Article body
Les microtubules sont des constituants essentiels de l’architecture dynamique de la cellule. Ils sont impliqués dans de nombreuses fonctions cellulaires fondamentales telles que la division cellulaire, le trafic intracellulaire ou la croissance des neurones. Ainsi, pour ce qui est de la division cellulaire, les microtubules s’organisent pour former le fuseau mitotique dont ils sont l’élément de base. Les microtubules sont des cylindres creux dont les parois sont constituées de polymères de tubuline, une protéine cellulaire hétérodimérique abondante comprenant une sous-unité α et une sous-unité β. Ces polymères linéaires et parallèles sont appelés protofilaments. In vivo, les microtubules sont constitués majoritairement de treize protofilaments. La cohésion des microtubules est assurée par des contacts longitudinaux entre molécules de tubuline dans les protofilaments, et par des contacts latéraux entre tubulines de protofilaments voisins. Dans la cellule, la dynamique d’assemblage et de désassemblage de la tubuline en microtubules est contrôlée par de nombreuses protéines endogènes (pour revue, voir [1]). Elle peut aussi être perturbée par des composés pharmacologiques exogènes qui, en affectant la formation du fuseau mitotique, arrêtent la division cellulaire et conduisent à l’apoptose [2].
La vinblastine est l’un de ces composés ; elle a été utilisée avec succès depuis plusieurs dizaines d’années pour traiter certains cancers et est à l’origine de l’inversion du pronostic de leucémies infantiles. Bien que l’on sache depuis longtemps que la vinblastine cible la tubuline, son site de fixation était inconnu et son mécanisme d’action largement incompris. L’une des raisons des difficultés de l’étude du mécanisme d’action de la vinblastine est qu’elle agrège la tubuline et donc que les solutions de tubuline et de vinblastine sont hétérogènes [3]. En déterminant par radiocristallographie la structure de la vinblastine liée à la tubuline, nous avons identifié le site de fixation de cet alcaloïde et expliqué comment la vinblastine induit la formation de spirales de tubuline aux dépens des microtubules [4].
Pour ce faire, nous avons incubé avec de la vinblastine des cristaux d’un complexe ternaire composé du domaine de type stathmine de la protéine RB3 (RB3-SLD) et de deux hétérodimères de tubuline (ce complexe est appelé T2R). RB3 est une protéine de la famille de la stathmine, une phosphoprotéine dont il a été proposé qu’elle serve de relais intégrateur des voies de signalisation [5]. Comme la stathmine, la protéine RB3 séquestre la tubuline pour former un complexe qui n’est pas incorporé dans les microtubules [6, 7]. L’ensemble des données biochimiques montre que la vinblastine se fixe au complexe tubuline-RB3-SLD de la même façon qu’à la tubuline. Dans le complexe T2R-vinblastine, la vinblastine se fixe à l’interface des deux hétérodimères de tubuline, en contact avec la sous-unité α de l’un et la sous-unité β de l'autre (Figure 1).
Figure 1
Structure du complexe tubuline-RB3-SLD-vinblastine.
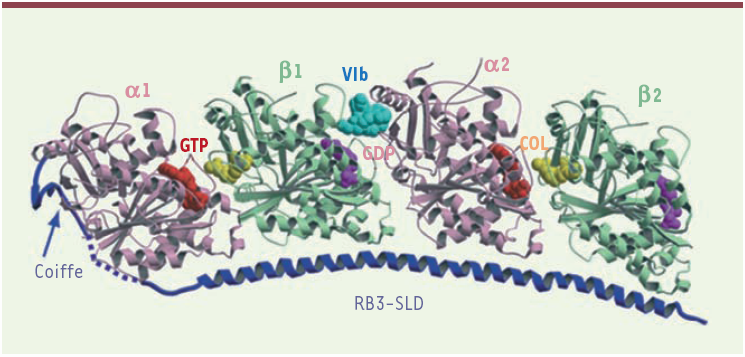
La vinblastine (cyan, Vlb) est située à l’interface entre deux hétérodimères de tubuline (α1-β1 et α2-β2), chaque monomère complexant un nucléotide (un GTP sur les sous-unités α et un GDP sur les sous-unités β). Outre ces hétérodimères de tubuline, le complexe comprend le domaine de type stathmine de la protéine RB3. Une molécule de colchicine (un autre médicament ciblant la tubuline, représenté en jaune et noté COL) est liée à chaque sous-unité β de la tubuline. La colchicine, ainsi que les nucléotides, sont représentés par le volume que leurs atomes occupent dans l’espace. Les protéines (tubuline et RB3-SLD) sont représentées par des flèches (brins de feuillets β) et des hélices (hélices α) reliées par un ruban fin qui suit la chaîne polypeptidique ; les tirets représentent une zone de la protéine RB3-SLD mal définie dans le cristal et probablement mal ordonnée.
L’analyse des résidus de la tubuline interagissant avec la vinblastine montre qu’une partie d’entre eux est impliquée dans les contacts longitudinaux des microtubules. En se fixant à la tubuline, la vinblastine empêche certains de ces contacts de s’établir et agit comme un coin qui force la formation de protofilaments courbes, par opposition aux protofilaments droits qui constituent les parois des microtubules. De ce fait, les contacts latéraux entre protofilaments, qui sont cruciaux pour la stabilité des microtubules, ne peuvent plus s’établir, avec pour conséquence principale le fait qu’à forte concentration de vinblastine, lorsque la perte de contacts microtubulaires devient importante, la tubuline s’assemble en spirales aux dépens de la formation de microtubules. Dans ces spirales, la liaison entre hétérodimères de tubuline est renforcée par des contacts avec des molécules de vinblastine, chaque molécule de vinblastine interagissant avec la sous-unité α d'un hétérodimère et la sous-unité β de l'autre, comme dans le complexe tubuline-RB3-SLD-vinblastine [4].
D’un point de vue plus proche des applications, le complexe T2R est un outil précieux pour l’étude biochimique du mécanisme d’action de la vinblastine et des composés qui se fixent au même site de la tubuline. En effet, comme on l’a vu, la vinblastine se fixe à T2R comme à la tubuline ; de plus, elle ne l’agrège pas. La structure que nous avons déterminée permet d’expliquer cette propriété : une partie de RB3-SLD, appelée coiffe (Figure 1) [8], occupe le site de la vinblastine sur la sous-unité α1 de la tubuline dans le complexe [4]. La vinblastine ne peut donc pas se fixer simultanément à deux complexes et n’agrège pas T2R.
La structure que nous avons déterminée clarifie le mécanisme d’action de la vinblastine. Elle ouvre la voie à la conception rationnelle de nouveaux composés à visée thérapeutique se fixant au même site de la tubuline que ce médicament. Le défi à relever dans cette démarche est de conserver l’efficacité de la vinblastine comme composé antimitotique mais d’en diminuer, voire d’en éliminer, deux propriétés indésirables : sa neurotoxicité et sa susceptibilité aux mécanismes de résistance de la cellule aux composés exogènes [2].
Appendices
Références
- 1. Desai A, Mitchison TJ. Microtubule polymerization dynamics. Ann Rev Cell Dev Biol 1997 ; 13 : 83-117.
- 2. Jordan MA, Wilson L. Microtubules as a target for anticancer drugs. Nat Rev Cancer 2004 ; 4 : 253-65.
- 3. Weisenberg RC, Timasheff SN. Aggregation of microtubule subunit protein. Effects of divalent cations, colchicine and vinblastine. Biochemistry 1970 ; 9 : 4110-6.
- 4. Gigant B, Wang C, Ravelli RB, et al. Structural basis for the regulation of tubulin by vinblastine. Nature 2005 ; 435 : 519-22.
- 5. Sobel A. Stathmin: a relay phosphoprotein for multiple signal transduction ? Trends Biochem Sci 1991 ; 16 : 301-5.
- 6. Gigant B, Curmi PA, Martin-Barbey C, et al. The 4 A X-ray structure of a tubulin: stathmin-like domain complex. Cell 2000 ; 102 : 809-16.
- 7. Charbaut E, Curmi PA, Ozon S, et al. Stathmin family proteins display specific molecular and tubulin binding properties. J Biol Chem 2001 ; 276 : 16146-54.
- 8. Ravelli RB, Gigant B, Curmi PA, et al. Insight into tubulin regulation from a complex with colchicine and a stathmin-like domain. Nature 2004 ; 428 : 198-202.
List of figures
Figure 1
Structure du complexe tubuline-RB3-SLD-vinblastine.
La vinblastine (cyan, Vlb) est située à l’interface entre deux hétérodimères de tubuline (α1-β1 et α2-β2), chaque monomère complexant un nucléotide (un GTP sur les sous-unités α et un GDP sur les sous-unités β). Outre ces hétérodimères de tubuline, le complexe comprend le domaine de type stathmine de la protéine RB3. Une molécule de colchicine (un autre médicament ciblant la tubuline, représenté en jaune et noté COL) est liée à chaque sous-unité β de la tubuline. La colchicine, ainsi que les nucléotides, sont représentés par le volume que leurs atomes occupent dans l’espace. Les protéines (tubuline et RB3-SLD) sont représentées par des flèches (brins de feuillets β) et des hélices (hélices α) reliées par un ruban fin qui suit la chaîne polypeptidique ; les tirets représentent une zone de la protéine RB3-SLD mal définie dans le cristal et probablement mal ordonnée.