

Résumés
Résumé
L’épigénétique est un champ d’études qui s’intéresse aux modifications biochimiques et aux changements dans la structure tridimensionnelle (3D) de l’ADN ayant pour effet de contraindre ou de faciliter la lecture et l’expression des gènes. Au cours des dix dernières années, l’épigénétique a attiré l’attention d’un nombre croissant de chercheurs en sciences sociales, puisqu’elle semble venir confirmer, cette fois sur le plan moléculaire, le rôle déterminant de l’environnement développemental (physico-chimique et psychosocial) des personnes dans la configuration de leur individualité biologique et dans la programmation de leur santé future. Cet article se penche sur les implications épistémologiques potentielles de l’épigénétique. Nous distinguons et décrivons trois perspectives socio-anthropologiques complémentaires, adoptées par différents auteurs, sur le rapprochement des concepts de « nature » et de « culture » par l’épigénétique : la socialisation du biologique, la biologisation du social et la superposition nature-culture.
Abstract
Epigenetics is a field of study that focuses on biochemical modifications and changes in the tridimensional (3D) structure of DNA that have the effect of constraining or facilitating the reading and expression of genes. Over the last ten years, epigenetics has captured the attention of several social scientists since it appears to confirm, this time at the molecular level, the determining role of the developing environment of people in configurating their biological individuality and programming their future health. This article discusses the potential epistemological implications of epigenetics. It distinguishes and describes three complementary socio-anthropological perspectives, taken by different authors, on the rapprochement of the concepts “nature” and “nurture” by epigenetics: the socialization of the biological, the biologization of the social, and the superimposition nature-nurture.
Corps de l’article
INTRODUCTION
L’épigénétique est « l’étude des changements dans l’activité des gènes, n’impliquant pas de modification de la séquence d’ADN et pouvant être transmis lors des divisions cellulaires » (INSERM, 2015). Elle s’intéresse au rôle de variations fines de la structure tridimensionnelle de l’ADN (ou chromatine), à des régions très précises et ayant pour effet de contraindre ou de faciliter la lecture et l’expression de gènes spécifiques. Des modifications épigénétiques telles que la méthylation de l’ADN permettent de réguler la production des protéines nécessaires aux différents types cellulaires qui composent un organisme, ce qui confère du même coup aux tissus et aux organes leur activité fonctionnelle particulière.
Au cours des quinze dernières années, la recherche en épigénétique a connu un essor fulgurant. Plusieurs maladies comme les cancers, les maladies cardiovasculaires, les désordres hormonaux, métaboliques ou inflammatoires, et certaines affections neuropsychologiques, ont été associées à une perturbation des mécanismes épigénétiques. L’intérêt des chercheurs en sciences sociales et des médias pour ce domaine fut propulsé, entre autres, par la recherche en épigénétique environnementale, une spécialité s’intéressant plus particulièrement aux causes externes[1] de l’altération à long terme des mécanismes épigénétiques. L’épigénétique environnementale suscite une vive attention parce qu’elle met en lumière, cette fois sur le plan moléculaire, le rôle déterminant de l’environnement physico-chimique (par exemple les polluants) et psychosocial (par exemple le stress) des personnes dans la configuration de leur spécificité phénotypique (Müller et al., 2017) – un effet qui pourrait même, dans certains cas, s’étendre au-delà des générations (Heard et Martienssen, 2014). Ce nouveau domaine de recherche apporte donc un éclairage supplémentaire sur la relation existant entre phénomènes sociaux et traits biologiques (Dupras et al., 2014; Meloni, 2014b).
Dans cet article, nous présentons les observations, les interprétations et les spéculations les plus importantes, formulées au cours des dix dernières années par des chercheurs provenant de diverses disciplines comme l’anthropologie, la philosophie, les sciences politiques et la sociologie, au sujet des implications épistémologiques potentielles du développement des connaissances en épigénétique. L’épigénétique soulève des questions hautement sensibles, qui mériteront une attention accrue et soutenue dans le futur, telles que l’accès aux données épigénétiques et la protection de la vie privée des participants à la recherche (Diemer et Woghiren, 2015; Dyke et al., 2015), ou encore le risque de pratiques discriminatoires basées sur l’information épigénétique des personnes (Rothstein et al., 2009; Rothstein, 2013; Saulnier et Dupras, 2017). L’analyse des implications éthiques, légales et sociales de l’épigénétique n’est toutefois pas l’objectif premier de cet article. Nous avons plutôt choisi de donner préséance aux écrits de type socio-anthropographique, et ce, afin de proposer une cartographie détaillée des perturbations épistémologiques sous-jacentes aux implications normatives potentielles de ce jeune champ de recherches. Ce niveau d’analyse nous permettra de mieux comprendre les assises conceptuelles des analyses prescriptives émergentes, de distinguer des perspectives en tension les unes avec les autres et de reconnaître l’étude des implications épistémologiques de l’épigénétique comme un champ de contestation.
La thèse centrale qui est explorée dans cet article est l’idée selon laquelle l’épigénétique serait sur le point d’amorcer une réconciliation des concepts depuis trop longtemps érigés en dichotomie de « nature » et de « culture ». En faisant la lumière sur les mécanismes biologiques par lesquels l’environnement des personnes (culture) entre en interaction avec leurs gènes (nature), la recherche en épigénétique ébranlerait les fondements théoriques mêmes de ce « schisme » conceptuel[2]. Selon Guthman et Mansfield, « cet échange de molécules environnementales et corporelles suggère une transformation de ce que nous entendons par “nature” et “culture”, d’une telle manière que la démarcation entre elles semble s’effacer, même s’anéantir » (2012, p. 12). La nouvelle « perméabilité » entre les pôles nature et culture proposerait, du même coup, une relation renouvelée, plus dynamique, entre les sciences naturelles et les sciences sociales : « La sociologie s’ouvre aux suggestions de la biologie, au moment même où la biologie devient sociale » (Meloni, 2014b, p. 594). À l’aube d’une nouvelle ère, qualifiée par Griffiths et Stotz (2013) de « post-génomique », les représentations théoriques émergentes de « gène socialisé », de « cerveau social » ou de « vision pro-sociale de l’évolution » ne sont que quelques illustrations du rapprochement conceptuel et disciplinaire vraisemblablement en cours.
Notre article est divisé en trois sections, correspondant à trois perspectives (ou approches) distinctes, adoptées par différents auteurs, dans leur étude des implications épistémologiques de l’épigénétique environnementale : la socialisation du biologique, la biologisation du social et la superposition nature-culture (figure 1). Nous nous intéresserons d’abord à l’impact de l’épigénétique sur les théories de l’évolution et sur le déterminisme biologique. Comme nous le verrons, l’épigénétique s’illustre pour plusieurs auteurs par sa capacité probable à perturber les théories néodarwiniennes de l’évolution et les vues gène-centristes de l’individualité humaine. Nous nous pencherons ensuite sur le déplacement conceptuel opposé du concept de culture vers celui de nature, créé notamment par l’internalisation et la numérisation de l’espace et du temps social par l’épigénétique. Nous discuterons entre autres de la menace réductionniste que ce nouveau champ d’études semble représenter pour certains auteurs. Dans la dernière section de l’article, nous verrons comment l’entrelacement conceptuel nature-culture pourrait affecter certains processus de normalisation et de stratification des populations humaines (par exemple les distinctions de genre ou la racialisation). Nous explorerons aussi de quelles manières les analyses de l’épigénétique semblent se constituer progressivement, entre espoir et méfiance, au gré des métaphores mobilisées, en un « discours épigénétique » parfois intéressé et, ainsi, en proie à la distorsion et à l’inflation. Enfin, nous étudierons la façon dont l’épigénétique pourrait bouleverser la conduite de la recherche en sciences sociales et en sciences naturelles au cours des années à venir.
Figure 1
Dissection du rapprochement nature-culture
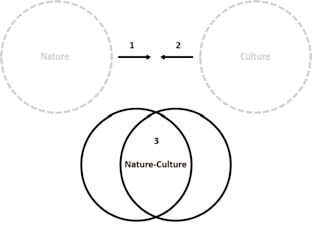
Perspectives distinctes et complémentaires sur les implications épistémologiques potentielles de l’épigénétique : 1) socialisation du biologique, 2) biologisation du social et 3) superposition nature-culture.
SOCIALISATION DU BIOLOGIQUE
Impact sur les théories de l’évolution
Depuis la complétion du Projet Génome Humain (2003), une période de réévaluation critique des théories gènes-centristes de l’évolution et de l’identité humaine s’est installée. En cette nouvelle ère « post-génomique », la nature humaine n’apparaît plus simplement biologique et immuable, mais également façonnée et modulée par son environnement (Meloni, 2015a). L’ère post-génomique marque la fin de l’hypothèse – que l’on reconnaît aujourd’hui simpliste, mais qui fut dominante durant la seconde moitié du xxe siècle – selon laquelle à un gène correspondrait un phénotype, et inversement, à un phénotype correspondrait un gène. Elle se traduit par l’importance de phénotypes multigéniques, de la régulation de l’expression des gènes, des interactions moléculaires, de la complexité étiopathogénique et des approches biopsychosociales. Dans ce contexte, l’épigénétique ne représenterait pas seulement un tournant chronologique, une nouvelle page d’histoire scientifique, mais également un tournant épistémologique menant à une transformation significative de la valeur théorique et pratique du gène, entre autres, en tant qu’unité fondamentale au centre des théories néo-darwinistes de l’évolution (Meloni et Testa, 2014; Lappé et Landecker, 2015).
Les mécanismes de programmation épigénétique suggèrent que la nature humaine est, dans une certaine mesure, adaptative (Godfrey et al., 2007). En fonction de la disponibilité des nutriments durant le développement de l’enfant, par exemple, des gènes associés au métabolisme des glucides seraient activement configurés de façon à optimiser l’utilisation et le stockage des calories consommées au cours de la vie. Ce processus d’optimisation métabolique favoriserait la survie de l’espèce en situation d’abondance ou de pénurie alimentaire (Drake et Liu, 2010). Une programmation épigénétique similaire permettrait à l’organisme de prévoir une réponse adéquate aux situations anxiogènes en fonction de la dangerosité de l’environnement développemental (Gershon et High, 2015). Entre l’adaptation biologique des individus à très court terme, grâce aux processus homéostatiques, et l’évolution des espèces à très long terme, grâce aux mutations génétiques aléatoires et à la sélection naturelle, les mécanismes épigénétiques permettraient une adaptation sur une échelle de temps « intermédiaire », s’étendant possiblement, dans certains cas, à quelques générations (Kuzawa et Thayer, 2011).
À ce jour, la plupart des découvertes publiées au sujet de la transhéritabilité des variants épigénétiques sont issues d’études menées chez des animaux non humains. Les conclusions sur la transmission de variants épigénétiques acquis, d’une génération aux suivantes, sont encore incertaines et font l’objet de controverses (Juengst et al., 2014; Rothstein et al., 2017). Pour certains auteurs, l’épigénétique environnementale devrait tout de même pousser les chercheurs à élargir la théorie synthétique de l’évolution des espèces (néo-darwinisme)[3] de façon à tenir compte des systèmes « écologiques » (ecologically-relevant) et interactionnistes d’hérédité (Bossdorf et al., 2008; Lerner et Overton, 2014). Pour eux, une théorie de l’évolution appropriée doit considérer l’interrelation entre processus adaptatifs et sélectifs, à la façon du modèle de « causalité récursive » de Rupert Riedl. Également surnommé « théorie de la complexité en évolution » (theory of evolving complexity), ce modèle est fondé sur l’analyse des mécanismes de rétroaction. Il aurait ainsi pour avantage d’accorder une plus grande attention à l’interaction complexe qui semble exister entre génétique et épigénétique (Sagl et al., 2007).
En atténuant l’attention portée auparavant de façon disproportionnée sur le réplicateur (le « gène égoïste » de Dawkins, 1976) à la faveur du véhicule (l’organisme ou l’individu pris dans toute sa complexité et ses interactions avec son environnement), l’épigénétique soulignerait le rôle crucial de la collaboration entre les membres d’une communauté sur les processus évolutifs. Ainsi, elle permettrait de dépasser la seule analyse des conséquences biologiques de l’évolution, tâche que s’était réservée la génétique moléculaire (Bateson, 2014). L’épigénétique offrirait même possiblement une solution moléculaire à la « difficulté spéciale » de Darwin : c’est-à-dire au mystère de la conservation des castes d’abeilles non fertiles – mais visiblement nécessaires à la survie de l’essaim – par le seul processus de sélection de gènes favorable à la survie et à la reproduction des individus. Les mécanismes épigénétiques, en permettant de dériver plusieurs phénotypes distincts à partir d’un seul génotype, seraient responsables de la division du travail chez les insectes sociaux et, ainsi, de la préservation des castes vulnérables[4]. Ces mécanismes biologiques auraient été conservés au fil de l’évolution parce qu’ils confèrent au collectif un avantage important face à la pression sélective de l’évolution darwinienne, en favorisant la survie et la reproduction du groupe et non seulement des individus (Herb, 2014).
La vision pro-sociale de l’évolution s’intéresse au rôle de l’altruisme et de la collaboration pour la survie des espèces (Meloni, 2014b). À cet égard, l’épigénétique apparaît confirmer, pour certains auteurs, une relation intime entre changements socioculturels et modifications biologiques, lorsqu’elle découvre les mécanismes moléculaires par lesquels une même espèce peut évoluer à travers les âges pour s’adapter aux contraintes imposées par son environnement. De concert avec le phénomène de construction de niches favorables[5], une adaptation biologique rapide aux aléas de l’environnement serait rendue possible grâce au caractère « fluide » des mécanismes épigénétiques. Pour plusieurs observateurs tels que Brooke et Larsen (2014), de telles analyses semblent conférer une aux découvertes en épigénétique une saveur « quasi lamarckienne » (ou néo-lamarckienne)[6].
Malgré les incertitudes scientifiques patentes et la résistance à l’égard d’une réhabilitation des thèses lamarckiennes (Haig, 2007; Juengst et al., 2014; Whitelaw, 2015), l’hypothèse d’une transmission transgénérationnelle de certains variants épigénétiques est encore à ce jour admissible – et toujours à l’étude (Heard et Martienssen, 2014). Deux observations importantes réduisent toutefois la probabilité d’une telle transmission : la séparation weismannienne[7] entre les cellules somatiques et germinales, et l’existence de deux phases de reprogrammation des cellules souches embryonnaires durant le développement foetal (Richards, 2006). En d’autres termes, l’absence d’un mécanisme connu de communication des modifications épigénétiques acquises par les différentes cellules du corps aux cellules reproductrices, et la (re)méthylation quasi totale de l’ADN durant l’embryogenèse, sont deux conditions extrêmement restrictives qui limitent la possibilité d’une transmission directe de telles modifications aux générations suivantes[8].
Pour certains auteurs, cependant, les critiques alarmistes qui condamnent d’emblée toutes les thèses à saveur lamarckienne – alimentées par exemple par la crainte d’un regain du lysenkoïsme[9] – contribuent à stigmatiser de façon exagérée les théories de l’hérédité douce (soft heredity) vraisemblablement appuyées par les progrès en épigénétique (Richards, 2006). En fait, l’idée selon laquelle l’hérédité épigénétique risque de réveiller « les fantômes » des théories de l’évolution pécherait elle-même d’un extrémisme inadmissible. C’est d’ailleurs cette culture dualiste, entre deux écoles froidement opposées (lamarckisme et darwinisme), qui serait l’obstacle principal à une meilleure compréhension de l’hérédité (Meloni, 2015b). Pour parvenir à un modèle adéquat, une collaboration renouvelée entre historiens, biologistes de l’évolution et anthropologues de la physiologie apparaît nécessaire (Brooke et Larsen, 2014). Les modèles simplistes de la biologie évolutive sont encore aujourd’hui persistants. Ils alimentent le scepticisme sur la pertinence d’inclure les mécanismes épigénétiques dans les théories de l’évolution (Gadjev, 2017).
Impact sur le déterminisme biologique
L’épigénétique représente pour plusieurs observateurs un élément de preuve tangible des limites problématiques des thèses déterministes prétendant expliquer par les gènes la majorité des variations biologiques, comportementales et identitaires entre les personnes et les populations. Une expression vivement contestée du déterminisme génétique, par exemple, est la réduction des comportements sociaux des individus à de simples composantes génétiques fixées dès la naissance – interprétation souvent alimentée par la recherche en sociobiologie. Désormais, la recherche en épigénétique nous offrirait l’occasion de repenser les formulations biologiques de l’individualité et de la psychologie humaine, sans toutefois tomber dans le piège de l’essentialisme génétique (Vijver et al., 2002; Canning, 2008; Kuzawa et Sweet, 2009). Celles-ci ne seraient plus simplement fixées à la conception et dès la naissance, mais seraient aussi progressivement construites suivant l’« histoire épigénétique » des personnes (Boniolo et Testa, 2012).
Comme l’observe Tolwinski (2013), plusieurs chercheurs affichent ouvertement un vif enthousiasme à l’égard de l’épigénétique. Certains croient d’ailleurs qu’elle aurait le pouvoir d’insuffler une « épistémologie anti-déterministe » en biologie. Gonon et Moisan, par exemple, soutiennent qu’« à l’inverse d’une présentation déterministe de la génétique, l’épigénétique nous propose une conception beaucoup plus complexe, souple et imprévisible de l’histoire individuelle » (2013, p. 29). La grande promesse de l’épigénétique, en tant que « science de la plasticité », résiderait dans l’idée même qu’il s’agit d’une science « non déterministe » (Mansfield et Guthman, 2015). Ainsi perçue, l’épigénétique ferait écho au stochastisme, théorie selon laquelle bon nombre de processus biochimiques surviennent de façon aléatoire et non prédictible (Landecker et Panofsky, 2013).
Selon d’autres auteurs, l’épigénétique risque au contraire d’introduire de nouvelles formes de déterminisme (Richardson, 2015). La « formalisation de la nourriture » en tant que facteur de risque pour la santé épigénétique, par exemple, pourrait renforcer le « déterminisme environnemental » (Landecker, 2010). De façon similaire, les travaux sur l’impact de la classe sociale ou du statut socio-économique sur la santé épigénétique pourraient générer des interprétations déterministes qui imputeraient une relation de causalité stricte de la culture vers la nature (Pickersgill et al., 2013) :
[…] in contrast to the received view of epigenetics as anti-deterministic or anti-essentialist, epigenetics research is often couched in language as deterministic as genetics research […] contemporary epigenetics discourse may be understood under the framework of epigenetic determinism, which we conceive as the belief that epigenetic mechanisms determine the expression of human traits and behaviors
Waggoner et Uller, 2015, p. 178
Le déterminisme épigénétique pourrait émerger d’au moins trois considérations : de l’influence de certains gènes sur la variabilité épigénétique, de l’influence de l’environnement développemental sur la programmation épigénétique, et de la possible transmission familiale de certaines modifications épigénétiques acquises (Waggoner et Uller, 2015). Pour certains auteurs, la prudence est de mise vu la recrudescence récente d’une « variété de nouveaux matérialismes » (Niewöhner, 2015) pouvant générer une nouvelle vague de « déterminisme somatique » (Lock, 2013) et un retour à un « cadre analytique déterministe dépassé » (Stelmach et Nerlich, 2015).
Si l’épigénétique nous permet de remettre en question les thèses gènes-centristes de l’évolution et de l’individualité humaine, elle n’implique pas sans condition l’abandon de celles-ci. Pour Waggoner et Uller, « plusieurs de ceux qui saluent l’épigénétique comme une solution au déterminisme biologique remplacent simplement un code déterministe par un autre » (2015, p. 191). Selon Meloni et Testa, « l’ouverture du génome au social […] est toujours sur le bord de s’effondrer et d’être réduite à une simple source de variabilité de l’expression des gènes » (2014, p. 449). Ces auteurs observent toutefois que les fondements épistémiques de ce nouveau risque de « réductionnisme épigénétique » sont particuliers :
Whereas in late twentieth century gene-centrism, from sociobiology onwards, we found an increasing attempt to expand the reach of nature into the field of nurture, here the somatic reductionism of epigenetics is the effect of an opposite epistemic claim: that neither nature nor nurture makes sense anymore, and everything is part of an integrated and blurred nature–nurture ontogenetic system
Meloni et Testa, 2014, p. 449
De façon similaire, c’est précisément parce qu’elle s’intéresse aux systèmes biologiques complexes, interactifs et influencés par des facteurs environnementaux et sociaux externes au corps que la recherche sur les origines développementales de la santé et de la maladie (DOHaD) est, à l’heure actuelle, en pleine effervescence (Wallack et Thornburg, 2016; Rial-Sebbag et al., 2016). L’épigénétique, une composante importante du mouvement DOHaD, paverait donc la voie pour une théorie et une pratique du juste milieu entre « nature » et « culture », une nouvelle approche que nous pourrions qualifier, avec Meloni (2015a), de « constructivisme corporel » (embodied constructivism).
BIOLOGISATION DU SOCIAL
Reconfiguration de l’espace social
En plus de prêter attention à la socialisation du biologique, quelques chercheurs en sciences sociales se sont intéressés, à l’inverse et de façon complémentaire, à la biologisation du social par la recherche en épigénétique. Ce déplacement conceptuel s’observerait, entre autres, par l’internalisation et la matérialisation de l’espace social. Nous assisterions ainsi, notamment, à la molécularisation de l’environnement :
Food’s biological activity and its connection to human health, its pathogenicity or therapeutic power, becomes understood in terms of how outer molecules articulate with inner molecules in a life of eating. […] The rise of “functional foods” places the emphasis on particular biologically active substances rather than on genes (or lack thereof), but the molecular optic here is also intensely cultivated
Landecker, 2010, p. 185-186
La recherche en épigénétique expose les mécanismes biochimiques par lesquels l’environnement développemental « s’intègre sous la peau » des personnes (Meloni, 2014a; Osborne, 2015). Elle nous conduirait ainsi vers une sorte d’internalisation des conditions de vie (Niewöhner, 2011; Dupras et Ravitsky, 2016b). L’adversité sociale, par exemple, serait soumise à une « intégration corporelle » (embodiment) (Thayer et Non, 2015). Cette transition, qui se produirait non seulement sur le plan moléculaire mais également sur les plans conceptuel et sémantique, serait sur le point d’offrir à la description des phénomènes sociaux et au concept de culture un langage matérialiste normalement réservé à la biologie (Davis, 2014).
L’épigénétique environnementale contribuerait par exemple à la « numérisation de l’environnement », une reconfiguration de l’environnement en des termes permettant désormais de le mesurer, de l’archiver et de le comparer plus facilement[10]. Dès lors, les signaux issus de l’environnement se retrouveraient encodés dans le vivant. Déchiffrer le mystère de leur origine complexe, multiple et diffuse reviendrait donc à examiner des codes moléculaires, « à la différence [toutefois] que ces codes prennent désormais la forme d’interrupteurs flexibles et réversibles plutôt que de circuits fixés » (Meloni et Testa, 2014, p. 441). L’épigénétique nous conduirait ainsi vers une nouvelle forme de socialité imprégnée dans les corps et la biologie humaine, une « socialité somatique » :
People do not need to translate their somatic self-understanding into a way of life any longer. Ways of life themselves are already standardised and correlated directly with their epigenetic effects on the human body […] Somatic sociality means sociality understanding and reproducing itself on the basis of biological knowledge about its epigenetic effects on the individual body. [It] is a form of sociality where the collectivising momentum is provided by molecular understandings of social life itself
Niewöhner, 2011, p. 292
Le type de molécularisation de la vie sociale qui semble se produire avec l’épigénétique apparaît cependant quelque peu différent de la molécularisation observée par le passé dans le contexte de la génétique. Il ne s’agirait plus désormais de réduire des comportements humains ou des phénomènes sociaux à la présence ou à l’absence d’un gène spécifique (par exemple le « gène guerrier » des peuples maoris), mais plutôt de reconnaître les interactions complexes et dynamiques entre l’espace extérieur et le corps compris comme « bioactif » : « Il ne s’agit pas d’une fusion de l’interne et de l’externe sous prétexte que tout est moléculaire, mais d’un réarrangement de l’interrelation » (Landecker, 2010, p. 180).
Reconfiguration du temps social
Le déplacement du concept de culture vers le concept de nature suggéré par la recherche en épigénétique se caractériserait aussi par une intégration corporelle du caractère temporel de l’expérience sociale: une sorte de « “perpétuation” des structures à travers le temps » par la biologie humaine (Landecker et Panofsky, 2013, p. 3). L’alimentation, par exemple, ne serait plus simplement perçue comme une réponse temporaire à des besoins nutritionnels ponctuels, mais aussi comme une source de transformation permanente de l’organisme sur le plan moléculaire :
This is a model in which food enters the body and in a sense never leaves it, because food transforms the organism’s being as much as the organism transforms it. It is a model for how social things (food, in particular) enter the body, are digested, and in shaping metabolism, become part of the body-in-time, not by building bones and tissues, but by leaving an imprint on a dynamic bodily process
Landecker, 2010, p. 177
C’est entre la plasticité et la stabilité des variants épigénétiques que semble s’opérer une sorte de dialectique de l’empreinte fondée sur des « états moléculaires semi-stables » (Loi et al., 2013). Avec l’épigénétique, l’attention de la communauté scientifique est ainsi réorientée, selon Landecker et Panofsky (2013), d’une conception obsolète du génome comme entité statique indépendante du temps vers la régulation des gènes en fonction des périodes de sensibilité des épigénomes au cours de la vie, de la fréquence et de la durée des expositions, puis de la persistance temporelle relative des différents variants épigénétiques acquis.
Si l’épigénétique introduit le concept de « génome réactif » soumis aux aléas du temps qui passe (Meloni, 2014c), elle souligne également l’importance de considérer la durée des effets d’une exposition environnementale ou d’une expérience sociale; ces effets étant eux-mêmes des objets physiques ayant leur propre durée de vie. Par exemple, le danger des polluants xénobiotiques résiderait non seulement dans une accumulation chronologique d’expositions à ces produits chimiques situés à l’extérieur du corps, mais surtout dans la transformation persistante de ses parties (Kuzawa et Sweet, 2009). Pour Lock, lorsque des périodes comme les générations, les trimestres de la gestation, l’adolescence ou la ménopause sont redéfinies par des taux de méthylation de l’ADN, « les effets de ces passages du temps deviennent miniaturisés dans les corps » (2015, p. 137). Ainsi voit-on émerger l’idée d’une « biohistoire » (Brooke et Larsen, 2014), d’un passé qui serait imprégné dans la chair de chacun. Autrement dit : « [t]he past bodily lives on – is re-membered – in our epigenetic markers » (Sullivan, 2013, p. 21).
Selon Warin et al. (2015), l’épigénétique serait compatible avec le caractère flexible et la capacité reproductrice des notions d’« habitus » et de « capital corporel » de Pierre Bourdieu.
In reading Bourdieu’s theory of practice in light of epigenetics, it is clear that bodies are embedded in temporo-spatial landscapes of materiality and history. The habitus creates a system of dispositions – “a past which survives the present and tends to perpetuate itself into the future by making itself present in practices” (Bourdieu 1977, p. 82). Extending habitus into biohabitus allows us to understand the situated and contingent nature of bodies across time
Warin et al., 2015, p. 68-69
Le concept de « biohabitus » proposé par ces auteurs est à la fois structuré et structurant : il s’agit d’« un produit et un producteur de mondes sociaux ». Son caractère cyclique rappellerait d’ailleurs la difficulté d’établir une démarcation nette entre extériorité et intériorité, puis d’intervenir sur des processus et des pratiques qui se renforcent par eux-mêmes et s’autoperpétuent à travers le temps (Warin et al., 2015).
Il est cependant possible, selon Lappé et Landecker (2015), que cette nouvelle temporalité qu’acquiert la biologie au moyen des avancées récentes en épigénétique ne puisse être réservée à l’épigénome seul. Si la méthylation de l’ADN est la plupart du temps considérée comme une modification biochimique n’affectant pas la séquence nucléotidique, la nature et les propriétés chimiques d’une cytosine méthylée sont, en réalité, altérées de façon significative. Pour cette raison, il semble que l’intégrité de la séquence d’ADN, après une telle modification chimique, doive être remise en question. Un paradoxe apparaît aussitôt, puisqu’en même temps que le discours dominant configure l’épigénétique comme champ d’études distinct de la génétique, les résultats générés par la recherche proposent un génome dépendant, lui aussi, du temps et des périodes de la vie. Pour Lappé et Landecker, c’est toute la codification biologique qui, à la suite de l’épigénétique, serait « ordonnée de façon narrative en fonction de la durée de la vie humaine » (2015, p. 153). Le déplacement conceptuel devrait donc s’appliquer tant au génome qu’à l’épigénome, en tant que transition « de la vie comme information au temps de la vie comme conformation » (« from life as information to lifetime as conformation ») (Lappé et Landecker, 2015, p. 171), c’est-à-dire de la vie comme produit d’une information génétique codifiée et numérique, au cours de la vie en tant que variation de la structure tridimensionnelle de la chromatine.
SUPERPOSITION NATURE-CULTURE
Selon Meloni et Testa (2014), la superposition des concepts de nature et de culture pourrait influencer l’étude des politiques publiques d’au moins trois façons : par l’émergence d’un « discours » par lequel de nouvelles normes et catégories populationnelles seront définies et caractérisées, les anciennes déconstruites ou renforcées; par l’éloge de la recherche en épigénétique comme « bastion ultime de preuves biologiques » sur la base desquelles il est possible de promouvoir des idéologies politiques, des dispositifs biopolitiques et des pratiques socioculturelles spécifiques; et par la reformulation de théories sociales et politiques[11] reposant sur la distinction entre nature et culture.
Normes, catégories et biopolitiques
Au cours de l’histoire, des groupes d’individus partageant des caractéristiques communes ont parfois été définis, soit en fonction de traits biologiques communs (semblables par nature), soit en fonction de comportements, de coutumes ou d’institutions sociales (semblables par culture) (Paul, 1998; Singh, 2012). Avec l’épigénétique, s’il n’est plus question d’opposer nature et culture, un regard nouveau sur les similitudes et les particularités existant au sein de l’espèce humaine apparaît nécessaire (Hendrickx et Van Hoyweghen, 2018). La question de l’attribution d’une valeur ou d’une fonction sociale à certains groupes d’individus – d’un rôle parental à un genre particulier, par exemple – se retrouverait plus que jamais au coeur de discussions particulièrement sensibles.
Distinctions de genre
Des études conduites en laboratoire chez les rats ont suggéré qu’un déficit en affectivité maternelle, tôt durant la vie, pouvait contribuer à une programmation épigénétique potentiellement pathogénique chez la progéniture. Prises de façon isolée et transposées hâtivement à l’espèce humaine, par les médias par exemple, de telles études pourraient renforcer injustement l’idée d’une spécialisation des genres, en suggérant que c’est avant tout de l’affectivité maternelle (et non paternelle) dont les petits ont besoin durant la tendre enfance (Lappé, 2016a). Quelques critiques féministes se sont d’ailleurs récemment intéressé·e·s aux divers processus par lesquels « la recherche en épigénétique configure le corps maternel en tant que site principal de programmation et de transmission épigénétique, puis en tant que cible principale d’interventions médicales et de santé publique » (Richardson, 2015, p. 210). La conceptualisation du corps maternel en tant qu’environnement dans la recherche sur l’autisme (Lappé, 2016b), par exemple, ou de façon plus générale « comme vecteur épigénétique » (Richardson, 2015) principal, à partir des études disponibles à ce jour, apparaît pour plusieurs extrêmement réductrice et non fondée (Sullivan, 2013; Valdez, 2018).
Selon Osborne (2015), la théorie de la construction sociale des genres de la féministe Judith Butler[12] pourrait être affectée par l’épigénétique. Pour Butler, la distinction des genres n’est pas un phénomène « naturel » et ne devrait pas s’appuyer strictement sur la base du sexe physique. Au contraire, les distinctions de genre s’apprennent, se pratiquent en se répétant, puis se « performent » – à la manière d’un rôle au théâtre. Les distinctions de genre seraient imposées par la tradition et les structures sociales, puis renouvelées par le produit de cette performance. Osborne (2015) a récemment proposé une réinterprétation de cette théorie de la performativité du genre. Pour l’auteur, l’analyse originale de Butler est quelque peu problématique, parce qu’elle se confine elle-même au « camp de la “culture” » et s’inscrit par défaut en rivale (plutôt qu’en complément) à toute corrélation biologique de la notion de genre. En réponse à cette faiblesse, l’épigénétique offrirait une piste de solution :
epigenetics offers a conceptual bridge that allows Butler’s theory of gender performativity to be used productively in conjunction with biological explanations of gender difference. […] epigenetics moves us away from the notion of gender identity as a fixed entity, and towards Butler’s argument that gender is instead an unstable constellation of behaviours
Osborne, 2015, p. 508
L’épigénétique permettrait ainsi de réconcilier les concepts de « nature » et de « culture » érigés en dichotomie par Butler tout en appuyant son idée importante d’une itération continue de la construction du genre, qu’elle comprend comme une « stylisation répétée du corps ».
Racialisation
La susceptibilité accrue des Afro-Américains aux maladies cardiovasculaires et aux naissances prématurées a été associée à des variants épigénétiques transmissibles entre les générations. Selon certains observateurs, les études qui permettent de faire de telles découvertes devraient servir à mieux comprendre et à minimiser les inégalités de santé injustes entre différents groupes (Kuzawa et Sweet, 2009; Dupras et al., 2014). Selon d’autres auteurs, cependant, il est possible que ce type d’étude contribue de façon insidieuse à accroître le phénomène de « racialisation » en renforçant l’idée selon laquelle il existe des distinctions fondamentales entre les membres de différentes races humaines (Meloni, 2017; Saulnier et Dupras, 2017) :
while it might seem that these new epigenetic models of plastic life should eliminate race by eliminating notions of discrete kinds given in nature, it appears that epigenetics offers a new form of racialization based on processes of becoming rather than on pre-given nature
Mansfield et Guthman, 2015, p. 6
Un tel risque de réification biologique des races apparaît particulièrement évident lorsque sont impliqués des traits physiques ou psychologiques dont le caractère pathologique est discutable, comme l’obésité (Guthman, 2012). Selon Mansfield (2012), des politiques de santé ciblant les vulnérabilités épigénétiques de certains groupes pourraient d’ailleurs intensifier le phénomène de racialisation lorsqu’elles prescrivent des changements de comportement préventifs uniquement à ces groupes. Aux États-Unis, ce type de stratégie de santé publique a été critiqué dans le contexte d’une mise en garde gouvernementale portant sur une trop grande consommation de poisson. Si cette mesure visait à informer la population sur les risques d’une contamination au méthylmercure chez les femmes enceintes, une conséquence immédiate fut l’exclusion et la stigmatisation d’habitudes alimentaires propres à des groupes socioculturels minoritaires.
La reconnaissance de vulnérabilités épigénétiques est étroitement liée à la définition d’une « normalité épigénétique » et, subséquemment, de catégories nosologiques. Cette entreprise complexe et sensible mérite une attention particulière si elle se voit offrir les assises d’interprétations prescriptives en santé (Dupras et Ravitsky, 2016a). La notion de « qualité d’un épigénome », par exemple, est déjà le sujet de débats scientifiques et sociaux, avec le déploiement récent d’efforts importants par les consortiums internationaux pour déterminer les « épigénomes références » (Meloni et Testa, 2014). La stratification épigénétique de la population pourrait se traduire en biopolitiques d’au moins deux sortes : l’élaboration de politiques de santé ayant pour but de surveiller, de monitorer et d’intervenir directement auprès des populations vulnérables, et la promotion de saines habitudes de vie que les personnes devraient adopter pour minimiser les risques épigénétiques à leur santé ou à celle d’autrui.
Already public health and the ever-popular “self-management” literature have taken up this new relevance of the organismic environment. Interestingly, environmental epigenetic knowledge is equally readily adopted by those in favor of increasing social welfare spending and public health measures to reduce social inequality as it is by those in favor of increasing individualistic attention to early life development
Niewöhner, 2015, p. 224
Pour certains auteurs, les préoccupations épistémologiques et biopolitiques les plus importantes semblent résider dans « la naturalisation brute et la réification subséquente de configurations matérielles-sémiotiques [surgissant des] lectures déterministes du genre, de la classe sociale et du comportement spécifique de certains groupes » (Niewöhner, 2015, p. 224; Meloni, 2017; Katz, 2013). Pour d’autres, les biopolitiques axées sur l’autonomisation (empowerment) et la responsabilisation des individus apparaissent les plus susceptibles de teinter l’application des connaissances en épigénétique (Landecker, 2010; Chiapperino et Testa, 2016; Dupras et Ravitsky, 2016b).
Métaphores et instrumentalisation d’un discours
Pour plusieurs observateurs, l’explication mécaniste de la régulation des gènes récemment offerte par l’épigénétique représente un « “hard” data » persuasif, une preuve matérielle tangible de l’influence critique de l’environnement sur la biologie, pouvant procurer un élan épistémique suffisant au rejet des théories gène-centristes du xxe siècle (Lock, 2015). Certains chercheurs en sciences sociales ont étudié l’épigénétique comme « discours » : par ses métaphores, son instrumentalisation politique et les différents degrés d’adhésion (et de contestation) qu’il suscite. Pour Mansfield (2012), par exemple, si l’épigénétique active l’imaginaire de la population parce qu’elle semble s’attaquer au fatalisme du déterminisme génétique, il faut comprendre que l’épigénétique n’est pas une vérité en soi et qu’elle possède de multiples visages.
Les métaphores sont en effet fréquentes et variées dans ce nouveau champ d’études. Alors que le génome était comparé à un livre, un code ou un programme, une carte ou un plan, les mécanismes épigénétiques seraient le plus souvent comparés à un interrupteur (switch) ou à un commutateur (switching), et les modifications épigénétiques à des marques (mark), à des étiquettes (tag) ou à des vignettes (label). D’autres images, se rapportant par exemple à la musique – le génome serait la partition et l’épigénétique le chef d’orchestre –, ont aussi été utilisées (Stelmach et Nerlich, 2015). Comme le font remarquer Meloni et Testa (2014), la puissance symbolique et rhétorique de ces métaphores a été exploitée aussi bien à des fins politiques que commerciales. Le plus souvent, elles serviraient à mettre en scène le pouvoir de l’environnement et de l’histoire des personnes sur la biologie.
Whereas in the past the direction of claims making went from the inside out, claims making now goes from the outside in. […] There is also much more of a focus on time and space, in terms of epigenetic effects potentially spanning several generations in time and spanning everything from the inside of the womb to social and cultural environments in space
Stelmach et Nerlich, 2015, p. 213
L’épigénétique serait aussi comprise comme une sorte d’« écriture » sur les gènes. Puis, lorsque persistante, elle se transformerait en une sorte de « mémoire » cellulaire des expositions ou des comportements individuels passés. Quelques analogies ont aussi été élaborées entre les mécanismes épigénétiques et certaines oeuvres littéraires populaires (Middleton, 2015).
L’adhésion au vocabulaire métaphorique émergent et à la promesse épigénétique serait toutefois hétérogène entre les parties prenantes (Pickersgill, 2016). Des entretiens semi-dirigés avec 26 chercheurs en épigénétique ont d’ailleurs permis à Tolwinski (2013) d’observer trois idées importantes sur lesquelles les experts ne s’entendent vraisemblablement pas : « la signification de l’épigénétique, la place accordée à l’incertitude dans les débats […] et les implications plus larges de ce champ » (p. 371). Trois groupes de chercheurs ont été créés sur la base de ces sujets de discorde : les « promoteurs de l’épigénétique » (champions), les « chercheurs modérés » (middle-ground) et les « sceptiques » (Tolwinski, 2013). La signification distincte accordée par différents observateurs à l’épigénétique pourrait être appelée à devenir un objet d’étude en soi.
Epigenetics […] can be seen as a site of (sometimes) competing discourses and expectations regarding what the science is, what it can do, and what its implications are for society […] promissory discourses circulate widely – galvanising research support and civil engagement, but also energising contestation and critique
Pickersgill, 2014, p. 481
Déjà, un nombre croissant de chercheurs en sciences sociales semblent s’intéresser aux raisons, aux postures et aux valeurs sous-jacentes à l’instrumentalisation de l’épigénétique à des fins politiques. Cette approche réflexive soulève plusieurs questions importantes, telles que :
how is epigenetics being imagined and developed; what forms of knowledge, practices and applications are considered salient; and how are these being compared and related to genomics? […] what notions of society and social groups are implicit within research and materialized through it; how will political and clinical gazes be (re)orientated through recourse to epigenetic imaginaries; and what kinds of social and ethical responses will be appropriate in light of any ensuing shifts in policy and practice?
Pickersgill et al., 2013, p. 442
La qualité de la diffusion des résultats de la recherche par les médias devrait aussi susciter l’intérêt des chercheurs, dans un contexte ou « [d]e nombreux documents médiatiques […] sont peu explicites ou même faux concernant la transmission d’une génération à l’autre » (Gonon et Moisan, 2013, p. 8). Pour certains auteurs, c’est grâce à une approche critique du discours épigénétique que « les postulats spéculatifs, les discours gonflés et les promotions médiatiques enthousiastes, en un mot tout ce qui crée la vogue autour de l’imaginaire épigénétique » (Meloni et Testa 2014, p. 439), pourront être détectés et évités (Juengst et al., 2014; Joly et al., 2016).
Implications méthodologiques pour la recherche
La superposition nature-culture semble ouvrir un horizon de possibilités pour la recherche à l’intersection des sciences naturelles et des sciences sociales. La juxtaposition de perspectives disciplinaires autrefois isolées, dorénavant étroitement interreliées, interactives et complémentaires suggérerait l’avènement d’une pensée « post-dualiste » et d’une « logique de co-production » des savoirs (Meloni, 2014b).
In particular, the co-productionist framework will allow to unpack how the processes of gathering, standardizing and certifying epigenetic evidence will align with political, legal and economic rationalities in bringing about new settlements (or possibly reinforcing existing ones) across some of the most persistent dichotomies that structure our reflection on the human experience: normal versus pathological (or enhanced), safe versus dangerous, natural versus artificial, individual versus collective
Meloni et Testa, 2014, p. 445
Tel que mentionné précédemment, la précaution serait toutefois de mise face à la menace de nouvelles formes de réductionnisme biologique, ou « réductionnisme somatique », découlant de la description simpliste de phénomènes hautement complexes, comme la justice sociale, par la variation de marques épigénétiques :
evidence of neoreductionism is already apparent in the practice of environmental epigenetics in which the majority of researchers are busy tracking molecular pathways conceptualized as mediators between nurture and nature. In these carefully controlled experiments, “miniaturized environments” (Landecker and Panofsky 2013) proximal to the body are in effect recognized as equivalent to the nurture concept
Lock, 2015, p. 151
Un paradoxe semble donc émerger entre la possibilité de réconciliation nature-culture offerte par l’épigénétique et les risques de dérives associés à la biologisation du social (Dupras et Ravitsky, 2016b). Comme l’observe Meloni (2014a), « l’importation des connaissances en biologie vers les sciences humaines et sociales contribue souvent à renforcer l’idée fondatrice selon laquelle la biologie vient toujours en premier dans la chaîne des facteurs causaux (ce qui est l’essence du biologisme) » (p. 742). Contrer un tel danger de réification de la nature aux dépens de la culture serait cependant possible, entre autres, en ne surévaluant pas le pouvoir épistémique de la biologie dans l’explication des phénomènes de la vie (Pickersgill, 2016).
La plupart des initiatives internationales de recherche en épigénétique, par exemple le NIH Roadmap Epigenomics Project et l’International Human Epigenome Consortium (IHEC), apparaissent modelées directement sur la méthode de cartographie et séquençage (map and sequence) couramment utilisée en génétique : comme si les modifications épigénétiques étaient, elles aussi, « binaires, statiques et stables » (Meloni, 2014a). Dupras et Ravitsky (2016b) soulignent qu’il est possible que les intérêts technoscientifiques, politiques et commerciaux motivant ces programmes de recherche de grande envergure aient une influence sur les méthodes adoptées ainsi que sur la traduction des résultats de la recherche en interventions de santé.
It seems fair to say that in much of epigenomics, mapping currently proceeds the way it does primarily because the sequencing consortia’s intrinsic logic and dynamic dictates this direction. It is a direction where clinical application and viable markets appear most likely.
Niewöhner, 2011, p. 282-283
Une des conséquences de ces motivations sous-jacentes serait que la grande majorité des études en cours en épigénétique laisseraient trop peu de place aux zones de gris. Pour Beck et Niewöhner (2006), les chercheurs préfèrent le plus souvent déterminer à l’avance des indicateurs biochimiques simples, afin de faciliter la reconnaissance de variations statistiquement significatives à la base du développement d’outils diagnostic ou d’interventions cliniques standardisés. Ainsi, pour ces auteurs, la conduite même de la recherche en épigénétique procéderait d’une forme de « réductionnisme pragmatique »[13].
Quelques exceptions existeraient toutefois. Une étude ethnographique, conduite dans un laboratoire d’épigénétique environnementale au Canada, aurait permis à Niewöhner (2011) d’observer et de décrire un changement méthodologique important dans la façon de mener certaines recherches en biologie. Dans ce laboratoire, la variation biologique ne serait plus simplement réduite à un saut incrémental substantiel, mais comprise comme une modulation fine à la hausse ou à la baisse. On s’y intéresserait aux effets biologiques les plus subtils, ainsi qu’à la variabilité de la réponse épigénétique entre les différentes cellules d’un même tissu.
Significance, then, is not produced through a singular statistical measure but rather builds up in layers of analysis and interpretation. […] Producing thick significance is not something that the group chooses to do. In many ways, it is an instance of “reality kicking back” that forces them to open up the experimental system (Barad 1998). Asking questions that call up organismic and environmental contexts confronts them with the fact that complex phenomena such as social behaviour are difficult to control
Niewöhner, 2011, p. 288
La recherche en épigénétique environnementale pourrait, semble-t-il, exiger une approche quelque peu renouvelée de la validité scientifique d’un résultat. Pour Niewöhner (2011), la méthode expérimentale à privilégier, désormais, serait une sorte de « customary biology » n’ayant plus comme prémisse l’existence prédéterminée d’une norme naturelle, mais bien une relation intime entre nature et culture.
Customary biology is a biology based on patterns of practice and regularities rather than “natural” laws; a biology (and a biomedicine) that is attuned to investigations of bodies, which are being used by people in culturally specific ways to which they have grown accustomed over time; and a biology that may be able to productively engage anthropology and social inquiry and vice versa
Niewöhner, 2011, p. 293-294
La concrétisation d’alliances constructives entre chercheurs en sciences biologiques et en sciences sociales exige des ressources financières considérables. Pour Panofsky, elle suppose également une réforme des infrastructures et la création de véritables équipes multidisciplinaires, « de façon à dépasser la division disciplinaire du travail (les biologistes dans le laboratoire, les chercheurs en sciences sociales évaluant l’environnement) » (2015, p. 1106).
Une adaptation du concept de « biologie locale »[14] a été proposée par Niewöhner (2011) comme repère épistémologique pour l’étude du caractère dynamique et des conditions particulières de la programmation épigénétique. Ce repère résiderait dans un processus méthodologique nouveau, nommé « localizing biology », encourageant les chercheurs en sciences sociales, notamment, à éviter de déconstruire systématiquement l’aspect universel de certains phénomènes biologiques. Par exemple, si la programmation épigénétique des gènes n’est pas identique chez toutes les personnes, les mécanismes épigénétiques, eux, semblent largement partagés au sein de l’espèce humaine. Pour Niewöhner (2011), c’est en reconnaissant les objectifs, les méthodes et la contribution des différents types d’expertise impliqués qu’une « co-laboration » constructive pourrait être établie entre chercheurs de divers horizons.
[…] localizing biology must not be understood as a normative commitment and demand from anthropology toward biology. It is not in this sense an interdisciplinary agenda to change biology. Rather it is an empirical question that has the ontological status of human biology as its research object. […] Co-laboration[15] is temporary joint epistemic work. It is non-teleological in character. It does not require a shared outcome. It is an undertaking with a disciplinary vanishing point that requires participants from different epistemic cultures
Niewöhner, 2015, p. 234-235
La co-laboration suggérée par Niewöhner se distingue de la collaboration et de l’interdisciplinarité en ce sens qu’elle reconnaît que tous les partis n’ont pas les mêmes objectifs. Afin de prévenir diverses formes de réductionnisme, tous doivent cependant adopter un regard réflexif sur leur propre posture disciplinaire. La proximité et la communication entre les membres de diverses disciplines seraient par ailleurs des conditions favorables à cette autocritique.
Selon Lappé et Landecker (2015), il est toutefois possible, ironiquement, qu’une division dépassée du travail en silos disciplinaires soit renforcée par l’épigénétique. Pour ces auteurs, l’épigénétique est trop souvent définie et mobilisée comme champ d’études distinct par rapport au champ de la génétique. Ce dualisme génétique-épigénétique pourrait conduire l’épigénétique à se refermer sur elle-même, à exagérer ses particularités et à minimiser ses ambiguïtés inhérentes (Dupras et Ravitsky, 2016a).
We noted that the distinction actually was working as a kind of cushion, allowing a separate space of experimentation and explanation from that of genetics and genomics, allowing the genome to remain invariant while the epigenome varied above it and around it. In a sense, the separation allowed investigators to hold the genome still (which was not hard, because it had been conceptualized as such) while opening shop just next door
Lappé et Landecker, 2015, p. 169-170
Ainsi, les chercheurs en épigénétique apparaissent-ils eux-mêmes vulnérables à l’isolement et à toutes sortes de réductionnisme méthodologique. Une co-laboration active et soutenue entre chercheurs provenant de plusieurs disciplines pourrait contribuer à minimiser les conséquences indésirables de simplifications abusives pouvant survenir dans la conduite de la recherche, la diffusion des savoirs et l’application des connaissances en épigénétique.
Parties annexes
Notes
-
[1]
Nous entendons par « externe » tout ce qui se trouve à l’extérieur des frontières physiques du corps biologique.
-
[2]
Si la démarcation entre l’étude de la nature et l’étude de la société est très ancienne, l’opposition entre les concepts « nature » et « culture » (« nature and nurture ») a été formalisée vers la fin du xixe siècle par le père de l’eugénisme, Francis Galton (1822-1911). Elle devait lui servir à distinguer les traits naturels nobles, qu’il fallait favoriser pour améliorer l’humanité, des traits naturels indésirables pouvant nuire à la survie de l’espèce. Plusieurs observateurs se sont opposés à une telle distinction au cours de l’histoire. C’est le cas de l’anthropologue Franz Boas (1858-1942) qui, déjà au début du xxe siècle, soulevait l’importance de l’environnement dans l’apparition du phénotype. Aujourd’hui, plusieurs auteurs comme l’écrivaine féministe Evelyn Fox-Keller continuent de critiquer vigoureusement une telle dichotomie (Lock, 2013).
-
[3]
La « théorie synthétique de l’évolution » est la perspective dominante en biologie évolutive. Aussi reconnue comme la théorie néodarwinienne, elle consiste en une combinaison des théories de la sélection naturelle de Charles Darwin, des lois de transmission familiale des traits héréditaires de Gregor Mendel et de la génétique populationnelle. À ces travaux s’est aussi ajoutée la génétique moléculaire de la seconde moitié du xxe siècle, de la révélation de la double hélice d’ADN par Francis Crick et James Watson en 1953 au séquençage du génome humain en 2003. La vision néodarwinienne de l’évolution soutient que l’unité héréditaire est une chose « solide » et fixe dans le temps. Selon la théorie synthétique de l’évolution, cette unité héréditaire principale – en l’occurrence le gène – n’est pas elle-même influencée par l’environnement (Richards, 2006).
-
[4]
Notons que la différence entre les abeilles travailleuses et la reine n’est pas génétique, mais bien épigénétique. C’est l’alimentation de la reine durant son développement (gelée royale) qui modifie l’expression de ses gènes et lui confère des traits phénotypiques distinctifs et sa position sociale unique (Jablonka et Lamb, 2002).
-
[5]
La « construction de niche » est une théorie selon laquelle la conservation des espèces au cours de l’évolution n’est pas seulement le fruit de la constitution biologique privilégiée (ou le « fitness ») de certaines d’entre elles, mais aussi celui de la façon dont certaines espèces modifient leur environnement. Chez l’humain, le développement d’une diversité d’institutions afférées entre autres à la protection sociale et à la santé publique est souvent perçu comme une construction de niches favorable à la survie de l’espèce (Laland et al., 2001).
-
[6]
Jean-Baptiste Lamarck (1744-1829) est un théoricien de l’évolution pré-darwinien qui soutenait que la survie de certaines espèces serait favorisée par des mécanismes d’adaptation à leur environnement. Ces mécanismes leur permettraient d’amplifier ou d’atrophier des traits biologiques pour mieux répondre aux conditions de vie. Ces traits favorables acquis pourraient ensuite, à l’occasion, être transmis aux générations suivantes. Cette théorie de l’hérédité « souple », longtemps discréditée par les travaux de Darwin sur la sélection naturelle de composantes biologiques acquises plutôt de façon aléatoire (mutations favorables de l’ADN), pourrait « renaître » aujourd’hui à la lumière des travaux en épigénétique sous l’appellation néo-lamarckisme (Richards, 2006).
-
[7]
Le biologiste August Weismann (1834-1914) réfuta la théorie lamarckienne de l’évolution sur la base de la distinction entre les cellules germinales et somatiques. Puisqu’une cellule différenciée ne peut pas « retourner en arrière » et retrouver la totipotence des cellules germinales, il est peu probable que les fonctions acquises par les cellules somatiques soient perpétuées jusqu’après la procréation qui, elle, se produit nécessairement à travers les cellules germinales (Richards, 2006).
-
[8]
Dans une perspective évolutive, la transmission directe pourrait, en fait, être plus avantageuse que la transmission transgénérationnelle, puisqu’elle permet une plus grande plasticité et réversibilité de la programmation épigénétique et confère ainsi une plus grande capacité d’adaptation des organismes aux conditions de vie parfois différentes entre les générations (Thayer et Non, 2015).
-
[9]
Trofim Denisovich Lysenko (1898-1976) est un agronome russe tristement célèbre et désavoué de façon générale par la communauté scientifique internationale pour avoir promu une version extrême du lamarckisme – et du même coup avoir nié l’importance de l’hérédité génétique – à des fins politiques et nationalistes dans les années 1930 à 1960 pour le gouvernement de l’Union soviétique (Richards, 2006).
-
[10]
Les discussions sur les limites du déterminisme génétique ont mené le chercheur en biosémiotique Jesper Hoffmeyer (2002) à proposer le concept de « fallacy of digitalism », qu’il définit comme « the more or less automatic preference for explanations that ascribe primacy to digitally coded information or see such information as an explanation bottom line » (Burbano, 2006, p. 854).
-
[11]
Les prémisses des certaines théories de la justice, par exemple, ont été discutées par plusieurs auteurs à la lumière des progrès récents en épigénétique (Stapleton et al., 2012; Loi et al., 2013; Lafaye, 2015).
-
[12]
Dans son ouvrage Gender Trouble, publié en 1990, la féministe Judith Butler définit le genre comme « the repeated stylization of the body, a set of repeated acts ... that congeal over time to produce the appearance of substance, of a natural sort of being ». Elle s’oppose au genre comme « catégorie “naturelle” objective » et soutient qu’il est le fruit de l’adhésion, au quotidien, à des pratiques et à des discours (Osborne, 2015).
-
[13]
Réductionnisme pragmatique : « Linear understandings of physiological change underpin most explanations of disease aetiology. For most complex or systemic diseases, these are rather crude, mechanistic perspectives that guide clinical practice rather than claim to fully represent (patho)physiology. This pragmatic reductionism serves medical science well as it enables the assembly of boundaries around objects of investigation » (Beck et Niewöhner, 2006, p. 223).
-
[14]
Le concept de « local biology » a été proposé en 1993 par l’anthropologue Margaret Lock à l’issue d’une série d’études sur l’influence de l’environnement sur les symptômes de la ménopause (Niewöhner, 2011).
-
[15]
L’utilisation du néologisme « co-laboration », d’abord en anglais, puis repris en français, est justifiée ainsi : « [Collaboration is] problematic as it conjures up the problematic notion of the collaborateur, that is, the one who cooperates with the enemy – not necessarily the prototype of a participant in a shared epistemic process. Collaboration is also based on shared intentions and goals between researcher and the one formerly known as informant » (Niewöhner, 2015, p. 236).
Bibliographie
- Bateson, Patrick, « Evolution, epigenetics and cooperation », Journal of Biosciences, vol. 39, 2014, p. 191-200.
- Beck, Stefan et Jörg Niewöhner, « Somatographic investigations across levels of complexity », BioSocieties, vol. 1, no. 2, 2006, p. 219-227.
- Boniolo, Giovanni et Giuseppe Testa, « The identity of living beings, epigenetics, and the modesty of philosophy », Erkenntnis, vol. 76, no. 2, 2012, p. 279-298.
- Bossdorf, Oliver, Christina L. Richards et Massimo Pigliucci, « Epigenetics for ecologists », Ecology Letters, vol. 11, 2008, p. 106-115.
- Brooke, John L. et Clarck Spencer Larsen, « The nurture of nature: Genetics, epigenetics, and environment in human biohistory », The American Historical Review, vol. 119, no. 5, 2014, p. 1500-1513.
- Burbano, Hernán A., « Epigenetics and genetic determinism », História, Ciências, Saúde-Manguinhos, vol. 13, no. 4, 2006, p. 851-863.
- Canning, Christopher, « Epigenetics: An emerging challenge to genetic determinism in studies of mental health and illness », Social Alternatives, vol. 27, no. 4, 2008, p. 14-21.
- Chiapperino, Luca et Giuseppe Testa, « The epigenomic self in personalized medicine: between responsibility and empowerment », The Sociological Review Monographs, vol. 64, no. 1, 2016, p. 203-220.
- Davis, Noela, « Politics materialized: Rethinking the materiality of feminist political action through epigenetics », Women: A Cultural Review, vol. 25, no. 1, 2014, p. 62-77.
- Diemer, Kendall et Melissa Woghiren, « Epigenetic privacy: Hacking your health? » dans Michael Crawford (dir.), Epigenetics in Society, Windsor, Epigenetics Study Group, 2015.
- Drake, Amanda et Lincoln Liu, « Intergenerational transmission of programmed effects: public health consequences », Trends in Endocrinology & Metabolism, vol. 21, no... 4, 2010, p. 206-213.
- Dupras, Charles et Vardit Ravitsky, « The ambiguous nature of epigenetic responsibility », Journal of Medical Ethics, vol. 42, no. 8, 2016a, p. 534-541.
- Dupras, Charles et Vardit Ravitsky, « Epigenetics in the neoliberal “regime of truth”: A biopolitical perspective on knowledge translation », Hastings Center Report, vol. 46, no. 1, 2016b, p. 26-35.
- Dupras, Charles, Vardit Ravitsky et Bryn Williams-Jones, « Epigenetics and the environment in bioethics », Bioethics, vol. 28, no. 7, 2014, p. 327-334.
- Dyke, Stephanie O. M., et. al., « Epigenome data release: A participant-centered approach to privacy protection », Genome Biology, vol. 16, no. 1, 2015, p. 142-154.
- Gadjev, Ilya, « Epigenetics, Representation, and Society », Zygon, vol. 52, no. 2, 2017, p. 491-515.
- Gershon, Naomi B. et Pamela C. High, « Epigenetics and child abuse: Modern day Darwinism: The miraculous ability of the human genome to adapt, and then adapt again », American Journal of Medical Genetics Part C: Seminars in Medical Genetics, vol. 169, no. 4, 2015, p. 353-360.
- Godfrey, Keith M., Karen M. Lillycrop, Graham C. Burdge, Peter D. Gluckman et Mark A. Hanson, « Epigenetic mechanisms and the mismatch concept of the developmental origins of health and disease », Pediatric Research, vol. 61, 2007, p. 5-10.
- Gonon, François et Marie-Pierre Moisan, « L’épigénétique, la nouvelle biologie de l’histoire individuelle? », Revue française des affaires sociales, vol. 1, 2013, p. 21-31.
- Guthman, Julie, « Doing justice to bodies? Reflections on food justice, race, and biology », Antipode, vol. 46, no. 5, 2012, p. 1153-1171.
- Guthman, Julie et Becky Mansfield, « The implications of environmental epigenetics: A new direction for geographic inquiry on health, space, and nature-society relations », Progress in Human Geography, vol. 37, no. 4, 2012, p. 486-504.
- Haig, David, « Weismann rules! OK? Epigenetics and the Lamarckian temptation », Biology & Philosophy, vol. 22, no. 3, 2007, p. 415-428.
- Heard, Edith et Robert A. Martienssen, « Transgenerational epigenetic inheritance: Myths and mechanisms », Cell, vol. 157, no. 1, 2014, p. 95-109.
- Hendrickx, Kim et Ine van Hoyweghen, « An epigenetic prism to norms and values », Frontiers in Genetics, vol. 9, 2018, p. 1-5.
- Herb, Brian R., « Epigenetics as an answer to Darwin’s “special difficulty” », Frontiers in genetics, vol. 5, 2014, p. 1-5.
- INSERM, Dossiers d’information : Épigénétique, Paris, INSERM, 2015, http://www.inserm.fr/thematiques/genetique-genomique-et-bioinformatique/dossiers-d-information/epigenetique (consulté le 18 janvier 2017).
- Jablonka, Eva et Marion J. Lamb, « The changing concept of epigenetics », Annals of the New York Academy of Sciences, vol. 981, no. 1, 2002, p. 82-96.
- Joly, Yann, Derek So, Katie Saulnier et Stéphanie O. M. Dyke, « Epigenetics ELSI: Darker than you think? », Trends in Genetics, vol. 32, no. 10, 2016, p. 591-592.
- Juengst, Eric T., Jennifer R. Fishman, Michelle L. McGowan et Richard A. Settersten, « Serving epigenetics before its time », Trends in Genetics, vol. 30, no. 10, 2014, p. 427-429.
- Katz, Michael B., « The biological inferiority of the undeserving poor », Social Work & Society, vol. 11, no. 1, 2013, p. 1-9.
- Kuzawa, Christopher W. et Elizabeth Sweet, « Epigenetics and the embodiment of race: Developmental origins of US racial disparities in cardiovascular health », American Journal of Human Biology, vol. 21, no. 1, 2009, p. 2-15.
- Kuzawa, Christopher W. et Zenata M. Thayer, « Timescales of human adaptation: The role of epigenetic processes », Epigenomics, vol. 3, no. 2, 2011, p. 221-234.
- Lafaye, Caroline Guibet, « Quelle théorie de la justice pour l’épigénétique? », Dialogue: Canadian Philosophical Review, vol. 54, 2015, p. 489-517.
- Laland, Kevin N., J. Odling-Smee et Marcus W. Feldman, « Cultural niche construction and human evolution », Journal of Evolutionary Biology, vol. 14, no. 1, 2001, p. 22-33.
- Landecker, Hannah, « Food as exposure: Nutritional epigenetics and the new metabolism », BioSocieties, vol. 6, no. 2, 2010, p. 167-194.
- Landecker, Hannah et Aaron Panofsky, « From social structure to gene regulation, and back: A critical introduction to environmental epigenetics for sociology », Annual Review of Sociology, vol. 39, 2013, p. 333-357.
- Lappé, Martine, « Epigenetics, Media Coverage, and Parent Responsibilities in the Post-Genomic Era », Current Genetic Medicine Reports, vol. 4, no. 3, 2016a, p. 92-97.
- Lappé, Martine, « The maternal body as environment in autism science », Social Studies of Science, vol. 46, no. 5, 2016b, p. 675-700.
- Lappé, Martine et Hannah Landecker, « How the genome got a life span », New Genetics and Society, vol. 34, no. 2, 2015, p. 152-176.
- Lerner, R. M. et Willis F. Overton, « Epigenetics, evolution, and embodiment: On the conceptual vacuity of evolutionary psychology », Open Access Genetics, vol. 1, 2014, p. 6-10.
- Lock, Margaret, « The epigenome and nature/nurture reunification: A challenge for anthropology », Medical Anthropology, vol. 32, no. 4, 2013, p. 291-308.
- Lock, Margaret, « Comprehending the body in the era of the epigenome », Current Anthropology, vol. 56, no. 2, 2015, p. 151-177.
- Loi, Michele, Lorenzo Del Savio et Elia Stupka, « Social epigenetics and equality of opportunity », Public Health Ethics, vol. 6, no. 2, 2013, p. 142-153.
- Mansfield, Becky, « Race and the new epigenetic biopolitics of environmental health », BioSocieties, vol. 7, no. 4, 2012, p. 352-372.
- Mansfield, Becky et Julie Guthman, « Epigenetic life: Biological plasticity, abnormality, and new configurations of race and reproduction », Cultural Geographies, vol. 22, no. 1, 2015, p. 3-20.
- Meloni, Maurizio, « Biology without biologism: Social theory in a postgenomic age », Socio-logy, vol. 48, no. 4, 2014a, p. 731-746.
- Meloni, Maurizio, « How biology became social, and what it means for social theory », The Sociological Review, vol. 62, no. 3, 2014b, p. 593-614.
- Meloni, Maurizio, « Epigenetics for the social sciences: Justice, embodiment, and inheritance in the postgenomic age », New Genetics and Society, vol. 34, no. 2, 2015a, p. 125-151.
- Meloni, Maurizio, « Heredity 2.0: The epigenetics effect », New Genetics and Society, vol. 34, no. 2, 2015b, p. 117-124.
- Meloni, Maurizio, « Race in an epigenetic time: Thinking biology in the plural », The British Journal of Sociology, vol. 68, no. 3, 2017, p. 389-409.
- Meloni, Maurizio et Giuseppe Testa, « Scrutinizing the epigenetics revolution », BioSocieties, vol. 9, no. 4, 2014, p. 431-456.
- Middleton, Peter, « Epigenetics and poetry: Challenges to genetic determinism in Michael Byers’ Long for This World, and Mei-mei Berssenbrugge’s “The Four Year Old Girl” », Textual Practice, vol. 29, no. 3, 2015, p. 517-545.
- Müller, Ruth, et. al., « The biosocial genome?: Interdisciplinary perspectives on environmental epigenetics, health and society », EMBO Reports, 2017, e201744953.
- Niewöhner, Jörg, « Epigenetics: Embedded bodies and the molecularisation of biography and milieu », BioSocieties, vol. 6, no. 3, 2011, p. 279-298.
- Niewöhner, Jörg, « Epigenetics: Localizing biology through co-laboration », New Genetics and Society, vol. 34, no. 2, 2015, p. 219-242.
- Osborne, Jim, « Getting under performance’s skin: Epigenetics and gender performativity », Textual Practice, vol. 29, no. 3, 2015, p. 499-516.
- Panofsky, Aaron, « Commentary: A conceptual revolution limited by disciplinary division », International Journal of Epidemiology, vol. 44, no. 4, 2015, p. 1105-1107.
- Paul, Diane B., The Politics of Heredity: Essays on Eugenics, Biomedicine, and the Nature-Nurture Debate, Ithaca (NY), SUNY Press, 1998.
- Pickersgill, Martyn, « Neuroscience, epigenetics and the intergenerational transmission of social life: Exploring expectations and engagements », Families, Relationships and Societies, vol. 3, no. 3, 2014, p. 481-484.
- Pickersgill, Martyn, « Epistemic modesty, ostentatiousness and the uncertainties of epigenetics: On the knowledge machinery of (social) science », The Sociological Review Monographs, vol. 64, no. 1, 2016, p. 186-202.
- Pickersgill, Martyn, Jörg Niewöhner, Ruth Müller, Paul Martin et Sarah Cunningham-Burley, « Mapping the new molecular landscape: Social dimensions of epigenetics », New Genetics and Society, vol. 32, no. 4, 2013, p. 429-447.
- Rial-Sebbag, Emmanuelle, Caroline Guibet Lafaye, Umberto Simeoni et Claudine Junien, « DOhaD and epigenetic information: Societal challenges », Médecine / Sciences, vol. 32, no. 1, p. 100-105.
- Richards, Eric J., « Inherited epigenetic variation: Revisiting soft inheritance », Nature Reviews Genetics, vol. 7, no. 5, 2006, p. 395-401.
- Richardson, Sarah S., « Maternal bodies in the postgenomic order: Gender and the explanatory landscape of epigenetics », dans Sarah Richardson et Hallam Stevens (dir.), Postgenomics Perspective on Biology after the Genome, Durham (NC), Duke University Press, 2015.
- Rothstein, Mark A., « Epigenetic exceptionalism », The Journal of Law, Medicine & Ethics, vol. 41, no. 3, 2013, p. 733-736.
- Rothstein, Mark A., Yu Cai et Gary E. Marchant, « The ghost in our genes: Legal and ethical implications of epigenetics », Health Matrix, vol. 19, no. 1, 2009, p. 1-62.
- Rothstein, Mark A., Heather L. Harrell et Gary E. Marchant, « Transgenerational epigenetics and environmental justice », Environmental Epigenetics, vol. 3, no. 3, 2017, p. 1-12.
- Sagl, Veronika, Roman Thaler, Astrid Gesche et Alexander G. Haslberger, « New understanding of epigenetics and consequences for environmental health and sustainability », Sustainable food production and ethics EurSafe 2007 (7th Congress of the European Society for Agriculture and Food Ethics, Vienne (Autriche), 2007, p. 376-381.
- Saulnier, Katie M. et Charles Dupras, « Race in the postgenomic era: Social epigenetics calling for interdisciplinary ethical safeguards », The American Journal of Bioethics, vol. 17, no. 9, 2017, p. 58-60.
- Singh, Ilina, « Human development, nature and nurture: Working beyond the divide », BioSocieties, vol. 7, no. 3, 2012, p. 308-321.
- Stapleton, Greg, Peter Schröder-Bäck et David Townend, « Equity in public health: An epigenetic perspective », Public Health Genomics, vol. 16, no. 4, 2012, p. 135-144.
- Stelmach, Aleksandra et Brigitte Nerlich, « Metaphors in search of a target: The curious case of epigenetics », New Genetics and Society, vol. 34, no. 2, 2015, p. 196-218.
- Sullivan, Shannon, « Inheriting racist disparities in health: Epigenetics and the transgenerational effects of white racism », Critical Philosophy of Race, vol. 1, no. 2, 2013, p. 190-218.
- Thayer, Zenata M. et Amy L. Non, « Anthropology meets epigenetics: Current and future directions », American Anthropologist, vol. 117, no. 4, 2015, p. 722-735.
- Tolwinski, Kasia, « A new genetics or an epiphenomenon? Variations in the discourse of epigenetics researchers », New Genetics and Society, vol. 32, no. 4, 2013, p. 366-384.
- Valdez, Natali, « The Redistribution of Reproductive Responsibility: On the Epigenetics of “Environment” in Prenatal Interventions », Medical Anthropology Quarterly, 2018, https://doi.org /10.1111/maq.12424.
- Van de Vijver, Gertrudis, Linda Van Speybroeck et Dani de Waele, « Epigenetics: A challenge for genetics, evolution, and development? », Annals of the New York Academy of Sciences, vol. 981, no. 1, 2002, p. 1-6.
- Waggoner, Miranda R. et Tobias Uller, « Epigenetic determinism in science and society », New Genetics and Society, vol. 34, no. 2, 2015, p. 177-195.
- Wallack, Lawrence et Kent Thornburg, « Developmental origins, epigenetics, and equity: Moving upstream », Maternal and Child Health Journal, vol. 20, no. 5, 2016, p. 935-940.
- Warin, Megan, Vivienne Moore, Michael Davies et Stanley Ulijaszek, « Epigenetics and obesity: The reproduction of habitus through intracellular and social environments », Body & Society, vol. 22, no. 4, 2015, p. 53-78.
- Whitelaw, Emma, « Disputing Lamarckian epigenetic inheritance in mammals », Genome Biology, vol. 16, no. 1, 2015, p. 60.
Liste des figures
Figure 1
Dissection du rapprochement nature-culture
Perspectives distinctes et complémentaires sur les implications épistémologiques potentielles de l’épigénétique : 1) socialisation du biologique, 2) biologisation du social et 3) superposition nature-culture.