Abstracts
Résumé
Les maladies cardiovasculaires (MCV) sont devenues l’une des causes majeures de mortalité dans les sociétés occidentales. De nombreux facteurs de risque sont associés à ces maladies, parmi lesquelles le diabète, l’obésité, la résistance à l’insuline, les dyslipidémies et l’hypertension. La prise en charge de ces maladies est devenue un enjeu majeur de santé publique, qui a donné lieu au développement d’un arsenal thérapeutique destiné à lutter contre ces anomalies. Les fibrates permettent de corriger efficacement les dyslipidémies et diminuent le risque d’accidents cardiovasculaires. Les thiazolidinediones (TZD) ou glitazones agissent efficacement sur le diabète en augmentant la sensibilité à l’insuline et en diminuant la glycémie. Ces deux familles de médicaments agissent au niveau moléculaire en activant les récepteurs nucléaires peroxisome proliferator-activated receptors (PPAR), qui jouent un rôle central dans la régulation du métabolisme lipidique et le contrôle de la glycémie. Dans cet article, nous aborderons le mode d’action des fibrates et des TZD et présenterons les nouvelles molécules en développement ayant pour cible pharmacologique les PPAR.
Summary
Cardiovascular diseases (CVD) remain the leading cause of mortality in the western societies. Several risk factors predispose to CVD including diabetes, obesity, insulin resistance, dyslipidemia and hypertension. Various pharmacological therapies have been developped to control the risk factors associated to CVD. Fibrates are able to correct dyslipidemia, therefore decreasing CVD risk. Thiazolidinediones (TZD) or glitazones by increasing insulin sensitivity decrease plasma glucose levels in diabetic patients. Both fibrates and TZD activate the peroxisome proliferator-activated receptors (PPARs), a family of nuclear receptors that play a central role in the control of lipid and glucose metabolism. In this review, we will discuss the mode of action of fibrates and TZD and we will present an overview on PPAR ligands under development.
Article body
Le diabète de type 2 est caractérisé par une hyperglycémie due à une résistance à l’insuline et à une insuffisance pancréatique [1]. Le syndrome de résistance à l’insuline, ou syndrome métabolique, qui précède l’établissement du diabète de type 2, est généralement associé à une dyslipidémie caractérisée par des concentrations plasmatiques élevées de triglycérides et particules de LDL (low density lipoprotein) petites et denses, et par des concentrations abaissées en cholestérol-HDL (high density lipoprotein) [2]. Ce profil est associé à un risque élevé de maladie cardiovasculaire (MCV) chez ces patients [1]. La prise en charge pharmacologique est un moyen de contrôler le développement des complications diabétiques. Ainsi, les fibrates et les TZD améliorent la dyslipidémie et la sensibilité à l’insuline, permettant de réduire efficacement le risque de MCV.
Les fibrates et les TZD sont des ligands synthétiques des récepteurs nucléaires peroxisome proliferator-activated receptors (PPAR). Trois isotypes de PPAR ont été décrits: α, δ(β) et γ. Une fois activés par leurs ligands, les PPAR forment des hétérodimères avec le récepteur nucléaire RXR (9-cis retinoic acid receptor) et modulent la transcription après s’être fixés sur des séquences spécifiques PPRE (peroxisome proliferator response elements) localisées dans les régions régulatrices de leurs gènes cibles. Ainsi, l’activation des PPAR peut contrôler l’expression des gènes impliqués dans le métabolisme des lipides et du glucose [3, 4].
La possibilité de corriger les anomalies métaboliques du diabète en activant les PPAR a conduit à un grand effort de recherche sur leurs agonistes. Cette revue analysera les aspects généraux du mécanisme d’action des agonistes de PPARα et PPARγ en clinique et leur utilisation vis-à-vis des complications diabétiques. Enfin, les avancées réalisées en termes de nouveaux agonistes mixtes PPARα/γ et leur utilité potentielle pour le traitement du diabète seront présentées.
Agonistes PPARα
Les fibrates tels que le fénofibrate ou le gemfibrozil sont utilisés cliniquement comme hypolipémiants. Ils ont été identifiés comme des ligands de PPARα. Ainsi, chez l’homme, l’activation de PPARα par les fibrates se traduit par une diminution des concentrations plasmatiques de triglycérides (TG) et une augmentation des niveaux de cholestérol-HDL.
La baisse des concentrations plasmatiques des triglycérides provient de la mise en oeuvre de différents mécanismes. D’une part, la lipolyse des VLDL (very low density lipoprotein) est stimulée: en effet, la transcription de la LPL (lipoprotéine lipase) est augmentée, tandis que l’expression de l’apolipoprotéine C-III (apoC-III) est diminuée. D’autre part, la capture des acides gras (AG) par la cellule et leur catabolisme intracellulaire sont stimulés: en effet, l’expression de FATP1 (fatty acid transport protein 1) et FAT/CD36 (fatty acid translocase), mais également de l’ACS (acyl-CoA synthétase) et de la CPT-I (carnitine palmitoyl acyl transférase-I) est augmentée. Ces effets entraînent une moindre disponibilité des AG pour la synthèse des TG et, par conséquent, des lipoprotéines riches en triglycérides [4]. Récemment, il a été démontré que l’activation de PPARα augmente l’expression de l’apoA-V [5], ce qui suggère un nouveau mécanisme par lequel PPARα abaisse la concentration des TG.
L’activation de PPARα est associée à une augmentation de la concentration du cholestérol-HDL due à l’augmentation de l’expression de l’apoA-I et de l’apoA-II, les principales apolipoprotéines présentes dans les HDL. Parallèlement, l’activation de PPARα entraîne une augmentation de l’expression des transporteurs ABCA-1 (ATP-binding cassette transporter A-1) et SR-BI/CLA-1 (scavenger receptor-BI) dans les macrophages [6, 7], un processus qui permet l’efflux de l’excès de cholestérol vers les HDL et son retour au foie via le « transport inverse du cholestérol ».
De nombreuses études suggèrent que le traitement par les fibrates réduit les facteurs de risque cardiovasculaire associés aux dyslipidémies chez le patient diabétique. L’étude VA-HIT (veterans affairs-high density lipoprotein cholesterol intervention trial), dans laquelle 25% des patients analysés sont diabétiques, montre une diminution du risque de MCV liée à une augmentation de cholestérol-HDL et une diminution des TG [8]. De plus, l’étude DAIS (diabetes atherosclerosis intervention study) a montré que le fénofibrate réduit la progression de l’athérosclérose des artères coronaires, ce qui peut être expliqué partiellement par la correction de la dyslipidémie diabétique [9]. Ces résultats, ainsi que ceux d’autres études [10, 11], suggèrent que les fibrates constituent un traitement efficace permettant de réduire le risque de MCV associé à la dyslipidémie diabétique. De plus, des études en cours, telles que le fenofibrate intervention and event lowering in diabetes (FIELD) trial, devraient permettre d’évaluer l’effet réel des fibrates sur le risque de MCV et la mortalité dans le diabète de type 2.
Agonistes PPARγ
Les TZD (la pioglitazone et la rosiglitazone) sont une classe de médicaments antidiabétiques oraux réduisant l’hyperglycémie et améliorant le profil lipidique chez les patients diabétiques de type 2. Un certain nombre d’arguments suggèrent que l’effet antidiabétique des TZD s’exerce via PPARγ. D’une part, il existe une excellente corrélation entre les capacités de liaison de PPARγ à son ligand in vitro et son effet hypoglycémiant in vivo. D’autre part, des mutations dans le gène de PPARγ ont été associées au développement de la résistance à l’insuline et du diabète [12].
PPARγ est exprimé pendant la différenciation des préadipocytes en adipocytes. Ainsi, l’activation de PPARγ par les TZD induit l’expression des gènes tels que LPL, FATP, CD36 et ACS, ce qui entraîne la lipolyse des TG plasmatiques et leur stockage dans les adipocytes [3, 13]. Par ailleurs, les TZD induisent l’expression de la glycérol-kinase (GyK) adipocytaire, qui stimulerait l’incorporation du glycérol dans les triglycérides et réduirait la sécrétion d’acides gras libres par les adipocytes [14]. Ces mécanismes contribueraient à réduire la quantité d’acides gras et de TG circulants.
Plusieurs études montrent que les TZD induisent une redistribution des graisses du compartiment viscéral vers le compartiment sous-cutané, ce qui est associé à une diminution de la concentration circulante d’AG libres [13]. De plus, les adipocytes produits en réponse aux TZD sont plus petits, et donc plus sensibles à l’action de l’insuline [15].
Selon le modèle de lipid stealing, le développement de l’insulinorésistance est lié à une mauvaise répartition des acides gras entre le tissu adipeux et les tissus périphériques tels que le foie et le muscle, où ils s’accumulent. La diminution de la libération d’AG libres améliore l’utilisation du glucose par les muscles (cycle glucose-acide gras), ce qui expliquerait l’effet hypoglycémiant des TZD.
Une augmentation de la concentration de TNFα (tumor necrosis factor α) et d’IL-6 (interleukine 6) circulante a été associée à la résistance à l’insuline [13, 16], mettant en évidence l’effet de certaines cytokines dans le développement de l’insulinorésistance. Il a été proposé que le TNFα inhibe la transduction du message induit par l’insuline, par l’intermédiaire d’un mécanisme impliquant une diminution de l’autophosphorylation du récepteur de l’insuline et une augmentation de la phosphorylation d’IRS (insulin receptor substrate) dans le tissu adipeux. De plus, le TNFα augmente la production des AG libres ainsi que l’expression de la leptine dans le tissu adipeux, ce qui peut également avoir un rôle indirect dans le développement de la résistance à l’insuline des muscles et du foie [13].
L’adiponectine est une protéine fortement exprimée dans le tissu adipeux. Elle augmente l’oxydation des acides gras dans le muscle squelettique et diminue la production de glucose par le foie, ce qui entraîne une diminution de la concentration d’AG libres circulants, des triglycérides et du glucose, tous ces effets pouvant entraîner une amélioration de la sensibilité à l’insuline et une diminution de l’athérosclérose. De plus, l’adiponectine abaisse la synthèse des molécules d’adhésion par les cellules endothéliales et inhibe la réponse inflammatoire [17]. Les niveaux sériques d’adiponectine sont diminués chez des patients obèses ou atteints du diabète de type 2 (voir l’article de C.Lacquemant et al., p.809 de ce numéro). Selon une étude récente [18], l’administration de rosiglitazone aux patients atteints d’un diabète type 2 entraînerait une augmentation de la concentration plasmatique d’adiponectine. Étant donné les effets anti-inflammatoires et anti-athérosclérotiques de l’adiponectine, le rôle des TZD sur l’expression de l’adiponectine pourrait contribuer aux effets bénéfiques de PPARγ sur l’inflammation et l’athérogenèse [17].
Les TZD, par le biais de leur activation de PPARγ, induisent l’expression des transporteurs ABCA-1 et SR-BI, ainsi que de l’apoE, dans le macrophage, ce qui pourrait avoir des effets bénéfiques face au développement de la lésion athérosclérotique chez l’homme [4].
Perspectives thérapeutiques
Modulateurs sélectifs de PPAR (SPPARM)
Parmi les TZD, toutes connues pour être des agonistes de PPARγ, il a été décrit des molécules agissant comme agonistes sélectifs de PPARγ [19]. Ainsi, il existe une catégorie de gènes communs aux TZD, régulés de manière identique par chaque TZD, et une catégorie de gènes régulés différemment par certaines TZD. Comment expliquer l’absence d’effet d’une TZD sur un gène donné, alors que son expression est modifiée par une autre TZD agissant pourtant sur le même récepteur nucléaire ?On sait que chaque TZD a une affinité différente pour le site de liaison aux ligands sur PPARγ, ce qui peut entraîner une conformation tridimensionnelle du complexe PPARγ-TZD variable selon chaque molécule. Ces différentes conformations peuvent aboutir au recrutement de cofacteurs distincts et, par conséquent, entraîner une régulation transcriptionnelle particulière pour chaque complexe. Ainsi, divers ligands peuvent, via un même récepteur nucléaire, induire des réponses biologiques différentes (Figure 1). Ce phénomène est à la base du concept de modulateur sélectif de PPAR, ou SPPARM (selective PPAR modulator) [20]. Les SPPARM de PPARγ ont pour objectif de dissocier l’effet sur la différentiation adipocytaire de l’effet sur le métabolisme de glucose. Ainsi, le LG100641 est un ligand de PPARγ capable de contrecarrer l’effet des TZD sur la différenciation adipocytaire, mais capable parallèlement d’augmenter la capture du glucose par ces cellules [21].
Figure 1
Modulateurs sélectifs de PPAR (selective PPAR modulator ou SPPARM).
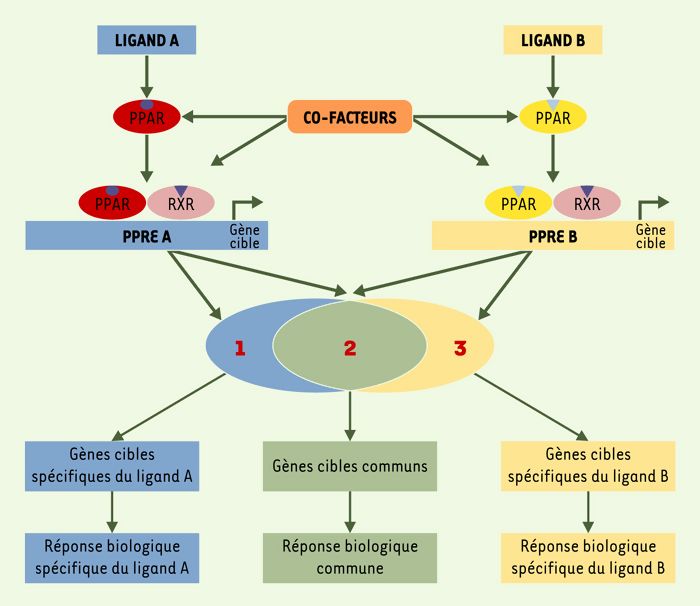
Chaque agoniste de PPAR (peroxisome proliferator-activated receptors), ligand A ou ligand B, peut entraîner une conformation tridimensionnelle différente du complexe PPAR-ligand. Ces conformations différentes, après liaison des hétérodimères avec le récepteur nucléaire RXR (9-cis retinoic acid receptor) et fixation sur des séquences spécifiques PPRE (peroxisome proliferator response elements), peuvent amener à un recrutement de cofacteurs différents et, par conséquent, entraîner une régulation transcriptionnelle particulière pour chaque complexe. Ainsi, les différents ligands peuvent, via un même récepteur nucléaire, induire des réponses biologiques communes, mais aussi des réponses biologiques qui leur sont spécifiques.
Antagonistes de PPARγ
Par ailleurs, des antagonistes de PPARγ ont été développés. Le GW0072 est une molécule appartenant à la famille des TZD capable d’inhiber la différenciation adipocytaire [22]. Récemment, une nouvelle classe d’antagonistes spécifiques de PPARγ, n’appartenant pas à la famille des TZD, a été décrite : SR-202 est un dérivé de l’acide para-chlorobenzylique qui contrecarre l’effet transcriptionnel de PPARγ induit par des TZD et inhibe la différenciation adipocytaire induite par les TZD ou les mélanges hormonaux des cultures cellulaires 3T3-L1 [23]. Peu d’essais ont été réalisés in vivo avec ces composés. Toutefois, le traitement par SR-202 prévient l’insulino-résistance induite par un régime hyperlipidique et améliore le profil lipidique chez la souris. De plus, il diminue l’adiposité de souris sauvages ayant un régime normal ou hyperlipidique [23]. On constate également, chez les souris traitées par le SR-202, des concentrations d’adipokines, leptine et TNFα diminuées, ce qui est cohérent avec la diminution de taille des adipocytes [23]. Ce phénomène pourrait participer à l’amélioration de la sensibilité à l’insuline.
Co-agonistes PPARα/γ
Étant donné que les effets des TZD sur le profil cardiovasculaire, et notamment sur le profil lipidique du patient diabétique sont insuffisants, l’activation conjointe de PPARα et de PPARγ pourrait permettre d’obtenir des effets complémentaires et synergiques, concernant à la fois l’amélioration du métabolisme des lipides et celle de la sensibilité à l’insuline et de l’utilisation du glucose. De plus, l’activation conjointe de PPARα et PPARγ pourrait limiter les effets secondaires liés au traitement par les TZD actuellement utilisées. C’est ainsi qu’une nouvelle famille de molécules, les co-agonistes PPARα/γ, a été développée (Figure 2; Tableau I).
Figure 2
Co-agonistes PPARα/γ.
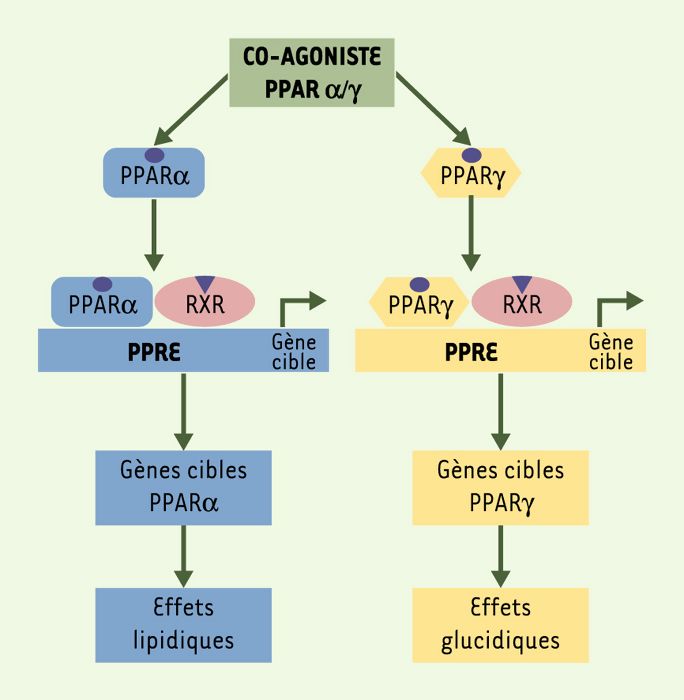
Les effets liés à l’activation des PPAR peuvent être complémentaires et synergiques. Ainsi, une nouvelle famille de molécules capables d’activer parallèlement PPARα et PPARγ a été développée. Elle peut entraîner à la fois des effets bénéfiques sur le métabolisme des lipides, via l’activation de PPARα et aboutir à l’amélioration de la sensibilité à l’insuline et de l’utilisation du glucose, via l’activation de PPARγ.
Tableau I
Ligands de PPARγ et co-agonistes PPARα/γ en développement.
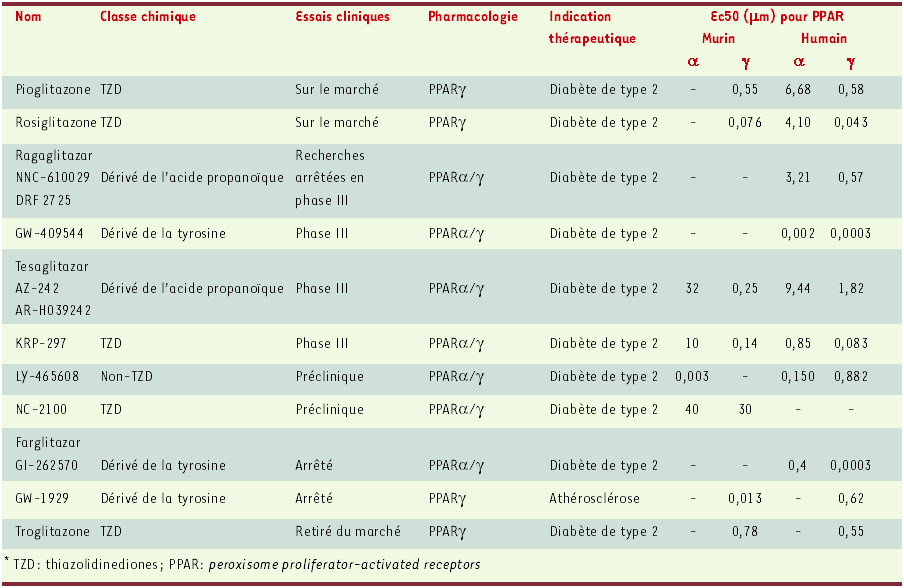
Dans des modèles animaux de résistance à l’insuline, on constate que des co-agonistes PPARα/γ diminuent les concentrations de triglycérides circulants [24], effet également observé chez des souris transgéniques pour l’Apo A-I humaine [25]. Ce phénomène peut être associé à l’augmentation de l’activité de la LPL adipocytaire [26], ainsi qu’à l’augmentation de la β-oxydation et à la diminution de la lipogenèse hépatique [27]. Par ailleurs, les co-agonistes PPARα/γ augmentent la concentration en HDL plasmatique [25], par un mécanisme probablement associé à une augmentation de la synthèse de l’apoA-I et à une augmentation de l’activité LCAT (lécithine : cholestérol-acyltransférase).
Les effets des co-agonistes PPARα/γ sur la glycémie et la sensibilité à l’insuline sont comparables à ceux des TZD. Dans des modèles d’insulinorésistance, les co-agonistes PPARα/γ diminuent la concentration en glucose plasmatique [24, 28], phénomène associé à une augmentation de la sensibilité périphérique à l’insuline [25, 29] et à une diminution de l’activité des enzymes de la néoglucogenèse [29]. Les co-agonistes PPARα/γ ne modifient pas la sécrétion d’insuline, même s’ils préviennent la réduction du contenu insulinique des îlots pancréatiques observée dans le diabète [28]. L’augmentation de la sensibilité à l’insuline par les co-agonistes PPARα/γ chez le rongeur serait plutôt la conséquence de la diminution des concentrations en acides gras circulants.
Le traitement avec des co-agonistes PPARα/γ peut entraîner une diminution de la masse adipocytaire viscérale [29]. Plusieurs mécanismes peuvent expliquer l’absence de développement de la masse adipeuse viscérale chez l’animal traité par les co-agonistes PPARα/γ, alors même qu’ils activent PPARγ dont le rôle dans la différenciation adipocytaire a été établi. Une possibilité pourrait impliquer l’augmentation de l’oxydation des acides gras via PPARα [30]. Une autre peut impliquer un déficit d’adipogenèse de novo; cette hypothèse semble moins probable car des études réalisées chez des animaux traités avec des agonistes PPARγ ou co-agonistes PPARα/γ montrent un effet équivalent sur l’adipogenèse [29]. Cependant, tout cela reste à confirmer.
Conclusions
Les effets bénéfiques des agonistes de PPARα et PPARγ sont maintenant bien connus. Cependant, afin d’améliorer encore le profil pharmacologique, une nouvelle génération de molécules a été développée. Ainsi, les SPPARM pourraient dissocier les effets de PPARγ en diminuant l’adipogenèse, mais en conservant les effets hypoglycémiants. Par ailleurs, les co-agonistes PPARα/γ combinent les effets hypolipémiants et hypoglycémiants observés séparément avec des agonistes PPARα et PPARγ. Même si les résultats observés jusqu’à présent sont préliminaires, les études actuellement en cours devraient permettre d’évaluer le bénéfice réel de ces nouvelles familles de molécules dans le traitement du diabète.
Appendices
Remerciements
Nous tenons à remercier particulièrement Karsten Wassermann pour sa collaboration à la rédaction de cet article.
Références
- 1. Beckman JA, Creager MA, Libby P. Diabetes and atherosclerosis: epidemiology, pathophysiology, and management.JAMA 2002; 287: 2570-81.
- 2. Haffner SM. Lipoprotein disorders associated with type 2 diabetes mellitus and insulin resistance.Am J Cardiol 2002; 90: 55i-61i.
- 3. Berger J, Moller DE. The mechanisms of action of PPARs.Annu Rev Med 2002; 53: 409-35.
- 4. Barbier O, Torra IP, Duguay Y, et al. Pleiotropic actions of peroxisome proliferator-activated receptors in lipid metabolism and atherosclerosis.Arterioscler Thromb Vasc Biol 2002; 22: 717-26.
- 5. Vu-Dac N, Gervois P, Jakel H, et al. Apolipoprotein A5, a crucial determinant of plasma triglyceride levels, is highly responsive to PPAR alpha activators.J Biol Chem 2003; 278: 17982-5.
- 6. Chinetti G, Gbaguidi FG, Griglio S, et al. CLA-1/SR-BI is expressed in atherosclerotic lesion macrophages and regulated by activators of peroxisome proliferator-activated receptors.Circulation 2000; 101: 2411-7.
- 7. Chinetti G, Lestavel S, Bocher V, et al. PPAR-alpha and PPAR-gamma activators induce cholesterol removal from human macrophage foam cells through stimulation of the ABCA1 pathway.Nat Med 2001; 7: 53-8.
- 8. Rubins HB, Robins SJ, Collins D, et al. Gemfibrozil for the secondary prevention of coronary heart disease in men with low levels of high-density lipoprotein cholesterol. Veterans affairs high-density lipoprotein cholesterol intervention trial study group.N Engl J Med 1999; 341: 410-8.
- 9. DAIS-group. Effect of fenofibrate on progression of coronary-artery disease in type 2 diabetes: the diabetes atherosclerosis intervention study, a randomised study.Lancet 2001; 357: 905-10.
- 10. BIP group. Secondary prevention by raising HDL cholesterol and reducing triglycerides in patients with coronary artery disease: the Bezafibrate Infarction Prevention (BIP) study.Circulation 2000; 102: 21-7.
- 11. Tenkanen L, Manttari M, Manninen V. Some coronary risk factors related to the insulin resistance syndrome and treatment with gemfibrozil. Experience from the Helsinki Heart Study.Circulation 1995; 92: 1779-85.
- 12. Barroso I, Gurnell M, Crowley VE, et al. Dominant negative mutations in human PPARgamma associated with severe insulin resistance, diabetes mellitus and hypertension.Nature 1999; 402: 880-3.
- 13. Arner P. The adipocyte in insulin resistance: key molecules and the impact of the thiazolidinediones.Trends Endocrinol Metab 2003; 14: 137-45.
- 14. Guan HP, Li Y, Jensen MV, Newgard CB, Steppan CM, Lazar MA. A futile metabolic cycle activated in adipocytes by antidiabetic agents. Nat Med 2002; 8: 1122-8.
- 15. Kahn CR, Chen L, Cohen SE. Unraveling the mechanism of action of thiazolidinediones.J Clin Invest 2000; 106: 1305-7.
- 16. Kopp HP, Kopp CW, Festa A, et al. Impact of weight loss on inflammatory proteins and their association with the insulin resistance syndrome in morbidly obese patients.Arterioscler Thromb Vasc Biol 2003; 23: 1042-7.
- 17. Holst D, Grimaldi PA. New factors in the regulation of adipose differentiation and metabolism.Curr Opin Lipidol 2002; 13: 241-5.
- 18. Yang WS, Jeng CY, Wu TJ, et al. Synthetic peroxisome proliferator-activated receptor-gamma agonist, rosiglitazone, increases plasma levels of adiponectin in type 2 diabetic patients.Diabetes Care 2002; 25: 376-80.
- 19. Camp HS, Li O, Wise SC, et al. Differential activation of peroxisome proliferator-activated receptor- gamma by troglitazone and rosiglitazone.Diabetes 2000; 49: 539-47.
- 20. Olefsky JM. Treatment of insulin resistance with peroxisome proliferator-activated receptor gamma agonists.J Clin Invest 2000; 106: 467-72.
- 21. Mukherjee R, Hoener PA, Jow L, et al. A selective peroxisome proliferator-activated receptor-gamma (PPARgamma) modulator blocks adipocyte differentiation but stimulates glucose uptake in 3T3-L1 adipocytes.Mol Endocrinol 2000; 14: 1425-33.
- 22. Oberfield JL, Collins JL, Holmes CP, et al. A peroxisome proliferator-activated receptor gamma ligand inhibits adipocyte differentiation.Proc Natl Acad Sci USA 1999; 96: 6102-6.
- 23. Rieusset J, Touri F, Michalik L, et al. A new selective peroxisome proliferator-activated receptor gamma antagonist with antiobesity and antidiabetic activity.Mol Endocrinol 2002; 16: 2628-44.
- 24. Lohray BB, Lohray VB, Bajji AC, et al. (-)3-[4-[2-(Phenoxazin-10-yl)ethoxy]phenyl]-2-ethoxypropanoic acid [(- )DRF 2725]: a dual PPAR agonist with potent antihyperglycemic and lipid modulating activity.J Med Chem 2001; 44: 2675-8.
- 25. Etgen GJ, Oldham BA, Johnson WT, et al. A tailored therapy for the metabolic syndrome: the dual peroxisome proliferator-activated receptor-alpha/gamma agonist LY465608 ameliorates insulin resistance and diabetic hyperglycemia while improving cardiovascular risk factors in preclinical models.Diabetes 2002; 51: 1083-7.
- 26. Vikramadithyan RK, Hiriyan J, Gershome C, Rajagopalan R, Chahrabarti R. Biochemical effects of the dual PPARalpha and gamma agonist ragaglitazar on glucose and lipid metabolism. San Francisco: American Diabetes Association, 2002: abstract 584-P.
- 27. Ide T, Nakazawa T, Mochizuki T, Murakami K. Tissue-specific actions of antidiabetic thiazolidinediones on the reduced fatty acid oxidation in skeletal muscle and liver of Zucker diabetic fatty rats.Metabolism 2000; 49: 521-5.
- 28. Shibata T, Takeuchi S, Yokota S, Kakimoto K, Yonemori F, Wakitani K. Effects of peroxisome proliferator-activated receptor-alpha and -gamma agonist, JTT-501, on diabetic complications in Zucker diabetic fatty rats.Br J Pharmacol 2000; 130: 495-504.
- 29. Ye JM, Iglesias MA, Watson DG, et al. PPARalpha/gamma ragaglitazar eliminates fatty liver and enhances insulin action in fat-fed rats in the absence of hepatomegaly.Am J Physiol Endocrinol Metab 2003; 284: E531-40.
- 30. Murakami K, Tobe K, Ide T, et al. A novel insulin sensitizer acts as a coligand for peroxisome proliferator-activated receptor-alpha (PPAR-alpha) and PPAR-gamma: effect of PPAR-alpha activation on abnormal lipid metabolism in liver of Zucker fatty rats.Diabetes 1998; 47: 1841-7.
List of figures
Figure 1
Modulateurs sélectifs de PPAR (selective PPAR modulator ou SPPARM).
Chaque agoniste de PPAR (peroxisome proliferator-activated receptors), ligand A ou ligand B, peut entraîner une conformation tridimensionnelle différente du complexe PPAR-ligand. Ces conformations différentes, après liaison des hétérodimères avec le récepteur nucléaire RXR (9-cis retinoic acid receptor) et fixation sur des séquences spécifiques PPRE (peroxisome proliferator response elements), peuvent amener à un recrutement de cofacteurs différents et, par conséquent, entraîner une régulation transcriptionnelle particulière pour chaque complexe. Ainsi, les différents ligands peuvent, via un même récepteur nucléaire, induire des réponses biologiques communes, mais aussi des réponses biologiques qui leur sont spécifiques.
Figure 2
Co-agonistes PPARα/γ.
Les effets liés à l’activation des PPAR peuvent être complémentaires et synergiques. Ainsi, une nouvelle famille de molécules capables d’activer parallèlement PPARα et PPARγ a été développée. Elle peut entraîner à la fois des effets bénéfiques sur le métabolisme des lipides, via l’activation de PPARα et aboutir à l’amélioration de la sensibilité à l’insuline et de l’utilisation du glucose, via l’activation de PPARγ.
List of tables
Tableau I
Ligands de PPARγ et co-agonistes PPARα/γ en développement.