Article body
L’infection par le virus Chikungunya est une arbovirose évoluant sur un mode épidémique, dans les continents africain et asiatique, mais aussi dans les îles de l’Océan Indien, comme le souligne l’épidémie de grande ampleur actuellement en cours à l’île de la Réunion, après avoir touché les Comores, Mayotte et l’île Maurice, depuis le début de l’année 2005 [1].
Le virus et son vecteur
Le virus Chikungunya est un arbovirus de la famille des Togaviridae, du genre Alphavirus, appartenant au complexe antigénique Semliki Forest [2]. La première description, qui remonte à 1952 en Tanzanie, lui a valu son nom qui signifie, en swahili, « qui marche courbé en avant » [3]. Il est responsable d’un tableau clinique articulaire similaire à celui observé dans d’autres alphaviroses, comme celles dues aux virus Ross River (VRR), Barmah Forest, O’Nyong-Nyong, Sindbis et Mayaro [4]. L’analyse phylogénétique des souches de Chikungunya permet d’identifier des clusters différents pour les souches d’Afrique de l’Est, de l’Ouest ou d’Asie, et de souligner la proximité importante avec le virus O’Nyong-Nyong [2]. Il s’agit d’un virus enveloppé à ARN simple brin, qui contient 3 protéines de structure, les glycoprotéines de surface E1 à activité hémagglutinante, et E2, et une protéine de capside C non glycosylée [5]. Il est transmis par des moustiques du genre Aedes, d’espèces différentes selon les zones considérées [6]. Les cycles de transmission diffèrent selon le continent : en Afrique, la transmission se maintient par un cycle sylvatique, rural, impliquant principalement Aedes furcifer et Aedes africanus, vecteurs par ailleurs de la fièvre jaune en cycle sylvatique, alors qu’en Asie, le cycle est plutôt urbain, impliquant Aedes aegypti et Aedes albopictus (Figure 1), moustiques anthropophiles et péridomestiques, impliqués aussi dans le cycle de transmission de la dengue [2, 7]. En Afrique, le réservoir du virus est animal, constitué par les primates, certains rongeurs et oiseaux, qui entretiennent le cycle sylvatique [4]. La présence de virus a pu être retrouvée chez le bétail, sans que son rôle dans la transmission n’apparaisse important. Aucun réservoir animal n’a été formellement documenté en Asie [2]. La large répartition géographique des vecteurs rend possible l’émergence du virus dans de nombreuses régions [6, 8], comme cela a pu être constaté pour d’autres arbovirus, comme le virus West Nile, introduit puis installé sur le continent nord-américain depuis 1999. La transmission trans-ovarienne du virus n’a été mise en évidence que dans des conditions expérimentales [9], et il n’est pas certain qu’elle ait lieu dans des conditions naturelles.
Figure 1
Aedes albopictus, le moustique en cause dans l’épidémie réunionnaise.
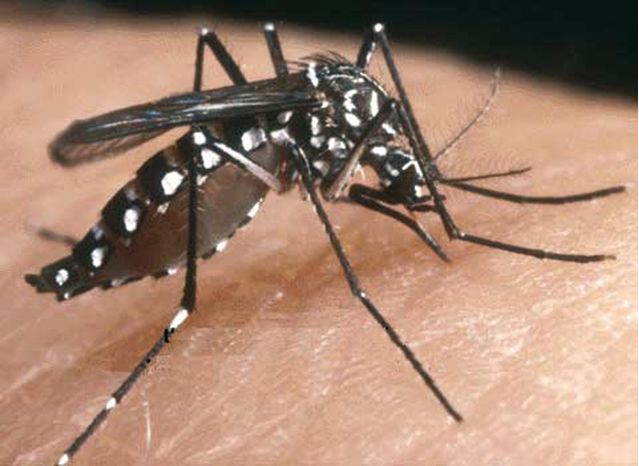
Au plan épidémiologique, depuis la description initiale en Tanzanie, ce virus a été régulièrement à l’origine de petites poussées épidémiques cycliques en milieu rural, principalement en Afrique australe et de l’Est, de l’Ouganda à l’Afrique du Sud, en Afrique Centrale, la dernière épidémie du Congo ayant eu la particularité d’être urbaine, comme les épidémies asiatiques [10], plus rarement en Afrique de l’Ouest, en particulier au Sénégal [11]. Il est considéré comme endémique en milieu rural en Afrique, responsable de nombreux cas probablement non diagnostiqués, et associé à une séroprévalence pouvant être supérieure à 50 % [11]. Parallèlement, des poussées épidémiques ont été observées en Inde, au Sri Lanka, en Asie du Sud-Est (Thaïlande, Myanmar, Vietnam, Laos, Cambodge, Indonésie, plus récemment Malaisie), aux Philippines [12, 13]. La fréquence plus importante des épidémies en Asie peut être reliée au caractère anthropophile des moustiques vecteurs en cause [2]. Jusqu’à l’épidémie actuelle, sa présence dans les îles de l’Océan Indien n’avait été révélée que par des enquêtes de séroprévalence, sans notification d’épidémies [6]. Des descriptions cliniques ont aussi été faites chez des voyageurs, la dernière au retour de Côte d’Ivoire [8]. Dans la majorité des cas, les épidémies surviennent après un pic de précipitations, favorisant la pullulation des vecteurs.
Les conséquences de l’infection chez l’homme
Chez l’homme, l’infection se manifeste le plus souvent sous forme de cas sporadiques ou de petites épidémies. Elle atteint de façon relativement homogène enfants et adultes, hommes et femmes. Elle peut être asymptomatique, mais, classiquement, le tableau clinique est celui d’une polyarthrite aiguë fébrile de début brutal, caractéristique de nombreuses alphaviroses [4], survenant après un délai d’incubation de 2 à 10 jours [8]. Cette atteinte articulaire concerne principalement les poignets et les chevilles, mais aussi les genoux, les articulations des mains et des pieds, beaucoup plus rarement les hanches ou les épaules. Elle se limite le plus souvent à des arthralgies, mais d’authentiques arthrites inflammatoires et ténosynovites peuvent être observées, responsables d’une impotence fonctionnelle majeure [13, 14]. À cette atteinte articulaire s’associent fréquemment des céphalées, des myalgies importantes (dans 70 % à 99 % des cas), une éruption cutanée maculo-papuleuse du tronc et des membres (dans environ 50 % des cas), parfois prurigineuse, qui apparaît volontiers après quelques jours, voire une polyadénopathie cervicale ou une conjonctivite [1, 4, 13]. Des formes hémorragiques mineures, avec gingivorragie, épistaxis ou purpura ont été décrites principalement en Asie, mais leur fréquence peut être élevée, évaluée à 23 % sur les 7 148 premiers cas recensés de l’épidémie actuelle de la Réunion [1]. Jusqu’à présent, les formes compliquées n’étaient qu’exceptionnellement décrites. Cependant, l’épidémie actuelle de la Réunion a permis de mettre en évidence l’existence de formes neurologiques graves jamais antérieurement décrites, mais non surprenantes au regard de la parenté avec d’autres alphavirus neurotropes proches (dont le virus de l’encéphalite équine) sévissant sur le continent américain : ces méningo-encéphalites et polyradiculonévrites ont ainsi été décrites principalement chez des personnes âgées ou présentant une pathologie sous-jacente, et chez des nouveau-nés, dont les mères avaient été infectées peu de temps avant l’accouchement, probablement en rapport avec une transmission materno-foetale tardive [1]. L’évolution se fait habituellement vers une amélioration rapide, avec disparition de la fièvre en 1 à 10 jours, des signes cutanés en 2 à 3 jours [4, 12], des signes articulaires en quelques semaines. Classiquement, cette maladie n’est pas connue pour être responsable de mortalité directe, mais une enquête est actuellement en cours à la Réunion, pour déterminer l’imputabilité directe ou indirecte du virus dans plusieurs dizaines de décès survenus dans un contexte d’infection aiguë [15]. La particularité évolutive, commune à plusieurs alphaviroses (surtout le VRR), est de voir perdurer l’atteinte articulaire sur un mode subaigu ou chronique, durant plusieurs mois, voire même plusieurs années : si cette évolution est fréquente pour le VRR, de l’ordre de 50 % des patients à un an et de 10 % à 50 % à 2 ans [4], elle ne représenterait dans une étude rétrospective sud-africaine « que » 10 % des patients 3 à 5 ans après une infection aiguë par le virus Chikungunya [14]. Elle se caractérise par la persistance durable d’arthralgies périphériques de type inflammatoire, plus rarement d’arthrites non destructrices, avec raideur articulaire et nécessité de dérouillage matinal, soit permanentes, soit d’évolution intermittente par poussées, et s’accompagnant volontiers de la persistance d’un syndrome inflammatoire modéré [14]. Elle s’observe d’autant plus fréquemment que l’âge est élevé. Le mécanisme en cause, étudié pour le VRR, pourrait témoigner de la persistance du virus dans le tissu synovial, le périoste, les tendons et ligaments, ainsi que de sa capacité de réplication intramonocytaire et d’induction de la sécrétion de médiateurs de l’inflammation, tels que l’interféron de type I ou le NO [4, 16]. Un modèle murin d’inflammation induite par le VRR a été développé récemment, permettant de suggérer le rôle majeur de l’immunité innée dans le développement de l’atteinte articulaire [16].
En phase aiguë, le diagnostic est moléculaire ou sérologique par recherche d’IgM par une méthode spécifique immuno-enzymatique (ELISA) : les IgM apparaissent en moyenne 3 à 5 jours après le début du tableau clinique [4]. Le diagnostic moléculaire, effectué en France dans les Centres Nationaux de Référence (CNR des Arbovirus, Institut Pasteur, Lyon ; Unité de Virologie, Section Diagnostic des Arbovirus, Institut de Médecine Tropicale du Service de Santé des Armées, Marseille), repose sur la mise en évidence de l’ARN viral par RT-PCR simple (voire nichée ou quantitative), réalisable sur sang ou autre type de prélèvement biologique [17]. L’isolement du virus est aussi possible par culture sur cellules. Pour les cas sporadiques, le diagnostic est en fait le plus souvent rétrospectif par sérologie, par ELISA ou des techniques plus anciennes (inhibition d’hémagglutination, fixation du complément, séroneutralisation sur plaques). Les IgG apparaissent entre le 8e et le 30e jour. La grande proximité phylogénétique avec le virus O’Nyong-Nyong et d’autres alphavirus rend possible les réactions croisées [4].
La prise en charge médicale est purement symptomatique, reposant sur des traitements antalgiques et anti-inflammatoires, plutôt après la phase aiguë pour ces derniers. Ces traitements n’ont cependant aucun effet préventif sur la survenue d’une évolution chronique [8]. Une corticothérapie peut s’avérer nécessaire dans les formes sévères d’évolution subaiguë-chronique. L’interféron α et la ribavirine auraient une action inhibitrice in vitro, avec effet synergique de l’association [18].
La prévention de cette infection est à la fois collective et individuelle, reposant sur la lutte anti-vectorielle. Elle nécessite, pour les individus, une prophylaxie d’exposition (vêtements longs, répulsifs cutanés, insecticides pyréthrinoïdes sur les vêtements, moustiquaire) et, collectivement, une lutte anti-vectorielle à large échelle, avec épandages précautionneux d’insecticides, adulticides et/ou larvicides, et élimination des gîtes larvaires potentiels, particulièrement en zone péri-domestique (pots de fleur, récipients divers, pneus usagés, déchets encombrants…) [1]. Un vaccin vivant atténué contre le virus Chikungunya a été évalué par l’armée américaine : les résultats préliminaires retrouvaient une bonne immunogénicité et une tolérance qui semblait acceptable [19]. Le caractère préliminaire de ces résultats empêche cependant la mise à disposition rapide d’une vaccination à grande échelle.
L’épidémie actuelle dans l’Océan Indien
Elle a débuté en janvier 2005 aux Comores, puis s’est étendue principalement aux îles Maurice et de la Réunion en mars 2005, avant de sembler s’éteindre au cours de l’hiver austral. Depuis le début de l’année 2006, avec la reprise des précipitations dans cette région, on observe une flambée épidémique de grande ampleur, touchant principalement la Réunion (à l’exception du centre), mais aussi les autres îles de l’Océan Indien (Seychelles, Mayotte, Maurice,… et probablement Madagascar) (Tableau I) [15, 20]. À la Réunion, durant la semaine du 20 au 26 février 2006, une estimation de 19 200 nouveaux cas a été rapportée. Parmi ces derniers, plus de 750 personnes ont consulté dans un service d’urgence hospitalier, 35 % à 40 % des cas ayant été hospitalisés [15].
Tableau I
Épidémie de Chikungunya dans l’Océan Indien en 2005-2006, au 28 février 2006.
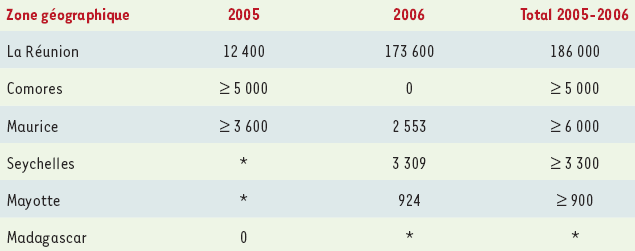
Les chiffres indiqués correspondent au nombre de cas suspects ou estimés (pour la Réunion). Ils sont tirés des sites Internet de l’Institut de Veille Sanitaire et de Promed, au 7 mars 2006.
*Données non disponibles (cas décrits, non quantifiés précisément).
Conclusions
L’infection par le virus Chikungunya, actuellement responsable d’une poussée épidémique de grande ampleur dans les îles de l’Océan Indien, associée à une morbidité significative, souligne son caractère régulièrement ré-émergent, ou émergent dans de « nouveaux » territoires déjà colonisés par les moustiques vecteurs, parmi lesquels Aedes albopictus. L’extension de son aire de répartition rend potentiellement « réceptifs » de nouveaux territoires, tout particulièrement le continent américain, justifiant une surveillance épidémiologique et entomologique pour dépister de telles nouvelles émergences.
Les caractéristiques de l’épidémie actuelle et l’importance de la morbidité associée amènent à se poser un certain nombre de questions d’ordre épidémiologique et physiopathologique. Son profil évolutif semble en effet différent de ce qui a été observé jusqu’alors en l’absence de cycle connu de transmission non humaine, alors que les conditions écologiques et hygrométriques locales sont peu différentes des autres régions où ce virus a été précédemment isolé. L’absence d’immunité préalable dans la population explique-t-elle à elle seule l’importance de cette morbidité ? La souche virale en cause a-t-elle acquis des caractères de virulence particuliers, lui permettant d’assurer une virémie plus importante et/ou plus prolongée, ou lui conférant un tropisme particulier, lui permettant notamment de traverser la barrière hémato-encéphalique et la barrière foeto-placentaire ? Une meilleure adaptation du virus à son hôte arthropode est-elle à l’origine d’une meilleure capacité vectorielle ? Les études qui vont résulter de l’épidémie actuelle devront tenter de répondre à ces questions.
Appendices
Références
- 1. Infection par le virus Chikungunya à l’Ile de la Réunion. BEH 2006 (hors série) : 1-4.
- 2. Powers AM, Brault AC, Tesh RB, et al. Re-emergence of Chikungunya and O’Nyong-Nyong viruses : evidence for distinct geographical lineages and distant evolutionary relationships. J Gen Virol 2000 ; 81 : 471-9.
- 3. Robinson MC. An epidemic of virus disease in Southern Province, Tanganyika Territory, in 1952-53. Clinical features. Trans R Soc Trop Med Hyg 1955 ; 49 : 28.
- 4. Jeandel P, Rosse R, Durand JP. Arthrites virales exotiques : place des alphaviroses. Med Trop 2004 ; 64 : 81-8.
- 5. Simizu B, Yamamoto K, Hashimoto K, et al. Structural proteins of Chikungunya virus. J Virol 1984 ; 51 : 254-8.
- 6. Zeller H. Dengue, arbovirus et migrations dans l’Océan Indien. Bull Soc Pathol Exot 1998 ; 91 : 56-60.
- 7. Diallo M, Thonnon J, Traore-Lamizana M, Fontenille D. Vectors of Chikungunya virus in Senegal: current data and transmission cycles. Am J Trop Med Hyg 1999 ; 60 : 281-6.
- 8. Pile JC, Henchal EA, Christopher GW, et al. Chikungunya in a north american traveler. J Travel Med 1999 ; 6 : 137-9.
- 9. Zytoon EM, El-Belbasi HI, Matsumura T. Transovarial transmission of Chikungunya virus by Aedes albopictus mosquitoes ingesting microfilariae of Dirofilaria immitis under laboratory conditions. Microbiol Immunol 1993 ; 37 : 419-21.
- 10. Pastorino B, Muyembe-Tamfum JJ, Bessaud M, et al. Epidemic resurgence of Chikungunya virus in Democratic Republic of the Congo : identification of a new central african strain. J Med Virol 2004 ; 74 : 277-82.
- 11. Thonnon J, Speigel A, Diallo M, et al. Epidémies à virus Chikungunya en 1996 et 1997 au Sénégal. Bull Soc Pathol Exot 1999 ; 92 : 79-82.
- 12. Mackenzie JS, Chua KB, Daniels PW, et al. Emerging viral diseases of Southeast Asia and the Western Pacific. Emerg Infect Dis 2001 ; 7 : 497-504.
- 13. Laras K, Sukri NC, Larasati RP, et al. Tracking the re-emergence of epidemic Chikungunya virus in Indonesia. Trans R Soc Trop Med Hyg 2005 ; 99 : 128-41.
- 14. Brighton SW, Prozesky OW, De la Harpe AL. Chikungunya virus infection : a retrospective study of 107 cases. South Afr Med J 1983 ; 63 : 313-5.
- 15. Observatoire Régional de la Santé de la Réunion. Épidémie de Chikungunya à la Réunion. Point au 2 mars 2006 pour la semaine 8 allant du 20 février au 26 février 2006. http://www.orsrun.net/PEHChik020306.pdf
- 16. Morrison TE, Whitmore AC, Shabman RS, et al. Characterization of Ross River Virus tropism and virus-induced inflammation in a mouse model of viral arthritis ans myositis. J Virol 2006 ; 80 : 737-49.
- 17. Pastorino B, Bessaud M, Grandadam M, et al. Development of a TaqMan® RT-PCR assay without RNA extraction step for the detection and quantification of African Chikungunya viruses. J Virol Methods 2005 ; 124 : 65-71.
- 18. Briolant S, Garin D, Scaramozzino N, et al. In vitro inhibition of Chikungunya and Semliki Forest viruses replication by antiviral compounds : synergistic effect of interferon-α and ribavirin combination. Antiviral Res 2004 ; 61 : 111-7.
- 19. Edelman R, Tacket CO, Wasserman SS, et al. Phase II safety and immunogenicity study of live Chikungunya virus vaccine TSI-GSD-218. Am J Trop Med Hyg 2000 ; 62 : 681-5.
- 20. Institut National de Veille Sanitaire. Épidémie de Chikungunya à la Réunion/Océan Indien. Point de situation au 24 février 2006. http://www.invs.sante.fr/presse/2006/ le_point_sur/chikungunya_240206/index.html
List of figures
Figure 1
Aedes albopictus, le moustique en cause dans l’épidémie réunionnaise.
List of tables
Tableau I
Épidémie de Chikungunya dans l’Océan Indien en 2005-2006, au 28 février 2006.
Les chiffres indiqués correspondent au nombre de cas suspects ou estimés (pour la Réunion). Ils sont tirés des sites Internet de l’Institut de Veille Sanitaire et de Promed, au 7 mars 2006.
*Données non disponibles (cas décrits, non quantifiés précisément).