Abstracts
Abstract
The silverleaf whitefly (SLW), Bemisia tabaci, is an exotic pest that causes sporadic crop damage in Canadian greenhouses, mainly on tomatoes. Its plant feeding is responsible for both direct damage as well as the transmission of plant viruses. Interestingly, omnivorous predators, which are used as biological control agents for multiple whitefly species, can establish on greenhouse crops even before the pest has established, using alternative food resources and host plants to support the predator. However, the efficiency of such indigenous omnivorous biological agents in the suppression of high density and intense SLW infestations remains unknown. This research aimed to assess the potential of two indigenous omnivorous biological agents, Dicyphus hesperus and Orius insidiosus to control SLW infestations and their effects on tomato plant health and yield at both early stages of the crop cycle, and at high pest densities. This study showed that populations of B. tabaci are able to grow rapidly. However, neither of the introduced densities of B. tabaci nor the infestation time had a significant effect on the incidence of tomato irregular ripening disorder (TIR) and yield. Our results also show that the introduction of three or five D. hesperus adults per cage reduces the number of B. tabaci larvae and pseudo-nymphs, while O. insidiosus did not significantly reduce SLW populations. Our results suggest that D. hesperus could control high levels of SWL and at an early stage in the cropping cycle.
Keywords:
- Bemisia tabaci,
- Dicyphus hesperus,
- Orius insidiosus,
- biological control,
- greenhouse,
- tomato irregular ripening disorder
Résumé
L’aleurode du tabac (SLW), Bemisia tabaci, est un ravageur exotique qui cause des dommages sporadiques dans les serres canadiennes, principalement en culture de tomates. Son comportement alimentaire est responsable de dommages directs et de la transmission de virus. Les prédateurs omnivores peuvent s’établir en serre avant l’arrivée des ravageurs, en utilisant des proies et hôtes alternatifs. Toutefois, l’efficacité des prédateurs indigènes contre B. tabaci reste inconnue. Cette étude avait pour objectif de tester la réponse des plants de tomate aux infestations hâtives et de fortes intensités de B. tabaci, et le potentiel de deux punaises omnivores indigènes, Dicyphus hesperus et Orius insidiosus. Les résultats démontrent que les populations de B. tabaci augmentent rapidement. Cependant, ni la densité ni le temps d’infestation n’avaient d’effet significatif sur le trouble du désordre de maturation (TIR) et sur le rendement. L’introduction de trois et cinq D. hesperus adultes réduisait le nombre de larves et nymphes de B. tabaci, alors que l’utilisation d’O. insidiosus n’a pas permis de réduire significativement les populations. Nos résultats suggèrent que D. hesperus pourrait réguler de fortes et brèves infestations de B. tabaci.
Mots-clés :
- Bemisia tabaci,
- Dicyphus hesperus,
- Orius insidiosus,
- contrôle biologique,
- serres,
- trouble du désordre de maturation de la tomate
Article body
INTRODUCTION
The silverleaf whitefly (SLW), also called the sweet potato whitefly, Bemisia tabaci (Gennadius, 1889) (Hemiptera: Aleyrodidae), is among the most common and globally distributed of crop pests (Castañé et al. 2020; Oliveira et al. 2001). It has been found to attack at least 600 plant species, including cotton, poinsettia, sweet potato, or squash, and is a severe pest of several greenhouse crops including tomato, pepper, beans, eggplant, and cucumber (Cook 1986; Oliveira et al. 2001). In Quebec, the SLW is generally less frequently observed than the greenhouse whitefly Trialeurodes vapora-riorum (Westwood, 1856), but its infestations have increased in recent years. The SLW represents a threat to greenhouse crops, particularly to tomato, since this species transmits over 90 viruses and generates plant physiological disorders (De Barro et al. 2011; Jones 2003; McAuslane and Smith 2018). In tomato crops, the SLW (biotype B) induces a ripening disorder, first detected in Florida (USA) in 1987 (McCollum et al. 2004), which caused losses of $25 million for producers of Florida in 1989 (McAuslane and Smith 2018). This physio-logical disorder, called tomato irregular ripening (TIR), disturbs the ripening of tomatoes by inhibiting fruit softening, that is to say, that fruit remains green and firm in the centre (McCollum et al. 2004; McKenzie and Albano 2009; Schuster et al. 1990). As few as 0.5 individuals per leaf for a short period of time can induce TIR (Schuster et al. 2019). Control methods, such as pesticides or biological control, therefore have to be applied in a timely manner to reduce whitefly density to below this threshold (Smith and Krey 2019). Consequently, growers often rely on frequent pesticide applications for their control needs (Calvo, Torres-Ruiz et al. 2018). However, the development of pesticide resistance in whitefly is well documented for many groups of pesticides (Bass et al. 2015; Cahill et al. 1996; Horowitz et al. 1999; Nauen and Denholm 2005; Palumbo et al. 2001; Prabhaker et al. 1985). As such, a number of biological control organisms have been identified for controlling the SLW including parasitoids and predators.
Natural enemies of the SLW include predators such as lady beetles (Coleoptera: Coccinellidae), predatory bugs (Heteroptera: Miridae and Anthocoridae), lacewings (Neuroptera: Chrysopidae and Coniopterygidae), mites (Acari: Phytoseiidae), and spiders (Araneae) (Gerling et al. 2001). In particular, commercially available biological agents include Amblyseius swirskii Athias-Henriot, 1962 (Acari), Delphastus catalinae (Horn, 1895) (Coleoptera: Coccinellidae), Dicyphus hesperus (Knight, 1943), Nesidiocoris tenuis (Reuter, 1895), and Macrolophus pygmaeus (Rambur, 1839) (Heteroptera: Miridae), as well as several species of parasitoids, including those from genera Encarsia. and Eretmocerus (Hymenoptera: Aphelinidae) (Greenberg et al. 2002; Hoddle and Van Driesche 1999; van Lenteren 2012). However, A. swirskii and D. catalinae are less effective on tomato plants due to the presence of trichomes (Buitenhuis et al. 2014; Guershon and Gerling 2006).
In Canada, N. tenuis and M. pygmaeus are not commercially available because they are not native species (Calvo et al. 2016). Hence, the predatory bug D. hesperus and aphelinid parasitoid wasps remain the only available biocontrol agents in Canada with which to effectively manage whitefly. Interestingly, predatory bugs can be introduced in greenhouses before the whitefly population becomes problematic, by adding Ephestia eggs or mullein plants as supplemental food or ovipositional substrates respectively (Sanchez et al. 2003; Smith and Krey 2019), which is rarely done with parasitoid wasps.
Previous experiments have revealed that Dicyphus bugs are effective predators for controlling several pests in tomato crops, including mites, whiteflies (McGregor et al. 1999), and especially the SLW (Calvo et al. 2016; Calvo, Torres et al. 2018; Calvo, Torres-Ruiz et al. 2018; Smith and Krey 2019). Whiteflies represent a food of higher quality for D. hesperus, as feeding on whitefly is associated with a higher predator growth rate and a reduced development time compared to feeding on mites (McGregor et al. 1999). Moreover, mullein plants can serve as reservoirs for D. hesperus (Smith and Krey 2019). In addition to D. hesperus, the generalist bug Orius insidiosus, which feeds on several pests such as thrips and aphids, has also been reported to feed on whitefly (Coll and Ridgway 1995). Therefore, this species could also represent a potential to control agent of the SLW in tomato production. O. insidiosus also feeds on pollen, and the presence of this resource promotes its development and increases its longevity (Wong and Frank 2013). This anthocorid bug effectively controls pests on several crops (e.g., bean, corn, tomato, pepper, cucumber) (Coll and Ridgway 1995).
The present study aims to evaluate the potential of these two indigenous and omnivorous biological agents, D. hesperus and O. insidiosus on high level and short time infestation of SLW and its impact on TIR and yield. We first tested the impact of B. tabaci densities and infestation time on the rate of TIR. Then, the effect of omnivorous predators on B. tabaci density was tested.
MATERIALS AND METHODS
Insect populations
Silverleaf whiteflies used in experiments originated from a commercial tomato greenhouse collected during 2018 and 2019 summers and were reared in the lab at the CRAM research centre until having enough individuals to perform the experiments. Biotype B was identified by genetic methods by the Laboratoire d’expertise et de diagnostic en phyto-protection of the Ministère de l’Agriculture, des Pêcheries et de l’Alimentation du Québec (MAPAQ, Québec, Canada). Whiteflies were reared on healthy eggplants in cages in controlled environment chambers, maintained at 25 °C, a 60% relative humidity, and a photoperiod 16:8 h (light:dark). A colony of D. hesperus maintained at the Université du Québec à Montréal (UQAM) (source population from Anatis bioprotection inc., Saint-Jacques-le-Mineur, Québec), as well as a commercial source (Anatis bioprotection inc.) for O. insidiosus provided the predatory bugs for trials in this study. For all experiments, adult whiteflies were introduced in a mesh sleeve covering a tomato leaf on 10-leaf plants.
Experimental site
The experiments took place in a greenhouse of the Centre de recherche agroalimentaire de Mirabel (CRAM), located in Mirabel (Quebec, Canada) during summer 2018. In the greenhouse, four rows of 10-leaf tomato plants (without flowers at the start of the experiment) of variety Beef Starbuck (known to be sensitive to whiteflies) were planted (distance between plants of 0.5 m on a row and row spacing of 1 m). The same set-up was used for experiment 1 (the study of whitefly density and damages) and experiment 2 (effect of predators on whitefly density).
Experiment 1: Whitefly density and plant damage
The effects of varying whitefly infestation intensity (densities of 0, 10, 50 or 100 whiteflies per treatment) and the duration of infestation (one versus three weeks) on tomato production (harvest date, number of tomatoes produced, mean tomato weight) and the presence of the maturation disorder were assessed in a trial with a randomized complete block design (80 plants total or 10 plants per treatment). A muslin sleeve was put around one of the two superior leaves of each plant (fully developed leaf), and adult whiteflies were introduced at rates of 10, 50, or 100 whiteflies depending on the treatment. The sleeves were sealed just after the introduction of the whiteflies. Control treatments consisted of leaves surrounded by a sleeve but with no whitefly added. One week later, all the 1-week treatment leaves were cut, and whiteflies (eggs, larvae, pseudo-nymphs, adults) were counted in the lab using a binocular. Three weeks after the introduction of whiteflies, all the 3-week treatment leaves were cut, and whiteflies (eggs, larvae, pseudo-nymphs, adults) were observed and counted in the lab. All tomatoes produced by plants were harvested when mature (up to three weeks after the end of the experiment), weighed, and the presence of maturation disorder was recorded (presence or absence) (green patches either on the tomato fruit or inside).
Experiment 2: Effect of predators on whitefly density
The effect of predatory bugs D. hesperus and O. insidiosus on whitefly density was determined in trials with a randomized complete block design (100 plants total, or 10 plants per treatment). A muslin sleeve was put around one of the two superior leaves of each plant (fully developed leaf), and 50 adult whiteflies were introduced within (except for the Bemisia control treatment). The sleeves were sealed just after the introduction of whiteflies. One week later, depending on the treatment, 1, 3 or 5 individuals of D. hesperus or O. insidiosus were introduced into the sleeve cages. No predators were introduced onto control plants. Four weeks later, the leaves within sleeves were cut. Whiteflies (eggs, larvae, pseudo-nymphs, adults) and predatory bugs (immatures, adults) were counted in the lab.
Statistical analyses
Whitefly density, ripening disorder, and yield
Three generalized linear model (GLM) procedures for data with a Poisson distribution (corrected for overdispersion) were used to compare the densities of SLW eggs, larvae, pseudo-nymphs and adults as a function of initial SLW density. The factor “duration” was also included in the egg density model, but not in other models because there were no pseudo-nymphs and adults on week 1.
The effects of SLW density, duration of infestation, and the interaction between these factors on the proportion of ripening disorder, the yield (number of tomatoes per plant), and the total weight of tomatoes per plant was assessed using, respectively, a GLM procedure for the binomial distribution, a GLM for Poisson distribution and a linear regression model.
Effect of predators on whitefly density
The effect of the initial predator (either O. insidiosus or D. hesperus) density on SLW density (all stages pooled) was also assessed through an analysis of variance (ANOVA) and a post-hoc means separation Tukey-Kramer test. The significance level was set at alpha = 0.05.
All analyses were done in R (R Core Team 2017).
Figure 1
Trial average number of Bemisia tabaci eggs (± SE) (a) or nymphs and pupae (b) per leaf relative to initial whitefly introduction rate and duration of infestation

The number of eggs, larvae or pseudo-nymphs increased with the initial density of B. tabaci introduced. Different letters indicate statistical differences among treatments.
Figure 2
Average rate of tomato irregular ripening (TIR) disorder (± SE) observed in function of the Bemisia tabaci introduction rate and duration of infestation

The introduction rate and the duration of infestation had no significant effect on the rate of TIR.
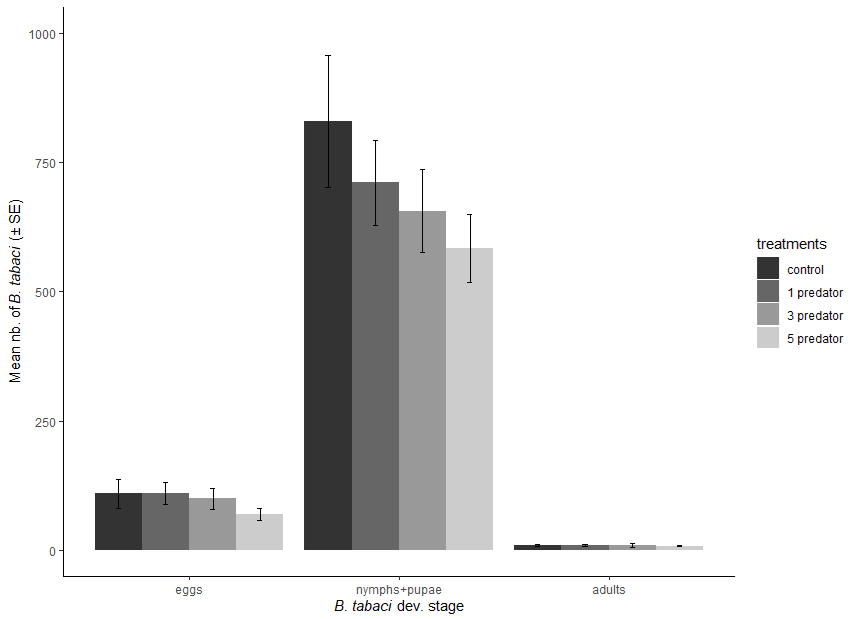
Figure 3
Trial average Bemisia tabaci density (± SE) in function of the density of Dicyphus hesperus

Statistical significance is indicated by different letters (α = 0.05).
RESULTS
Whitefly density, ripening disorder, and yield
Populations of the SLW were established very quickly on tomato plants, with a majority of eggs laid during the first week following the whiteflies’ introduction. In the third week, the whitefly population was mainly composed of larvae and pseudo-nymphs (Fig. 1). The number of eggs was lower over the period of three weeks (β = -0.51 ± 0.16, t = -3.24; P = 0.002). The number of eggs (t = 6.80, P < 0.0001), larvae or pseudo-nymphs (t = 2.79, P = 0.01) and adults (t = 2.70, P = 0.02) increased with the initial density of B. tabaci introduced.
In the control treatment, 8% (± 3% SE) tomatoes were classified as irregularly ripened. This rate was not affected by initial whitefly density (P = 0.17) nor duration of infestation (P = 0.29) (Fig. 2). No interaction between SLW initial density and infestation duration (P = 0.56) was observed.
The mean tomato yield and total tomato weight per plant were respectively 3.72 (± 1.88 SD) tomatoes per plant and 185.31 g (± 47.96 SD). Neither initial SLW density nor duration of infestation had an impact on tomato yield (density: P = 0.77, duration: P = 0.18) or total tomato weight per plant (density: P = 0.82, duration: P = 0.56). The interaction of SLW density and infestation duration had no effect on yield (P = 0.12) nor on weight of tomatoes (P = 0.17).
Effect of predators on whitefly density
Five weeks after introducing 50 adult whiteflies, 109.6 (± 27.7 SE) eggs, 829.3 (± 126.8 SE) larvae or pseudo-nymphs and 9.3 (± 1.6 SE) adult whiteflies were observed in the muslin sleeve in the control treatment. The introduction of three and five D. hesperus adults reduced the number of whiteflies larvae and pseudo-nymphs by 42.8% and 49.9%, respectively (F(3, 36) = 4.01, P = 0.01) (Fig. 3). The lower density of D. hesperus (one individual) had no significant effect on SLW larvae and pseudo-nymph populations. The introduction of D. hesperus did not reduce the number of SLW eggs (F(3, 36) = 0.86, P = 0.47) and adults (F(3, 36) = 1.68, P = 0.19) (Fig. 3).
The introduction of O. insidiosus had no significant effect on the density of SLW eggs (F(3, 36) = 0.86, P = 0.47), larvae or pseudo-nymphs (F(3, 36) = 1.29, P = 0.29) and adults (F(3, 36) = 0.12, P = 0.95) (Fig. 4).
Figure 4
Bemisia tabaci density (± SE) in function of the density of Orius insidiosus
The O. insidiosus introduction rate had no significant effect on B. tabaci density.
DISCUSSION
High level and short time infestation of SLW did not impede tomato irregular ripening disorder nor incur a yield reduction, even if the threshold of 0.5 nymphs/leaflet was largely reached. Globally, the predatory bug D. hesperus had a negative effect on the populations of SLW, whereas O. insidiosus did not significantly impact SLW’s populations. A decrease in the larvae and pseudo-nymph populations of B. tabaci by 42.8% and 49.9% was observed at an initial density of three and five D. hesperus for the initial 50 B. tabaci release treatment.
Whitefly density, ripening disorder, and yield
The impact of B. tabaci presence on ripening disorder in tomato plants has yet to be fully understood (McCollum et al. 2004; McKenzie and Albano 2009). Past work has found that SLW feeding suppresses ethylene biosynthesis in tomato fruit (McCollum et al. 2004), and this inhibition of ethylene biosynthesis disrupts ripening, which leads to the non-uniform colour and inhibition of fruit softening associated with the TIR disorder (Hamilton et al. 1991; McCollum et al. 2004; Oeller et al. 1991). Schuster (2002) also reported that the TIR could be induced by very low whitefly densities, as low as 5 nymphs per 10 leaflets (Schuster et al. 2019), while high densities can increase the severity of TIR (Schuster 2002; Schuster et al. 1990). In contrast, TIR severity was not related to whitefly density in our experiment, even if the threshold was largely reached. This very low prevalence of TIR (8%) could be due to different factors, such as tomato plants phenology at the time of the infestation or confined infestation on the plant. McKenzie and Albano (2009) noted that timing is a significant factor in the induction of TIR by whiteflies, and that only a low density of whitefly nymphs within two weeks of harvest can induce TIR. In our experiment, whiteflies were introduced on one leaf of 10-leaf plants (one leaf presenting between 5 and 7 leaflets), then removed after one or three weeks, while the harvest occurred later in the season. Therefore, it can be concluded that either the infestation period was not long enough for the SLW to have a strong impact on the tomato plants and induce TIR, or the tomato plants could have recovered from the whitefly populations before fruit maturation occurred. Nevertheless, our results suggest that a rapid intervention after noticing a SLW infestation could limit the impact of TIR.
Effect of predators on whitefly density
Using generalist predators represent an interesting strategy to rapidly control SLW populations. For example, in an experiment carried out in a greenhouse in Mexico, a rate of introduction of one Dicyphus bug per tomato plant and Ephestia eggs induced an almost 90% reduction in adult whitefly populations (Calvo et al. 2016). In another expe-riment, the introduction of a single D. hesperus on a tomato plant infested with 30 adult whiteflies significantly reduced whitefly populations after five weeks (seven times fewer larvae and pseudo-pupae in treatments with one Dicyphus than in treatments without predator) (Calvo, Torres et al. 2018). Another experiment demonstrated a 60% reduction of whitefly nymphs in five weeks with the introduction of 2, 4 or 6 pairs of D. hesperus during three consecutive weeks (Smith and Krey 2019). We observed similar results in our experiment, since D. hesperus had a significant impact on the SLW density in a relatively short time (five weeks), making it a good candidate for the control of the SLW in tomato crops. However, the decrease of whitefly populations was lower in our experiment (53%) than in Calvo, Torres et al. (2018) or Smith and Krey (2019). This difference could be attributed to the absence of supplementary food resources in our experiment. In fact, the addition of Ephestia eggs (Lepidoptera: Pyralidae), Artemia cysts (Crustacea: Branchiopoda, Artemiidae) or mullein plants to D. hesperus diet have been found to increase their population density (Labbé et al. 2018; Smith and Krey 2019).
In contrast, O. insidiosus had no discernible impact on the SLW than D. hesperus in our experiment, which means that this anthocorid bug does not play a significant role in the biological control of the SLW in tomato crops. Only a few studies focused on O. insidiosus as a predator of whiteflies (Coll and Ridgway 1995; Watve and Clower 1976), while many studies reported an effective use of O. insidiosus to control thrips and mite populations (Cloyd and Herrick 2017; Van de Veire and Degheele 1992; Xu et al. 2006). According to these previous studies and our results, it may be preferred to use O. insidiosus as a biological control agent to control other pests (especially thrips and mites) than whiteflies. It could be interesting to evaluate this predator in combination with D. hesperus against a multiple prey species.
CONCLUSION
Overall, our results show that the predatory bug, D. hesperus, could be used as an effective biological control agent to control infestations of the SLW. At low pest densities in particular, Dicyphus bugs may be more efficient to control whiteflies than parasitoids because they are more efficient at finding prey patches since these two biological control agents have different foraging behaviours (Bennett et al. 2009). Firstly, Dicyphus bugs use both olfactory and visual cues to detect whiteflies (Hazard 2008; Sinia et al. 2004), while parasitoids use mainly olfactory cues (Birkett et al. 2003), which makes them more effective at high pest densities. Secondly, mirid bugs, which are zoophytophagous, can initiate plant defences. Interestingly, some zoophytophagous mirid species feeding on tomato plants have been found to reduce the plants’ attractiveness to the SLW and activate the jasmonic acid signalling pathway, which attracts the parasitoid Encarsiaformosa Gahan, 1924 (Hymenoptera: Aphilinidae) (Pérez-Hedo et al. 2015). Although Dicyphus bugs are zoophytophagous and can feed on tomato plants, studies usually reveal no economic damage is incurred by this feeding behaviour (Calvo et al. 2016), especially if additional food resources are provided (Ephestia eggs or Artemia cysts). Finally, an optimal way of controlling the SLW would be to use a combination of biological control agents, such as the combination of entomo-pathogenic fungus and D. hesperus (Alma et al. 2007), D. hesperus and parasitoids (Calvo, Torres-Ruiz et al. 2018), or a combination of these three types of agents (Labbé et al. 2009). Although intraguild predation of mirid bugs on parasitoids has been reported (Bennett et al. 2009; McGregor and Gillespie 2005), the use of parasitoids in addition to D. hesperus resulted in better and more cost-effective control of the SLW (Calvo, Torres-Ruiz et al. 2018). However, further studies are needed to determine economic thresholds, the number of biological control agents to release in the greenhouse, and the best timing of intervention.
Appendices
Acknowledgements
Thanks to Maud Lemay, Mylène Vaillancourt, Stefano Campagnaro, Pascale Boulay, and Myriam Bonneville-Décarie for help in conducting the experiments. This study was funded through the Program Prime-vert component 4 2013-2018, provided by the Quebec Ministry of Agriculture, Fisheries and Food.
REFERENCES
- Alma, C.R., M.S. Goettel, B.D. Roitberg, and D.R. Gillespie. 2007. Combined effects of the entomopathogenic fungus, Paecilomyces fumosoroseus Apopka-97, and the generalist predator, Dicyphus hesperus, on whitefly populations. BioControl 52: 669-681.
- Bass, C., I. Denholm, M.S. Williamson, and R. Nauen. 2015. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 121: 78-87.
- Bennett, J.A., D.R. Gillespie, J.L. Shipp, and S.L. Vanlaerhoven. 2009. Foraging strategies and patch distributions: intraguild interactions between Dicyphus hesperus and Encarsia formosa. Ecol. Entomol. 34: 58-65.
- Birkett, M.A., K. Chamberlain, E. Guerrieri, J.A. Pickett, L.J. Wadhams, and T. Yasuda. 2003. Volatiles from whitefly-infested plants elicit a host-locating response in the parasitoid, Encarsia formosa. J. Chem. Ecol. 29: 1589-1600.
- Buitenhuis, R., L. Shipp, C. Scott-Dupree, A. Brommit, and W. Lee. 2014. Host plant effects on the behaviour and performance of Amblyseius swirskii (Acari: Phytoseiidae). Exp. Appl. Acarol. 62: 171-180.
- Cahill, M., W. Jarvis, K. Gorman, and I. Denholm. 1996. Resolution of baseline responses and documentation of resistance to buprofezin in Bemisia tabaci (Homoptera: Aleyrodidae). Bull. Entomol. Res. 86, 117–122.
- Calvo, F.J., A. Torres, E.J. González, and M.B. Velázquez. 2018. The potential of Dicyphus hesperus as a biological control agent of potato psyllid and sweetpotato whitefly in tomato. Bull. Entomol. Res. 108: 765-772.
- Calvo, F.J., A. Torres-Ruiz, J. Velázquez-González, E. Rodríguez-Leyva, and J.R. Lomeli-Flores. 2016. Evaluation of Dicyphus hesperus for biological control of sweet potato whitefly and potato psyllid on greenhouse tomato. BioControl 61: 415-424.
- Calvo, F.J., A. Torres-Ruiz, J. Velázquez-González, E. Rodríguez-Leyva, and J.R. Lomeli-Flores. 2018. Improved sweetpotato whitefly and potato psyllid control in tomato by combining the mirid Dicyphus hesperus (Heteroptera: Miridae) with specialist parasitic wasps. J. Econ. Entomol. 111: 549-555.
- Castañé, C., J. van der Blom, and P.C. Nicot. 2020. Tomatoes. Pages 487-511 in M.L. Gullino, R. Albajes, and P.C. Nicot (eds.), Integrated pest and disease management in greenhouse crops. Springer, Cham, Switzerland.
- Cloyd, R.A., and N.J. Herrick. 2017. Ecology and role of the rove beetle, Dalotia coriaria, and insidious flower bug, Orius insidiosus, in greenhouse biological control programs. Adv. Entomol. 5: 115-126.
- Coll, M., and R.L. Ridgway. 1995. Functional and numerical responses of Orius insidiosus (Heteroptera: Anthocoridae) to its prey in different vegetable crops. Ann. Entomol. Soc. Am. 88: 732-738.
- Cook, M.J.W. 1986. Bemisia tabaci: a literature survey on the cotton whitefly with an annotated bibliography. CAB International Institute of Biological Control, Ascot, United Kingdom. 121 pp.
- De Barro, P.J., S.-S. Liu, L.M. Boykin, and A.B. Dinsdale. 2011. Bemisia tabaci: a statement of species status. Annu. Rev. Entomol. 56: 1-19.
- Gerling, D., Ò. Alomar, and J. Arnò. 2001. Biological control of Bemisia tabaci using predators and parasitoids. Crop Prot. 20: 779-799. doi:10.1016/S0261-2194(01)00111-9
- Greenberg, S.M., W.A. Jones, and T.-X. Liu. 2002. Interactions among two species of Eretmocerus (Hymenoptera: Aphelinidae), two species of whiteflies (Homoptera: Aleyrodidae), and tomato. Environ. Entomol. 31: 397-402.
- Guershon, M., and D. Gerling. 2006. Effects of plant and prey characteristics on the predatory behavior of Delphastus catalinae. Entomol. Exp. Appl. 121: 15-21.
- Hamilton, A.J., M. Bouzayen, and D. Grierson. 1991. Identification of a tomato gene for the ethylene-forming enzyme by expression in yeast. Proc. Natl. Acad. Sci. U.S.A. 88: 7434-7437.
- Hazard, T.J.R. 2008. Close-range prey location, recognition and foraging decisions by the generalist predator, Dicyphus hesperus knight (heteroptera: miridae). Simon Fraser University, Department of Biological Sciences. Available online [https://summit.sfu.ca/item/9102].
- Hoddle, M.S., and R. Van Driesche. 1999. Evaluation of Eretmocerus eremicus and Encarsia formosa (Hymenoptera: Aphelinidae) Beltsville strain in commercial greenhouses for biological control of Bemisia argentifolii (Homoptera: Aleyrodidae) on colored poinsettia plants. Fla. Entomol. 82: 556-569.
- Horowitz, A.R., Z. Mendelson, M. Cahill, I. Denholm, and I. Ishaaya. 1999. Managing resistance to the insect growth regulator, pyriproxyfen, in Bemisia tabaci. Pestic. Sci. 55: 272-276. doi:10.1002/(SICI)1096-9063(199903)55:3<272::AID-PS908>3.0.CO;2-Y
- Jones, D.R. 2003. Plant viruses transmitted by whiteflies. Eur. J. Plant Pathol. 109: 195-219.
- Labbé, R.M., D. Gagnier, A. Kostic, and L. Shipp. 2018. The function of supplemental foods for improved crop establishment of generalist predators Orius insidiosus and Dicyphus hesperus. Sci. Rep. 8: 17790.
- Labbé, R.M., D.R. Gillespie, C. Cloutier, and J. Brodeur. 2009. Compatibility of an entomopathogenic fungus with a predator and a parasitoid in the biological control of greenhouse whitefly. Biocontrol Sci. Technol. 19: 429-446.
- McAuslane, H.J., and H.A. Smith. 2018. Sweetpotato whitefly B biotype, Bemisia tabaci (Gennadius) (Insecta: Hemiptera: Aleyrodidae). IFAS Extension University of Florida. Gainesville, Florida, USA. Available online [https://edis.ifas.ufl.edu/publication/IN286].
- McCollum, T.G., P.J. Stoffella, C.A. Powell, D.J. Cantliffe, and S. Hanif-Khan. 2004. Effects of silverleaf whitefly feeding on tomato fruit ripening. Postharvest Biol. Technol. 31: 183-190.
- McGregor, R.R., and D.R. Gillespie. 2005. Intraguild predation by the generalist predator Dicyphus hesperus on the parasitoid Encarsia formosa. Biocontrol Sci. Technol. 15: 219-227.
- McGregor, R.R., D.R. Gillespie, D.M.J. Quiring, and M.R.J. Foisy. 1999. Potential use of Dicyphus hesperus knight (Heteroptera: Miridae) for biological control of pests of greenhouse tomatoes. Biol. Control 16: 104-110.
- McKenzie, C.L., and J.P. Albano. 2009. The effect of time of sweetpotato whitefly infestation on plant nutrition and development of tomato irregular ripening disorder. HortTechnology 19: 353-359.
- Nauen, R., and I. Denholm. 2005. Resistance of insect pests to neonicotinoid insecticides: current status and future prospects. Arch. Insect Biochem. Physiol. 58: 200-215.
- Oeller, P.W., M.-W. Lu, L.P. Taylor, D.A. Pike, and A.A. Theologis. 1991. Reversible inhibition of tomato fruit senescence by antisense RNA. Science 254: 437439.
- Oliveira, M.R.V., T.J. Henneberry, and P. Anderson. 2001. History, current status, and collaborative research projects for Bemisia tabaci. Crop Prot. 20: 709-723. doi:10.1016/S0261-2194(01)00108-9
- Palumbo, J.C., A.R. Horowitz, and N. Prabhaker. 2001. Insecticidal control and resistance management for Bemisia tabaci. Crop Prot. 20: 739-765. doi:10.1016/S0261-2194(01)00117-X
- Pérez-Hedo, M., S. Bouagga, J.A. Jaques, V. Flors, and A. Urbaneja. 2015. Tomato plant responses to feeding behavior of three zoophytophagous predators (Hemiptera: Miridae). Biol. Control 86: 46-51.
- Prabhaker, N., D.L. Coudriet, and D.E. Meyerdirk. 1985. Insecticide resistance in the sweetpotato whitefly, Bemisia tabaci (Homoptera: Aleyrodidae). J. Econ. Entomol. 78: 748-752.
- R Core Team. 2017. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available online [https://www.R-project.org].
- Sanchez, J.A., D.R. Gillespie, and R.R. McGregor. 2003. The effects of mullein plants (Verbascum thapsus) on the population dynamics of Dicyphus hesperus (Heteroptera: Miridae) in tomato greenhouses. Biol. Control 28: 313-319. doi:10.1016/S1049-9644(03)00116-6
- Schuster, D.J. 2002. Action threshold for applying insect growth regulators to tomato for management of irregular ripening caused by Bemisia argentifolii (Homoptera: Aleyrodidae). J. Econ. Entomol. 95: 372-376.
- Schuster, D.J., T.F. Mueller, J.B. Kring, and J.F. Price. 1990. Relationship of the sweetpotato whitefly to a new tomato fruit disorder in Florida. HortScience 25: 1618-1620.
- Schuster, D.J., H.A. Smith, and P.A. Stansly. 2019. A threshold for timing applications of insecticides to manage the silverleaf whitefly and irregular ripening on tomato. IFAS Extension University of Florida. Gainesville, Florida, USA. Available online [https://edis.ifas.ufl.edu/publication/IN499].
- Sinia, A., B. Roitberg, R.R. McGregor, and D.R. Gillespie. 2004. Prey feeding increases water stress in the omnivorous predator Dicyphus hesperus. Entomol. Exp. Appl. 110: 243-248.
- Smith, H.A., and K.L. Krey. 2019. Three release rates of Dicyphushesperus (Hemiptera: Miridae) for management of Bemisia tabaci (Hemiptera: Aleyrodidae) on greenhouse tomato. Insects 10: 213.
- Van de Veire, M., and D. Degheele. 1992. Biological control of the western flower thrips, Frankliniella occidentalis (Pergande)(Thysanoptera: Thripidae), in glasshouse sweet peppers with Orius spp.(Hemiptera: Anthocoridae). A comparative study between O. niger (Wolff) and O. insidiosus (Say). Biocontrol Sci. Technol. 2: 281-283.
- van Lenteren, J.C. 2012. The state of commercial augmentative biological control: plenty of natural enemies, but a frustrating lack of uptake. BioControl 57: 1-20.
- Watve, C.M., and D.F. Clower. 1976. Natural enemies of the bandedwing whitefly in Louisiana. Environ. Entomol. 5: 1075-1078.
- Wong, S.K., and S.D. Frank. 2013. Pollen increases fitness and abundance of Orius insidiosus Say (Heteroptera: Anthocoridae) on banker plants. Biol. Control 64: 45-50.
- Xu, X., C. Borgemeister, and H.-M. Poehling. 2006. Interactions in the biological control of western flower thrips Frankliniella occidentalis (Pergande) and two-spotted spider mite Tetranychus urticae Koch by the predatory bug Orius insidiosus Say on beans. Biol. Control 36: 57-64.
List of figures
Figure 1
Trial average number of Bemisia tabaci eggs (± SE) (a) or nymphs and pupae (b) per leaf relative to initial whitefly introduction rate and duration of infestation
The number of eggs, larvae or pseudo-nymphs increased with the initial density of B. tabaci introduced. Different letters indicate statistical differences among treatments.
Figure 2
Average rate of tomato irregular ripening (TIR) disorder (± SE) observed in function of the Bemisia tabaci introduction rate and duration of infestation
The introduction rate and the duration of infestation had no significant effect on the rate of TIR.
Figure 3
Trial average Bemisia tabaci density (± SE) in function of the density of Dicyphus hesperus
Statistical significance is indicated by different letters (α = 0.05).
Figure 4
Bemisia tabaci density (± SE) in function of the density of Orius insidiosus
The O. insidiosus introduction rate had no significant effect on B. tabaci density.