Article body
La migration des oiseaux a toujours intrigué les biologistes. Elle les intrigue toujours aujourd’hui. Par exemple, ils se demandent comment les oiseaux qui migrent la nuit au-dessus du Sahara [1] parviennent à équilibrer leur budget thermique lorsque la température de l’air y dépasse 20 °C. À ce sujet, des études effectuées en soufflerie ont montré que le vol prolongé était impossible lorsque la température de l’air dépassait 10 °C, pour cause d’hyperthermie ou de déshydratation. Ces résultats paradoxaux ont fait dire à Biesel et Nachtigall [2] que, dans des conditions naturelles, les oiseaux devaient profiter d’un mécanisme de thermorégulation dont ne tenaient pas compte les conditions d’une soufflerie. Vingt ans plus tard, ce mécanisme demeure toujours aussi mystérieux.
L’ennemi n° 1
Conçu d’abord pour assurer le confort par temps froid et venteux, le plumage de l’oiseau constitue un isolant thermique remarquable. Toutefois, il devient un sérieux handicap lorsque l’oiseau doit s’adonner par temps chaud à un exercice aussi intense et prolongé qu’un vol migratoire typique. Le vol battu exige, en effet, de décupler la dépense énergétique. Plus de 80 % de l’énergie consommée apparaît sous forme de chaleur corporelle que l’oiseau doit dissiper intégralement s’il veut maintenir une température stable. Cette chaleur peut être perdue suivant trois modes : l’évaporation mettant en cause l’eau, le rayonnement où entrent en jeu les photons et la convection prenant en compte l’air en mouvement. Si l’on se réfère aux études classiques réalisées en soufflerie, les deux premiers modes expliquent chacun la perte d’environ 10 % de la chaleur produite et le troisième les 80 % restants [2-4]. Les 10 % évaporatoires constituent toutefois un maximum absolument essentiel pour éviter la déshydratation, de sorte que la marge d’ajustement aux conditions ambiantes réside fondamentalement dans l’exploitation des phénomènes convectifs et radiatifs.
L’intervention céleste
Tous les corps réels émettent de l’énergie par rayonnement photonique. Toutefois, si les photons hautement énergétiques émis par le soleil impressionnent facilement les rétines humaines, ceux de plus basse énergie émis dans l’infrarouge moyen par des objets à température voisine de la température ambiante échappent à la perception sensorielle directe.
Cela nous rend inconscient de l'influence marquée qu'a l'humidité atmosphérique sur l'énergie infrarouge émise par le ciel vers la terre. Maximale par temps couvert, cette énergie est typiquement réduite par temps clair à celle qu'émettrait un corps noir 20 °C plus froid que l'air. Un déficit de température de cette ampleur fait d'un ciel clair nocturne un puits radiatif capable de refroidir significativement les objets qui lui sont exposés. Ce pouvoir de refroidissement permet de comprendre pourquoi les nuits sont souvent plus froides par temps dégagé, aussi bien dans les déserts chauds que dans les régions tempérées. On lui attribue également en automne les gelées au sol si redoutées des maraîchers puisqu'elles peuvent apparaître sournoisement par nuit claire, alors que la température de l'air est demeurée bien au-dessus du point de congélation de l'eau.
L’exposition à un ciel clair pourrait-elle refroidir la surface du plumage d’un oiseau en vol et ainsi faciliter l’évacuation de la chaleur transportée des muscles du vol à la peau par le sang ? Pour répondre à cette question, nous ne pouvions pas avoir recours aux souffleries conventionnelles puisque leurs parois, toujours isothermes à l’air, ne peuvent reproduire le déficit ciel-air. Nous avons donc construit une soufflerie spéciale, et sans doute unique, dont la paroi supérieure pouvait être refroidie avec de la glace sèche jusqu’à 50 °C en dessous de la température de l’air, même à des vitesses de vent atteignant les 75 km/h. Cela a représenté un défi assez éprouvant, spécialement pour nous, biologistes non férus de génie mécanique et, de surcroît, non subventionnés. Comme la taille de notre soufflerie ne permettait pas d’y faire voler des pigeons, nous avons soutenu nos sujets en position de vol plané et nous les avons placés dans un état thermique semblable à celui associé au vol battu, en les exposant à des micro-ondes d’une intensité comparable à celle utilisée en physiothérapie.
D’après nos résultats, lorsque l’écart entre la température du plumage et celle de l’air est suffisant, la convection permet une dissipation de chaleur adéquate quelle que soit la température du ciel. En revanche, lorsque l’air dépasse les 15 °C, l’écart plumage-air diminue au point où la convection ne suffit plus, même la nuit, à garder la température du plumage assez basse pour assurer un bon refroidissement cutané. L’effet refroidissant du ciel devient alors critique. Nos résultats montrent qu’il peut rafraîchir significativement la surface du plumage, même si la chute de la température des plumes diminue forcément l’écart plumage-air et donc la perte convective. L’effet était suffisamment fort dans nos conditions pour que l’importance du rayonnement puisse dépasser celle de la convection à des valeurs habituelles du déficit ciel-air (Figure 1). À des valeurs extrêmes du déficit, nous avons même observé des écarts plumage-air négatifs, signalant que l’air chauffait alors les plumes.
Figure 1
Influence de la température du ciel sur les contributions respectives du rayonnement et de la convection à la dissipation non évaporatoire de la chaleur vers le haut chez un pigeon en vol simulé, à une température d’air de 25 °C et à une vitesse de vent de 10 m/s.
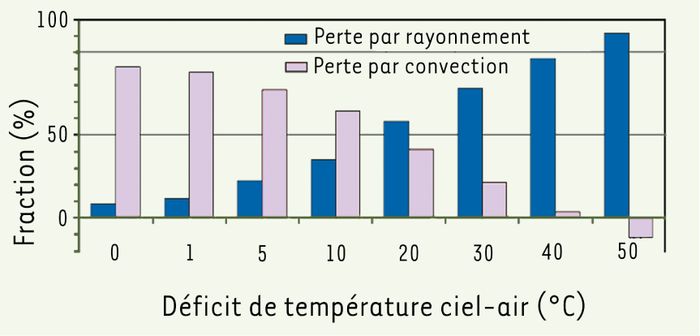
Les valeurs ont été prédites par le biais d’un modèle de régression multiple (r2 = 0,96) appliqué à l’ensemble de nos mesures de température superficielle du plumage faites par infrarouge [5]. Un déficit ciel-air de 0 °C correspond à l’ambiance rayonnante d’une soufflerie conventionnelle ; un déficit de 20 °C correspond aux conditions moyennes observées dans la nature. À des déficits supérieurs à 42 °C, la fraction rayonnante dépasse 100 % parce que l’énergie radiante comprend la chaleur que le plumage gagne par contact avec l’air en mouvement (convection).
La fin de l’histoire ?
De telles mesures montrent que le pouvoir refroidissant du ciel peut être important pour l’équilibre thermique de l’oiseau migrateur dans des conditions où la température superficielle du plumage constitue une limite à la dissipation de la chaleur produite par ses muscles. Nos résultats aident donc à résoudre le paradoxe évoqué plus haut, et ils contribuent aussi à expliquer pourquoi la majorité des vols migratoires ont lieu par nuit claire. Toutefois, l’évaluation précise de la contribution du pouvoir refroidissant du ciel à la thermorégulation des oiseaux pendant la migration ne sera possible qu’au moment où nous disposerons de mesures obtenues chez des sujets volant librement en milieu naturel.
Appendices
Références
- 1. Klaassen M, Biebach H. Flight altitude of trans-Sahara migrants in autumn: a comparison of radar observations with predictions from meteorological and water and energy balance models. J Exp Biol 2000 ; 31 : 47-55.
- 2. Biesel W, Nachtigall W. Pigeon flight in a wind tunnel. IV. Thermoregulation and water homeostasis. J Comp Physiol 1987 ; 157B : 117-28.
- 3. Torre-Bueno JR. Evaporative cooling and water balance during flight in birds. J Exp Biol 1978 ; 75 : 231-6.
- 4. Ward S, Rayner JMV, Möller U, et al. Heat transfer from starlings Sturnus vulgaris during flight. J Exp Biol 1999 ; 202 : 1589-602.
- 5. Léger J, Larochelle J. On the importance of radiative heat exchange during nocturnal flight in birds. J Exp Biol 2006; 209 : 103-14.
List of figures
Figure 1
Influence de la température du ciel sur les contributions respectives du rayonnement et de la convection à la dissipation non évaporatoire de la chaleur vers le haut chez un pigeon en vol simulé, à une température d’air de 25 °C et à une vitesse de vent de 10 m/s.
Les valeurs ont été prédites par le biais d’un modèle de régression multiple (r2 = 0,96) appliqué à l’ensemble de nos mesures de température superficielle du plumage faites par infrarouge [5]. Un déficit ciel-air de 0 °C correspond à l’ambiance rayonnante d’une soufflerie conventionnelle ; un déficit de 20 °C correspond aux conditions moyennes observées dans la nature. À des déficits supérieurs à 42 °C, la fraction rayonnante dépasse 100 % parce que l’énergie radiante comprend la chaleur que le plumage gagne par contact avec l’air en mouvement (convection).