Article body
Le protéasome 26S est un complexe comprenant plus de 30 sous-unités différentes, chargé de dégrader les protéines [1]. Il est composé, chez les eucaryotes, d’une unité centrale 20S en forme cylindrique et de deux unités 19S (une à chaque extrémité). Le complexe 19S est responsable de la reconnaissance, de la dénaturation et du transfert du substrat à l’intérieur du complexe 20S où réside l’activité protéolytique (Figure 1). La reconnaissance du substrat requiert la présence d’une chaîne polyubiquitine attachée de manière covalente au substrat. L’ubiquitine est une petite protéine de 76 acides aminés de structure globulaire. La réaction d’ubiquitinylation est indépendante du protéasome [2]. Par cette réaction, l’extrémité carboxy-terminale d’une première ubiquitine se forme à partir une liaison isopeptidique avec une lysine du substrat, puis une deuxième ubiquitine est attachée à la première ubiquitine et ainsi de suite pour former une chaîne d’environ 4 à 8 molécules reliées entre elles. On a longtemps cru que, pour être transféré à l’intérieur du complexe 20S, le substrat devait se débarrasser de cette chaîne polyubiquitinylée. En effet, le diamètre intérieur du complexe 20S est seulement de 13 Å tandis que le substrat et sa chaîne d‘ubiquitines forme une structure de 25 à 30 Å. Des protéases capables de cliver les molécules d’ubiquitine du substrat (désubiquitinyler) ont depuis longtemps été identifiées. Ces isopeptidases, aussi dénommées DUB (deubiquitinating enzymes), possèdent un domaine catalytique comprenant une cystéine et deux histidines. Malgré une analyse détaillée de plusieurs de ces DUB, aucune n’a pu, jusqu’à présent, être impliquée dans la désubiquitinylation des substrats au sein du protéasome.
Figure 1
Représentation schématique de l’organisation du protéasome.
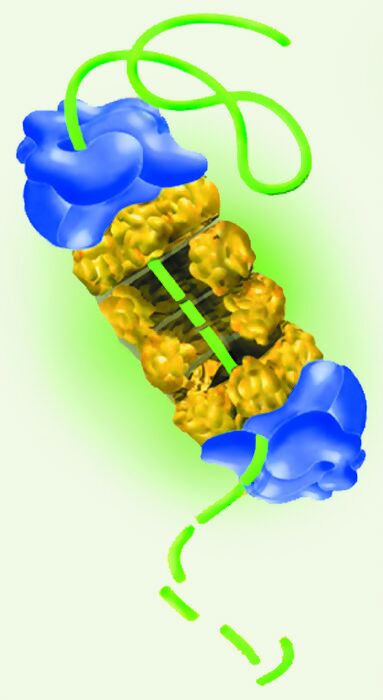
En bleu, les deux sous-unités 19S; en jaune, le complexe central 20S. La protéine intacte est figurée par un trait vert continu, et la protéine dégradée par une trait vert discontinu. (avec l’autorisation de l’US Department of Energy Human Genome Program or US Department of Energy Genomes to Life Program et tiré du site http://www.ornl.gov/hgmis).
Le protéasome: mais quelle DUB?
R. Verma et al. ont reconstitué un système de dégradation protéique in vitro en utilisant des protéasomes 26S purifiés à partir de la levure Saccharomyces cerivisiae [3]. Sic1, une protéine impliquée dans la régulation de CDK1 (cyclin dependent kinase 1), est ubiquitinylée et rapidement dégradée lors de la progression du cycle cellulaire. Comme le montrent R. Verma et al, il est possible de reconstituer ce processus de dégradation en incubant Sic1 préalablement ubiquitinylé (Ub-Sic1) avec les protéasomes purifiés. Lorsque les auteurs bloquent l’activité protéolytique du protéasome par l’inhibiteur epoxomicine, qui agit en s’associant aux sous-unités β du complexe 20S, Ub-Sic1 est désubiquitinylée mais n’est pas dégradée [3]. Les activités de désubiquitinylation et de dégradation du protéasome ont ainsi été dissociées. Les auteurs ont ensuite cherché à bloquer l’activité de désubiquitinylation en traitant leurs protéasomes purifiés avec des inhibiteurs traditionnels de DUB comme l’ubiquitine aldéhyde. Étonnamment, ils n’y sont pas parvenus, ce qui indique que l’activité de désubiquitinylation du protéasome ne semble pas dépendre d’une DUB classique. Or une telle DUB, Ubp6, existe bien, comme le révèle l’analyse par spectrométrie de masse de protéasomes purifiés. Cependant, sa suppression ne semble pas empêcher l’activité de désubiquitinylation du protéasome [3], et le mystère restait donc entier quant à la source de l’activité de désubiquitinylation. Les auteurs auraient pu en rester là, s’ils n’avaient pas mis à profit une étude menée sur Nedd8, une autre protéine similaire à l’ubiquitine.
Nedd8, le cousin bien utile
Nedd8 est un homologue de l’ubiquitine et forme aussi une liaison covalente avec son substrat, mais seulement de manière monomérique. Une précédente étude avait montré que le « signalosome » (CSN) est capable de promouvoir le clivage de Nedd8 de son substrat [4]. CSN est un complexe composé de 8 protéines. Il a été impliqué dans différents processus comme le développement, la régulation de la transcription ou l’activation de certaines kinases, sans qu’on ait pu toutefois déterminer sa fonction précise. Dans l’espoir d’identifier la protéine responsable du clivage de Nedd8, G.A. Cope et al. ont analysé les séquences des différents membres du CSN. Ils ont ainsi pu mettre en évidence un domaine présent dans Cns5 et tous ses orthologues [5]. Ce domaine, JAMM (Jab1/MPN domain metalloenzyme) est constitué d’un résidu Glu en amont et du motif His-X-His-X10-Asp qu’on trouve dans certains sites d’hydrolases se liant à un ion métallique. Les auteurs ont émis l’hypothèse selon laquelle JAMM pouvait former un site catalytique de type métalloprotéase capable de cliver Nedd8. Pari gagné, puisque l’ajout de 1,10 phénanthroline, un chélateur d’ions métalliques ayant une forte affinité pour le zinc, inhibe le clivage de Nedd8 de son substrat [5]. Une inhibition identique est observée lorsque les résidus conservés du domaine JAMM de Csn5 sont mutés [5]. G.A. Cope et al. proposent que le résidu Glu se lie à une molécule d’H2O tandis que les trois autres résidus (2xHis et Asn) stabilisent l’ion métallique au sein du site catalytique (Figure 2A). Csn5 ferait ainsi partie d’une nouvelle classe de protéases, différentes des DUB traditionnelles, capables de cliver des protéines similaires à l’ubiquitine.
Figure 2
Le motif JAMM de Csn5 et Rpn11.
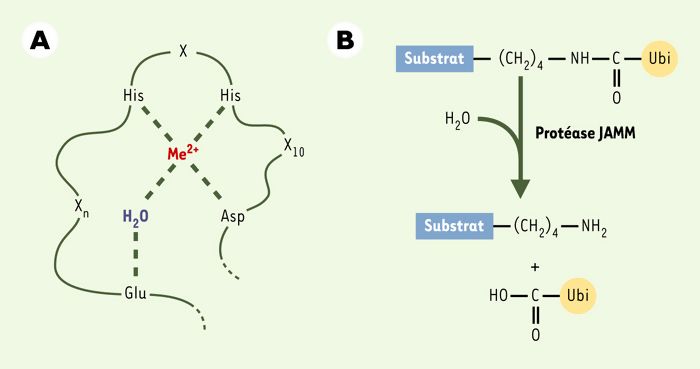
A. Représentation de la coordination d’un ion zinc (Me2+) et d’une molécule H2O au sein du site catalytique JAMM. B. Clivage de la liaison isopeptidique entre le substrat et Nedd8 ou ubiquitine (Ubi) par une protéase JAMM.
Retour à la désubiquitinylation
La structure du signalosome partage des similitudes avec la partie supérieure 19S du protéasome. En effet, on retrouve des domaines conservés entre plusieurs sous-unités de ces deux complexes.
Rpn11, une sous-unité de 19S, est une protéine homologue de Csn5 contenant aussi un domaine JAMM. On pouvait donc imaginer que ce domaine pouvait être responsable de l’activité de désubiquitinylation du protéasome. Pour le démontrer, R. Verma et al. ont muté le motif JAMM de Rpn11 ou introduit un chélateur d’ions métalliques dans leurs expériences. Cette fois-ci, l’activité de désubiquitinylation du protéasome est inhibée [3]. De plus, une souche de levure n’exprimant que la forme mutée de Rpn11 n’est pas viable, indiquant le rôle essentiel du domaine JAMM. En effet, le protéasome ne peut dégrader un substrat que si la chaîne polyubiquitine est enlevée [3]. Dans une étude indépendante, Yao et Cohen sont arrivés aux mêmes conclusions en utilisant des protéasomes purifiés à partir de tissus animaux et en mutant différents résidus du motif JAMM de Rpn11 chez la levure [6]. Ces deux études montrent donc l’importance du motif JAMM de Rpn11 dans la désubiquitinylation de substrats par le protéasome.
Perspectives: le protéasome est ouvert!
Ces études récentes révèlent la présence d’une nouvelle classe de métalloprotéases impliquées dans le clivage de chaînes polyubiquitinylées ou de Nedd8 (Figure 2B). Des analyses structurales supplémentaires sont encore nécessaires pour démontrer que le centre catalytique réside bien dans le motif JAMM et requiert la coordination d’un ion métallique comme le zinc. Comme les auteurs l’ont noté, d’autres protéines contiennent le motif JAMM, et il sera intéressant de déterminer si ces dernières possèdent la même activité isopeptidique, et d’identifier leurs substrats.
La présence du complexe 19S au sein du protéasome a souvent posé problème puisqu’il n’existe pas chez les bactéries et les archéobactéries. Cependant, ces derniers organismes ne possèdent pas d’ubiquitine et l’entrée des substrats est ainsi simplifiée. Les eucaryotes ont développé un système plus complexe. Il est intéressant de noter que les deux équipes ont trouvé que la désubiquitinylation requiert la présence d’ATP [3, 6]. La base de 19S comprend, en effet, plusieurs ATPases, dont la fonction présumée est de dénaturer le substrat puis de le transférer à l’intérieur du complexe 20S. Il n’est pas surprenant de constater que la désubiquitinylation est couplée à ce processus, et en dépend. Ce dernier exemple illustre la complexité de la machinerie du protéasome, et d’autres études seront encore nécessaires pour compléter ce puzzle.
Appendices
Remerciements
Je remercie Julien Weber et Gabriela Alexandru pour leurs corrections et commentaires.
Références
- 1. Voges D, Zwickl P, Baumeister W. The 26S proteasome: a molecular machine designed for controlled proteolysis. Annu Rev Biochem 1999; 68: 1015-68.
- 2. Weissman AM. Themes and variations on ubiquitinylation. Nat Rev Mol Cell Biol 2001; 2: 169-78.
- 3. Verma R, Aravind L, Oania R, et al. Role of Rpn11 metalloprotease in deubiquitination and degradation by the 26S proteasome. Science 2002; 298: 611-5.
- 4. Lyapina S, Cope G, Shevchenko A, et al. Promotion of NEDD-CUL1 conjugate cleavage by COP9 signalosome. Science 2001; 292: 1382-5.
- 5. Cope GA, Suh GS, Aravind L, et al. Role of predicted metalloprotease motif of Jab1/Csn5 in cleavage of Nedd8 from Cul1. Science 2002; 298: 608-11.
- 6. Yao T, Cohen RE. A cryptic protease couples deubiquitination and degradation by the proteasome. Nature 2002; 419: 403-7.
List of figures
Figure 1
Représentation schématique de l’organisation du protéasome.
En bleu, les deux sous-unités 19S; en jaune, le complexe central 20S. La protéine intacte est figurée par un trait vert continu, et la protéine dégradée par une trait vert discontinu. (avec l’autorisation de l’US Department of Energy Human Genome Program or US Department of Energy Genomes to Life Program et tiré du site http://www.ornl.gov/hgmis).
Figure 2
Le motif JAMM de Csn5 et Rpn11.
A. Représentation de la coordination d’un ion zinc (Me2+) et d’une molécule H2O au sein du site catalytique JAMM. B. Clivage de la liaison isopeptidique entre le substrat et Nedd8 ou ubiquitine (Ubi) par une protéase JAMM.