Abstracts
Abstract
The pathogenicity of Gremmeniella abietina var. balsamea isolated from balsam fir (Abies balsamea) was tested on different conifer hosts, including A. balsamea. Pathogenicity of the fungus was positive on balsam fir only. This pathogen could not infect other conifers, not even spruce species which are reported as hosts for the taxon G. abietina var. balsamea. Also, isolates from spruces and pines were pathogenic only on their respective hosts. These results raise questions on the taxonomic status of the two pathogens classified as G. abietina var. balsamea.
Keywords:
- Scleroderris canker,
- Picea mariana,
- Pinus banksiana
Résumé
Gremmeniella abietina var. balsamea isolé du sapin baumier (Abies balsamea) a été soumis à une épreuve de pouvoir pathogène chez différentes espèces de conifères, y compris A. balsamea. Le pouvoir pathogène de ce champignon était positif seulement sur le sapin baumier. Ce champignon pathogène n’a pu infecter d’autres conifères, pas même les épinettes qui sont pourtant des hôtes reconnus du taxon G. abietina var. balsamea. De plus, les isolats provenant des épinettes et des pins étaient pathogènes seulement sur leurs hôtes respectifs. Ces résultats soulèvent des questions quant au statut taxonomique des deux champignons pathogènes classés comme G. abietina var. balsamea.
Mots clés:
- Chancre scléroderrien,
- Picea mariana,
- Pinus banksiana
Article body
Gremmeniella abietina (Lagerb.) Morelet is frequently found on Pinus spp. (Morelet 1969), and it also occurs on balsam fir (Abies balsamea (L.) Mill.) in the boreal forest of North America. On balsam fir, this fungus was recognized as the taxon Gremmeniella abietina (Lagerb.) Morelet var. balsamea Petrini, Petrini, Laflamme & Ouellette (Petrini et al. 1989). This variety also comprises Gremmeniella entities occurring on spruces, namely white spruce (Picea glauca (Moench) Voss) and black spruce (P. mariana (Mill.) B.S.P.). Smerlis (1967) tested the pathogenicity of different isolates of Gremmeniella on several native conifer species in Quebec. The results from his trial show that the isolates from black spruce caused symptoms on Picea spp., but not on A. balsamea, Larix spp., or Pinus spp. (Smerlis 1967). Another pathogenicity test conducted by Smerlis (1968) gave erratic results and the author suggested that the use of recently transplanted or suppressed trees may have caused such results. Later, and under field conditions using well-established trees, Laflamme et al. (1996) tested the host preference of six isolates, two per host, from naturally regenerated black spruce, balsam fir and jack pine (Pinus banksiana Lamb.). Our results clearly showed that each isolate thrives on the host from which it was isolated.
In this note, we present results on fungal isolation from diseased balsam fir leaders as well as the results of pathogenicity tests conducted using isolates from naturally occurring cankers caused by G. abietina var. balsamea on balsam fir and black spruce and by G. abietina var. abietina on jack pine and red pine (North American race). Cross inoculations were performed in 1983 on several conifer hosts to prove through Koch’s postulate the pathogenicity of these Gremmeniella isolates.
Isolations from diseased balsam fir stem leaders were made in 1983 from samples collected in the Réserve faunique des Laurentides (47°35’N; 71°13’W), Quebec, Canada. Three small pieces of necrotic bark were cut with a sterile scalpel, surface sterilized for 5 s in 1% HgC12 solution, washed in sterile distilled water and placed in tubes containing 3% malt agar. The cultures were incubated at 15°C. A total of 100 leaders were sampled.
Twenty-one different fungi were isolated from the naturally infected balsam fir leaders. Gremmeniella abietina var. balsamea was obtained from 89% of the samples. Sydowia polyspora (Brev. & v. Tav.) E. Müll. ranked second with a frequency of 31%. Lophium mytilinum (Pers. ex Fr.) Fr. and Epicoccum purpurascens Ehrenb. ex Schlecht. were isolated from 21 and 16% of the leaders, respectively. The other mostly unidentified fungi ranged in frequencies from 1 to 6%.
Inoculations on eastern white cedar (Thuja occidentalis L.), creeping juniper (Juniperus horizontalis Moench) and ground juniper (J. communis L. var. depressa Pursh) were done in 1983 in Saint-André-de-Kamouraska (47°41’N; 69°44’W), and those on eastern hemlock (Tsuga canadensis (L.) Carr.) were done in Saint-Louis-de-Blandford, Arthabasca (46°15’N; 72°00’W). The other tree species (Table 1) were treated at the Valcartier Forest Station (46°54’N; 71°29’W) of the Canadian Forest Service, near Quebec City. The inoculations were carried out with i) mycelium from monoascospore isolates, ii) mycelium from monoconidia isolates, iii) ascospores, and iv) conidia.
All inoculations with mycelium were done in the fall of 1983 on current-year stem or branch leaders by removing needles from the base of shoots and cutting a tongue-like section in the bark, a method previously described by Smerlis (1969). Six mono-ascospore isolates of G. abietina var. balsamea from balsam fir were used as inocula; four cultures originated from the Réserve faunique des Laurentides and the two others were prepared from samples collected at Lac Albanel (50°54’N; 73°18’W) in the James Bay region, and in Saint-Jean-de-Cherbourg (48°51’N; 67°07’W) in the Matane region. Two monoconidial isolates of G. abietina var. balsamea from balsam fir were used; they originated from the Réserve faunique des Laurentides. Inoculations with G. abietina var. abietina from jack pine were carried out with one monoascospore and one monoconidial isolate of the North American strain prepared from specimens collected in Saint-Urbain (47°33’N; 70°32’W) in the Charlevoix region. Finally, inoculations with G. abietina var. balsamea from black spruce were carried out in 1983 with one monoascospore and one monoconidial isolate prepared from specimens collected in the Réserve faunique des Laurentides. All cultures were grown on 3% malt agar at 15°C. The inoculations ranged in number per tree species or source of inoculum from 5 to 23, but the original data are not available anymore. Isolations from positive inoculations were made on 3% malt agar and the tubes were maintained at 15°C.
Inoculations with ascospores and conidia were carried out in Valcartier in mid-July of 1983 in duplicate on 50 cm x 50 cm plots on which nine seedlings had been planted. The tree species treated are listed in Table 1. The method of inoculation with ascospores discharged from apothecia has already been described (Smerlis 1976). In short, branches bearing mature apothecia were attached to a wire over the seedlings. Branches of red pine (P. resinosa Ait.) bearing apothecia of G. abietina var. abietina (North American strain) were collected in Sainte-Hedwidge (48°35’N; 72°21’W) in the Lac-Saint-Jean region, and those of black spruce and balsam fir (G. abietina var. balsamea) were collected in the Réserve faunique des Laurentides.
Conidial suspensions, prepared at the rate of four pycnidia per 100 mL of distilled water, were made from fructifications of samples collected on the three tree species at the previously mentioned localities. Each treated plot was sprayed with 25 mL of spore suspension with a hand-operated sprayer. The number of conidia per mL is not available anymore. Distilled water was applied on the control plots.
Table 1
Results of inoculation with Gremmeniella abietina var. balsamea from balsam fir and spruce and G. abietina var. abietina from jack pine and red pine (North American race) on several conifer species with four types of inoculum: 1 = mycelium from monoascospore isolates, 2 = mycelium from monoconidial isolates, 3 = ascospores discharged from apothecia, and 4 = conidial suspensions; + = symptoms; - = no symptoms
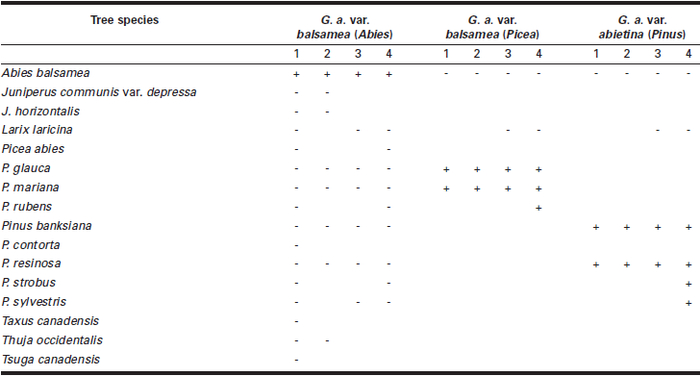
Inoculations with pure cultures of G. abietina var. balsamea from balsam fir on that host resulted in symptoms ranging in frequencies from 67 to 100%. Cankers up to 8 cm long, generally causing mortality of the inoculated leaders, were formed. The fungus was reisolated from all sets of inoculations, although not from all individual infections tested. Pycnidia of G. abietina var. balsamea with conidia developed during the first summer following the inoculations on some of the leaders treated with cultures of either form of the spores. Apothecia were observed on dead leaders the second summer after the inoculations. Treatments with ascospores or conidial suspensions of G. abietina var. balsamea from balsam fir similarly resulted in symptoms only on balsam fir (Table 1). All inoculated balsam fir seedlings were infected. Applications of conidial suspensions caused symptoms manifested by defoliation of most or all branch leaders and also of the stem terminal. The defoliation was generally associated with subsequent mortality of the seedlings.
The inoculations with mycelium of G. abietina var. balsamea from balsam fir on the other tree species were all negative (Table 1). Controls where distilled water was applied, likewise, showed no signs or symptoms.
Inoculations with G. abietina var. abietina (North American strain) and G. abietina var. balsamea from spruce by any of the three methods caused symptoms on pines and spruces, respectively, and they were not pathogenic on balsam fir (Table 1). Controls showed no signs or symptoms.
These pathogenicity tests show that G. abietina var. balsamea from balsam fir is pathogenic only on balsam fir. It also shows that G. abietina var. balsamea from spruce is pathogenic only on spruce as was demonstrated previously on four spruce species: P. glauca, P. mariana, P. rubens Sarg., and P. abies (L.) Karst. (Smerlis 1967). Finally, G. abietina var. abietina caused diseases only on pine species.
In the reappraisal of the genus Gremmeniella, Petrini et al. (1989) decided not to create new species of Gremmeniella found on spruce and balsam fir in eastern Canada because of the lack of well-defined morphological differences between specimens from pines, spruces and balsam fir. Dimensions of asci, ascospores and conidia showed overlaps from specimens collected on these tree species. However, electrophoresis showed four different patterns among the reference cultures, two on pines (North American race and European race), one on Larix spp., and the other one represented by isolates from spruces and balsam fir in eastern Canada. On that basis, Petrini et al. (1989) created two varieties inside the type species, the variety abietina, mainly found on pines, and the variety balsamea, affecting spruce and balsam fir in the province of Quebec (Canada). Also, a phylogeny study of Gremmeniella spp. based on sequences of the 5.8S rDNA and internal transcribed spacer (ITS) region shows a level of divergence of ITS between the two varieties abietina and balsamea as large as the level of divergence between the species G. laricina and G. abietina var. abietina or between G. laricina and G. abietina var. balsamea (Hamelin and Rail 1997). In the same study, the host-related genetic differentiation in G. abietina var. balsamea is supported by sequences of the ITS region.
In view of the host specificity of Gremmeniella found on spruces and balsam fir in eastern Canada, as well as the differences shown by phylogenic studies, we strongly suggest reviewing the taxonomic status of the two fungal pathogens causing diseases on spruce and balsam fir, which are currently included in the taxon G. abietina var. balsamea.
Appendices
Acknowledgements
The assistance provided by the late Marcel Saint-Laurent, technician at the Laurentian Forestry Centre, is acknowledged. We thank Pierre DesRochers and Danny Rioux for their critical reading of the manuscript, and Louis Parrot, former professor at Université Laval, for his assistance with the search of Smerlis’s documents.
References
- Hamelin, R.C., and J. Rail. 1997. Phylogeny of Gremmeniella spp. based on sequences of the 5.8S rDNA and internal transcribed spacer region. Can. J. Bot. 75 : 693-698.
- Laflamme, G., A. Ylimartimo, and R. Blais. 1996. Host preference of two Gremmeniella abietina varieties on balsam fir, jack pine and black spruce in eastern Canada. Can. J. Plant Pathol. 18 : 330-334.
- Morelet, M. 1969. Un discomycète inoperculé nouveau. Bull. Soc. Sci. Nat. Archéol. 183 : 9.
- Petrini, O., L.E. Petrini, G. Laflamme, and G.B. Ouellette. 1989. Taxonomic position of Gremmeniella abietina and related species: a reappraisal. Can. J. Bot. 67 : 2805-2814.
- Smerlis, E. 1967. Occurrence and pathogenicity of Scleroderris lagerbergii in Quebec. Plant Dis. Rep. 51 : 584-585.
- Smerlis, E. 1968. Additional information on the pathogenicity of Scleroderris lagerbergii. Plant Dis. Rep. 52 : 738-739.
- Smerlis, E. 1969. Synonymy and pathogenicity of Dermea piceina. Can. J. Bot. 47 : 213-214.
- Smerlis, E. 1976. Evaluation of fungicides for control of Gremmeniella abietina. I. Laboratory and preliminary field assays. Can. For. Serv., Laurentian For. Res. Cent., Info. Rep. LAU-X-23.
List of tables
Table 1
Results of inoculation with Gremmeniella abietina var. balsamea from balsam fir and spruce and G. abietina var. abietina from jack pine and red pine (North American race) on several conifer species with four types of inoculum: 1 = mycelium from monoascospore isolates, 2 = mycelium from monoconidial isolates, 3 = ascospores discharged from apothecia, and 4 = conidial suspensions; + = symptoms; - = no symptoms