la population est réduite, plus l’effet de la dérive est important » (Ridley, 1997 : 135).
-
[3]
La sévérité de la règle a diminué avec le temps. Voir, ci-dessous, la section « La dispense religieuse et la mesure de parenté ».
-
[4]
Ces données ont été fournies par le Centre interuniversitaire de recherches sur les populations (SOREP), aujourd’hui Projet BALSAC, de l’Université du Québec à Chicoutimi, dirigé par G. Bouchard.
-
[5]
On calcule le degré de consanguinité en comptant le nombre de chaînons qui séparent chacun des conjoints de l’ancêtre qu’ils ont en commun. Ainsi, une consanguinité 4-4 signifie que 4 chaînons séparent chacun des conjoints de l’ancêtre commun, et ainsi de suite. Voir, ci-dessous, la section « La dispense religieuse et la mesure de parenté ».
-
[6]
Dans le cadre de ce travail, la naissance d’au moins un enfant à l’île est le critère retenu pour attester la fécondité du couple dans la population.
Références bibliographiques
- ASSELIN, G. 2003. Facteurs contribuant à l’homogénéisation du pool génique de la population humaine de l’Île-aux-Coudres à partir de l’étude des contributions génétiques de ses fondateurs. Université du Québec à Montréal, mémoire de maîtrise en biologie, 119 p.
- BOISVERT, M. 1992. Mortalité infantile et consanguinité dans une population isolée du Québec. Université du Québec à Montréal, mémoire de maîtrise en biologie, 253 p.
- BOISVERT, M., et F. M. MAYER. 1994. « Mortalité infantile et consanguinité dans une population endogame du Québec », Population, 3 : 685-724.
- BOUCHARD, G., H. CHARBONNEAU, B. DESJARDINS, É. HEYER et M. TREMBLAY. 1995. « Mobilité géographique et stratification du pool génique canadien-français sous le Régime français », dans Y. LANDRY, J. A. DICKINSON, S. PASLEAU et C. DESAMA, éd. Les chemins de la migration en Belgique et au Québec, XVIIe-XXe siècles, chapitre III, « Migrations et comportement démographique ». Beauport, Publications MNH : 51-59.
- CANADA. Bureau fédéral de la statistique. 1871 à 1961. Recensements canadiens. Ottawa.
- CHAPMAN, A. M., et A. M. JACQUARD. 1971. « Un isolat d’Amérique centrale : les Indiens Jicaques du Honduras », dans Génétique et populations. Hommage à Jean Sutter. INED, Travaux et documents, Cahier 60 : 163-185. Paris, PUF.
- CHARBONNEAU, H., B. DESJARDINS, A. GUILLEMETTE, Y. LANDRY, J. LÉGARÉ et F. NAULT. 1987. Naissance d’une population. Les Français établis au Canada au XVIIe siècle. INED, Travaux et documents, Cahier 118. Paris, PUF, et Montréal, PUM.
- COLLARD, C. 1985. « Parenté et communauté à Rivière-Frémiote 1880-1960 », Anthropologie et Sociétés, 9, 3 : 57-85.
- COLLARD, C. 1991. « Idéologie et pratique de la parenté : de la classification des parents aux stratégies familiales », dans G. BOUCHARD et M. DE BRAEKELEER, éd. Histoire d’un génome. Population et génétique dans l’est du Québec. Québec, Presses de l’Université du Québec : 121-144.
- COLLARD, C. 1999. Une famille, un village, une nation. La parenté dans Charlevoix, 1900-1960. Québec, Les éditions du Boréal, 194 p.
- DÉPATIE, S., M. LALANCETTE et C. DESSUREAULT. 1987. Contributions à l’étude du régime seigneurial canadien. LaSalle, Hurtubise HMH, Cahiers du Québec, Histoire, 292 p.
- DESJARDINS-MACGREGOR, L. 1992. La transmission du patrimoine à l’Isle-aux-Coudres au XVIIIe siècle. Université McMaster, thèse de doctorat (Géographie), 451 p.
- DESSUREAULT, C. 2001. « Parenté et stratification sociale dans une paroisse rurale de la vallée du Saint-Laurent au milieu du XIXe siècle », Revue d’histoire de l’Amérique française, 54, 3 : 411-447.
- FLEURY, M., et L. HENRY. 1965. Nouveau manuel de dépouillement et d’exploitation de l’état civil ancien. Paris, INED, 182 p.
- GAGNON, A., et É. HEYER. 2001. « Fragmentation of the Québec population genetic pool (Canada). Evidence from the genetic contribution of founders per region in the 17th and 18th centuries », American Journal of Physical Anthropology, 114, 1 : 30-41.
- GAGNON, A., H. CHARBONNEAU et B. DESJARDINS. 1998. « Apparentement des conjoints par région au Québec ancien », dans G. BRUNET, A. FAUVE-CHAMOUX et M. ORIS, éd. Le choix du conjoint. Lyon, BOSC, Les chemins de la recherche, 43 : 133-150.
- GAGNON, D., M. LIMA, M. BOISVERT, E. LABELLE et F. M. MAYER. 2004. Application de l’analyse généalogique à diverses problématiques en génétique des populations et en épidémie génétique. Dix-septièmes Entretiens du Centre Jacques Cartier, colloque « Usages et pratiques de la généalogie dans les sciences humaines, sociales et biologiques », Montréal, 7-8 octobre.
- GOMILA, J., et L. GUYON. 1969. « Étude comparative des petites communautés rurales. Méthode et premiers résultats à propos de la consanguinité à Bois-Vert (Québec) », Population, 24, 6 : 1127-1153.
- HARTL, D. 1994. Génétique des populations. Traduit par Nicolas Borot (édition américaine, 1988). Paris, Flammarion, Médecine-Sciences.
- HEYER, É. 1999. « One founder/one gene hypothesis in a new expanding population: Saguenay (Québec, Canada) », Human Biology, 71, 1 : 99-109.
- HEYER, É., et M. TREMBLAY, 1995. « Variability of the genetic contribution of Quebec population founders associated to some deleterious gene », The American Journal of Human Genetics, 56 : 970-978.
- JACQUARD, A. 1972. « Évolution du patrimoine génétique des Kel Kummer », Population, 4-5 : 784-800.
- JACQUARD, A. 1974. Génétique des populations humaines. Paris, PUF, 399 p.
- JACQUARD, A. 1977 Concepts en génétique des populations. Paris, Masson.
- JETTÉ, R., D. GAUVREAU et M. GUÉRIN. 1991. « Aux origines d’une région : le peuplement fondateur de Charlevoix avant 1850 », dans G. BOUCHARD et M. DE BRAEKELEER, éd. Histoire d’un génome. Population et génétique dans l’est du Québec. Québec, Presses de l’Université du Québec : 76-106.
- LABERGE, C. 1967. « La consanguinité des Canadiens français », Population, 22, 5 : 861-896.
- LALANCETTE, M. 1980. La Seigneurie de l’Isle-aux-Coudres au XVIIIe siècle. Université de Montréal, mémoire de maîtrise (Histoire), 224 p.
- MALÉCOT, G. 1948. Les mathématiques de l’hérédité. Paris, Masson, 63 p.
- MARTIN, Y. 1957. « L’Isle-aux-Coudres : population et économie », Cahiers de géographie, 2 : 167-195.
- MAYER, F. M. 1977. Évolution de la structure génétique d’une population ouverte : La Serpentine. Université de Montréal, thèse de doctorat en anthropologie, 684 p. + annexe.
- MAYER, F. M., et M. BOISVERT. 2005. « Étude de l’organisation sociale à l’Isle-aux-Coudres au 19e siècle à l’aide du recensement de 1851 et du Registre de la population: les non membres de la famille », dans Société de généalogie et d’archives de Rimouski (SGAR). Les Actes du colloque (3e congrès de la Fédération québécoise des sociétés de généalogie, Rimouski, 21-23 mai 2004).
- MAYER, F. M., C. BONAÏTI et J. BENOIST. 1985. « Genealogical approach to the genetic study of hypoacusia in a Caribbean isolate », dans R. CHAKRABORTY et E. J. R. SZATHMARY, éd. Diseases of Complex Etiology in Small Populations, Ethnic Differences and Research Approaches. Progress in Clinical and Biological Research, 194 : 227-244. New York, Alan R. Liss, Inc.
- PHILIPPE, P. 1969. Structure de la consanguinité à l’Isle-aux-Coudres. Université de Montréal, Faculté des sciences sociales, économiques et politiques, Département d’anthropologie, mémoire de maîtrise, 111 p.
- PHILIPPE, P. 1971. « Population logistique à l’Isle-aux-Coudres ? » Biométrie humaine, 6 : 102-111.
- PHILIPPE, P., et J. GOMILA. 1971. « Structure de population et mariages consanguins à l’Isle-aux-Coudres (Québec) », Population, 26, 4 : 707-716.
- PHILIPPE, P., et J. GOMILA. 1972. « Inbreeding effects in a French Canadian isolate. 1. Evolution of inbreeding », Zeitschrift für Morphologie und Anthropologie, 64, 1 : 54-59.
- RIDLEY, M. 1997. Évolution biologique. Paris et Bruxelles, De Boeck Université. Traduction de la 2e édition anglaise (1996) par Raymond Rasmont.
- TREMBLAY, M., É. HEYER et M. ST-HILAIRE. 2000. « Comparaisons intergénérationnelles de l’endogamie à partir des lieux de mariage et de résidence. L’exemple de la population du Saguenay », Cahiers québécois de démographie, 29, 1 : 119-146.
- TREMBLAY, M., M. JOMPHE et H. VÉZINA. 2001. « Comparaison de structures patronymiques et génétiques dans la population québécoise », dans G. BRUNET, P. DARLU et G. ZEI, éd. Le patronyme : histoire, anthropologie et société. Paris, CNRS-Éditions : 367-389.
- VÉZINA, H., M. TREMBLAY et L. HOULE. 2004. « Mesure de l’apparentement biologique au Saguenay-Lac-St-Jean (Québec, Canada) à partir de reconstitutions généalogiques », Annales de démographie historique, 2 : 67-84.
- WRIGHT, S. 1922. « Coefficients of inbreeding and relationship », American Naturalist, 56 : 330-338.
 10.7202/006290ar
10.7202/006290arListe des figures
Figure 1
Couples fin d'information généalogique (FIG), couples immigrants (I)
Figure 2
Distribution, par période de 20 ans, des couples fin d'information ou issus de fin d'information selon divers critères de repérage dans le Registre de population (conjoints nés ou non dans la population)
* Immigrant fin d'information généalogique : fondateur.
Liste des tableaux
Tableau 1
Évolution de la population de l’Isle-aux-Coudres (1721-1967)
Tableau 2
Distribution, par période de 20 ans, des couples fin d’information (A) ou issus de fin d’information (B), selon divers critères de repérage dans le Registre de population, et selon que les conjoints sont nés ou non dans la population a
FIG : fin d’information généalogique.
IFIG : immigrant fin d’information généalogique = fondateur.
Tableau 3
Répartition des unions célébrées à l’Isle-aux-Coudres selon que les conjoints sont apparentés a ou non et selon que les conjoints sont nés ou non dans la communauté
Apparentement calculé ou déclaré (dispenses religieuses).
Tableau 4
Distribution, par période de 20 ans, des mariages célébrés à l’Isle-aux-Coudres (1720-1967), selon que les conjoints sont nés ou non dans la communauté
Tableau 5
Distribution, par période de 20 ans, des unions fécondes célébrées à l’Isle-aux-Coudres (1720-1967), selon que les conjoints sont nés ou non dans la communauté
Tableau 6
Distribution, par période de 20 ans, des unions sans date de mariage connues par naissance d’enfant, selon que les conjoints sont nés ou non dans la communauté
Tableau 7
Distribution, par période de 20 ans, des mariages entre apparentés célébrés à l’Isle-aux-Coudres (1720-1967), selon que les conjoints sont nés ou non dans la communauté
Tableau 8
Distribution, par période de 20 ans, des unions fécondes entre apparentés célébrées à l’Isle-aux-Coudres (1720-1967), selon que les conjoints sont nés ou non dans la communauté
Tableau 9
Pourcentage d’unions fécondes chez les couples mariés (A) et chez les couples apparentés (B) mariés à l’Isle-aux-Coudres (1720-1967), selon que les conjoints sont nés ou non dans la communauté
Tableau 10
Variables dans la valeur de la contribution génétique de deux fondateurs chez les 720 individus en âge de se reproduire en 1967