Abstracts
Résumé
Pendant longtemps, la toxicomanie a été associée sur le plan neurobiologique à la modulation à court terme de différents systèmes de neurotransmission. Les stratégies de traitement ciblaient conséquemment les récepteurs auxquels se lie directement la substance étant source d’abus. Ces approches ont contribué à améliorer le soulagement des symptômes d’intoxication et de sevrage, tout en favorisant l’accès à des services psychosociaux adaptés. Toutefois, les données soulignent, chez certains sous-groupes d’individus, l’efficacité parfois mitigée de ces interventions visant à diminuer de façon soutenue la consommation et les symptômes associés à la toxicomanie, particulièrement le craving. Les avancées récentes en neurosciences ont permis de mieux comprendre les mécanismes neurobiologiques expliquant la vulnérabilité à la rechute. D’une conception essentiellement dopaminergique et striatale, les théories biologiques de la toxicomanie intègrent maintenant la contribution des systèmes glutamatergique, opioïde et endocannabinoïde, de même que l’interaction entre ces différentes composantes au sein des structures corticales et sous-corticales. L’intérêt semble avoir migré des phénomènes neurobiologiques à court terme vers la modulation prolongée du fonctionnement des structures en jeu dans la toxicomanie. Ce changement de paradigmes a mené à l’émergence de plusieurs stratégies thérapeutiques visant à diminuer les risques de rechute en modulant de façon plus spécifique les circuits neuronaux dont le fonctionnement est altéré par la prise chronique de substances. Les systèmes endocannabinoïde et glutamatergique, notamment, apparaissent comme une cible de choix pour le traitement du craving et la prévention de la rechute. Le présent article a pour objectif de résumer certains des plus récents courants en matière de conceptualisation neurobiologique de la toxicomanie de même que les nouvelles pistes de traitement en découlant.
Mots-clés :
- Toxicomanie,
- craving,
- système endocannabinoïde,
- rechute,
- neurobiologie de la toxicomanie,
- glutamate
Abstract
For years, the neurobiology of drug addiction was characterized by the short-term modulation of different neurotransmission systems, therapeutic strategies directly targeting the receptors that are bound by substances. These approaches have helped to improve the treatment of drug intoxication and withdrawal, while promoting access to a broad array of psychosocial services. However, the data highlight the mixed effectiveness of these interventions to induce a sustained decrease in consumption and other symptoms of addiction, especially craving, among subgroups of individuals. Recent advances in neuroscience have led to a growing understanding of the neurobiological mechanisms underlying the vulnerability to relapse and other behaviors associated with addiction. The primarily dopaminergic and striatal hypothesis of these phenomena has been replaced by a theory incorporating the contribution of the glutamatergic, endocannabinoid and opioid systems, as well as the interaction between these various components within cortical and sub-cortical structures. The focus has moved from the short-term neurobiological changes to the long-lasting modulation of the structures involved in addiction. This paradigm shift led to the emergence of several therapeutic strategies that aim at reducing the risk of relapse by modulating specific neural circuits whose functions are altered by chronic substance use. The endocannabinoid and glutamate systems, in particular, are promising targets for the treatment of craving and relapse. This article aims to summarize some of the latest trends in the neurobiology of addiction as well as new avenues of treatment.
Keywords:
- Addiction,
- craving,
- endocannabinoid system,
- relapse,
- neurobiology of addiction,
- glutamate
Resumen
Durante mucho tiempo la toxicomanía estuvo asociada en el plano neurobiológico con la modulación a corto plazo de diferentes sistemas de neurotransmisión. Las estrategias de tratamiento apuntaban, por consiguiente, a los receptores que se vinculan directamente con la sustancia que es el origen del abuso. Estos enfoques contribuyeron a mejorar el alivio de los síntomas de intoxicación y abstinencia, favoreciendo al mismo tiempo el acceso a servicios psicosociales adaptados. Sin embargo, en ciertos grupos de individuos, los datos destacan la eficacia a veces mitigada de estas intervenciones destinadas a disminuir de manera continua el consumo y los síntomas relacionados con la toxicomanía, particularmente el craving. Los recientes progresos de la neurociencia permitieron comprender mejor los mecanismos neurobiológicos que explican la vulnerabilidad ante la recaída. De concepción esencialmente dopaminérgica y estratial, las teorías biológicas de la toxicomanía integran ahora la contribución de los sistemas glutamatérgico, opiáceo y endocanabinoide, así como la interacción entre estos diferentes componentes dentro de las estructuras corticales y subcorticales. El interés parece haberse desplazado de los fenómenos neurobiológicos de corto plazo hacia la modulación prolongada del funcionamiento de las estructuras que están en juego en la toxicomanía. Este cambio de paradigmas condujo al surgimiento de numerosas estratégicas terapéuticas destinadas a disminuir los riesgos de recaída al modular más específicamente los circuitos neuronales cuyo funcionamiento está alterado por el consumo crónico de drogas. Los sistemas endocanabinoide y glutamatérgico, principalmente, aparecen como un objetivo a privilegiar para el tratamiento del craving y la prevención de la recaída. El presente artículo está destinado a resumir algunas de las más recientes corrientes en materia de conceptualización neurobiológica de la toxicomanía y las nuevas vías de tratamiento a las que dichas corrientes dan origen.
Palabras clave:
- Toxicomanía,
- craving,
- sistema endocanabinoide,
- recaída,
- neurobiología de la toxicomanía,
- glutamato
Article body
Introduction
Malgré l’ampleur des problèmes liés à la consommation de substances licites et illicites (UNODC, 2008), les processus neurobiologiques en cause dans le phénomène de la toxicomanie ne sont encore que partiellement compris. La neuroimagerie et le développement récent des technologies liées à la génétique, entre autres, ont toutefois permis des progrès majeurs dans l’étude des neurosciences. D’une conception essentiellement dopaminergique et striatale, les théories biologiques de la toxicomanie intègrent maintenant la contribution des systèmes glutamatergique, opioïde et endocannabinoïde, de même que l’interaction entre ces différentes composantes au sein du striatum et des structures mésocorticolimbiques. Ce changement de paradigme a mené à l’émergence de plusieurs stratégies thérapeutiques visant la réduction du risque de rechute en modulant de façon plus spécifique le fonctionnement de ces structures corticales et sous-corticales.
Nous tenterons ici de décrire les stratégies pharmacologiques actuellement utilisées dans le traitement médical de la toxicomanie. Nous soulignerons les domaines où ces traitements se sont avérés efficaces, mais aussi leurs lacunes et faiblesses. L’illustration des mécanismes neurobiologiques sous-jacents à la rechute apparaît comme une méthode intéressante pour explorer les avenues thérapeutiques possibles, tout en reconnaissant les limites des connaissances actuelles. Nous nous attarderons à la vulnérabilité des utilisateurs de drogues face à la rechute, au cours de la période suivant l’arrêt de la consommation. Cette période post-sevrage est maintenant identifiée comme une phase importante du cycle de la toxicomanie. Bien que les données présentées ici s’inscrivent dans le courant médical du traitement de la toxicomanie, il s’agit de la description d’un modèle intégrant d’autres conceptualisations existantes. Loin d’alimenter l’historique dichotomie entre les processus psychologiques, environnementaux et neurobiologiques en cause dans le phénomène de la dépendance aux drogues, nous soulignerons l’importance relative de chacune de ces composantes et leur interaction, faisant ainsi foi d’un processus complexe, plutôt que de composantes parallèles et simplifiées.
Traitements actuels
L’effet des drogues est en grande partie expliqué par leur capacité à passer la barrière hémato-encéphalique et à se lier à différents récepteurs neuronaux, entraînant une cascade de modifications de la neurotransmission cérébrale et du fonctionnement cellulaire. À court terme, cette propriété peut expliquer les symptômes d’intoxication lorsque la substance est consommée en quantité importante. Par exemple, l’héroïne et les autres opioïdes se lient aux récepteurs opioïdes de type mu, kappa et delta au sein du cortex, du système limbique et des ganglions de la base pour entraîner analgésie, sédation et euphorie. Le delta-9-tétrahydrocannabinol (THC), la substance psychoactive contenue dans le cannabis, se lie aux récepteurs cannabinoïdes de type (CB1) dans le cortex, le système limbique, le cervelet et les ganglions de la base pour provoquer désinhibition, analgésie et euphorie. Les amphétamines agissent sur la neurotransmission noradrénergique et augmentent la transmission dopaminergique par l’inhibition de la recapture de la dopamine dans la fente synaptique de même que par une augmentation de la libération de dopamine, causant ainsi de l’euphorie, une diminution de la fatigue et une augmentation de la vigilance et de la tension artérielle (Feltenstein et See, 2008).
Dans le contexte d’une prise de drogue de façon répétitive, il peut apparaitre une consommation dite pathologique. Selon le Manuel diagnostique et statistique des troubles mentaux (APA, 2000), un diagnostic d’abus est différencié d’une utilisation non pathologique s’il y a la présence d’un mode répétitif de consommation menant à des problèmes sur les plans social, interpersonnel, physique ou légal. Quant à la dépendance, elle est caractérisée non seulement par des conséquences négatives sur le fonctionnement social et la santé, mais aussi par des changements physiologiques (tolérance et sevrage) et des comportements témoignant d’une perte de contrôle sur l’utilisation de la drogue. La toxicomanie est ainsi caractérisée par l’utilisation compulsive et non contrôlée de substances ainsi que par des épisodes répétés d’intoxications, d’arrêts plus ou moins prolongés de consommation et de rechutes. Des interventions adaptées sont nécessaires pour chacune de ces conditions (Figure 1). Les épisodes d’intoxication sont en général traités efficacement par un soutien médical standard (sécurisation des voies respiratoires, surveillance des fonctions cardiaque et hépatique, hémodialyse) (Kleber et coll., 2007). Dans les cas les plus sévères, des interventions pharmacologiques spécifiques peuvent être nécessaires, les molécules utilisées entrant en compétition avec la drogue pour se lier aux récepteurs par lesquels s’exercent les effets toxiques de la substance (effet antagoniste). La naloxone, un antagoniste des récepteurs opioïdes qui renverse les symptômes d’intoxication à l’héroïne ou aux opiacés (Galanter et Kleber, 2008), et le flumazenil, un antagoniste puissant des récepteurs GABA utilisé pour l’intoxication aux benzodiazépines (Kleber et coll., 2007), sont les deux antagonistes les plus largement utilisés en situation d’urgence. Après quelques heures d’abstinence peut se développer un syndrome de sevrage, caractérisé par des symptômes physiques et psychologiques souvent intolérables qui contribuent à la difficulté de rester abstinent à court terme. Dans les cas plus sévères, les symptômes peuvent être soulagés par la prise d’une médication qui remplace l’effet pharmacodynamique de la substance sevrée. Deux stratégies pharmacologiques souvent adoptées pour le soulagement des symptômes de sevrage sont les benzodiazépines pour le sevrage à l’alcool et la méthadone pour le syndrome de retrait aux opioïdes, en particulier à l’héroïne (Amato et coll., 2005 ; Kleber et coll., 2007 ; Ntais et coll., 2005). Tant les services internes qu’externes sont couramment utilisés pour le traitement des cas graves de sevrage aigu nécessitant une désintoxication. Bien qu’étant nécessaire chez certains patients présentant des risques de complications médicales, la désintoxication est rarement suffisante en soi pour permettre une rémission prolongée ou une diminution soutenue de la consommation (Mattick et Hall, 1996 ; Gossop et coll., 2002).
Figure 1
Différentes phases du cycle de la toxicomanie et quelques exemples d’interventions associées

Lorsque l’utilisateur de drogues ou d’alcool parvient à maintenir son abstinence pendant une période prolongée, le sevrage aigu fait place à une longue période de symptômes subaigus, caractérisée par un sevrage à bas bruit, une adaptation difficile à une vie sans substance et un réseau social appauvri, ainsi qu’à des épisodes récurrents de désir intense de consommer et d’intolérance au stress. Ces facteurs augmentent la susceptibilité à la rechute et la période de sevrage subaigu apparaît donc comme un moment où les interventions thérapeutiques peuvent avoir un impact significatif sur l’évolution à long terme de la toxicomanie.
Stimulation directe des récepteurs : agonisme
Une des stratégies thérapeutiques couramment utilisées dans le traitement de la toxicomanie consiste à stimuler directement les récepteurs ciblés par la substance qui est source de dépendance, et d’administrer cette médication de façon plus ou moins prolongée (traitement de substitution). Sur le plan théorique, cette stratégie permet de soulager les symptômes de sevrage et de bloquer l’effet pharmacologique des drogues d’abus, tout en contrôlant le mode d’administration et le dosage de la médication utilisée. De plus, ce type de traitement implique pour le patient des contacts réguliers avec les services de santé, qui peuvent mettre en oeuvre des stratégies thérapeutiques complémentaires, notamment sur le plan psychosocial, et un suivi des comorbidités physiques et psychologiques.
Utilisée couramment depuis plus de 40 ans pour le traitement de la dépendance aux opioïdes, la méthadone est un agoniste complet des récepteurs opioïdes de type mu (ROM) qui sont ciblés par les opioïdes comme l’héroïne (Dole et Nyswander, 1965). La demi-vie de la méthadone (environ 24 heures) et son absorption gastro-intestinale permet un dosage quotidien par voie orale (Garrido et Troconiz, 1999 ; Eap et coll., 2002). En se liant aux ROM, la méthadone soulage les symptômes de sevrage (Amato et coll., 2005). Plusieurs études ont aussi démontré que le traitement de substitution à long terme avec la méthadone diminue le risque d’infections transmissibles par le sang (par ex. : le virus de l’immunodéficience humaine-VIH) lié à la consommation de drogues intraveineuses (Lott et coll., 2006 ; Millson et coll., 2007). Ce traitement est associé à une diminution de la consommation (Mattick et coll., 2003), de la criminalité (Johansson et coll., 2007) et de la mortalité (Gibson et coll., 2008). Malheureusement, le suivi avec les services de santé est souvent conditionnel à la prescription de méthadone. Or, même quand les modes de prescriptions de méthadone sont optimisés (dosage élevé et flexible), 40 % des patients maintenus sous méthadone abandonnent leur traitement de substitution dans la première année de suivi, le taux d’abandons dépassant 50 % lorsque des dosages sous-optimaux sont utilisés (Bao et coll., 2009). Parmi les hypothèses soulevées pour expliquer ces taux de rétention relativement faibles, notons que les individus dépendants à l’héroïne continuent sous méthadone d’éprouver un désir intense de consommer (aussi appelé craving) lorsqu’ils sont exposés à des stimuli liés à la consommation ou au stress (Ilgen et coll., 2008 ; Langleben et coll., 2008). Des interventions qui ciblent ces symptômes résiduels (recherche de drogue, désir de consommer) pourraient permettre d’optimiser l’efficacité des programmes de maintien sous méthadone. Par ailleurs, la méthadone est associée à des effets secondaires parfois intolérables (constipation, dysphorie) et plus rarement létaux (dépression respiratoire, arythmie cardiaque). L’index thérapeutique de la méthadone est étroit, et les risques de surdoses mortelles sont particulièrement élevés chez les usagers naïfs (Eap et coll., 2002). Ceci est particulièrement inquiétant, surtout que la diversion de la méthadone à des fins d’usage illicite est un problème récurrent dans plusieurs groupes d’utilisateurs (Roche et coll., 2008 ; Hopwood et coll., 2003 ; Waldvogel et coll., 2005).
La buprénorphine, un agoniste partiel des récepteurs opioïdes de type mu qui agit également sur les récepteurs kappa, delta et ORL-1 (opiate receptor-like 1) (Lutfy et Cowan, 2004), semblait pouvoir répondre, du moins sur le plan théorique, aux dangers et limites inhérents à la prescription de méthadone. En effet, les risques de surdosage et de diversion sont théoriquement moindres, particulièrement dans le cas de la Suboxone, une formulation combinant la buprénorphine avec de la naloxone, laquelle vient bloquer la stimulation des récepteurs opioïdes en cas de prise parentérale ou de rechute à l’héroïne (Walsh et Eissenberg, 2003 ; Simojoki et coll., 2008). La longue demi-vie de la buprénorphine (Elkader et Sproule, 2005) permet son administration trois fois par semaine, ce qui peut faciliter l’adhérence au traitement. Ces caractéristiques font de la buprénorphine une stratégie prometteuse du traitement de la dépendance aux opioïdes. Des études récentes soutiennent d’ailleurs l’efficacité de la buprénorphine, l’associant à une diminution de la prise d’opioïdes (Connock et coll., 2007 ; Magura et coll., 2007). Toutefois, cette médication possède une puissance pharmacologique limitée par rapport à la méthadone et n’est pas supérieure quant à son efficacité pour diminuer le risque de rechute et assurer la rétention en traitement (Mattick et coll., 2008). Parmi les limites que pose l’utilisation de la buprénorphine dans le traitement de la dépendance aux opioïdes, notons que son administration est problématique chez les patients dépendants qui nécessitent une analgésie avec un narcotique et le risque de diversion s’est avéré significatif dans certaines études (Winstock et coll., 2008).
L’utilisation de morphine à libération lente s’est aussi avérée efficace et bien tolérée pour le traitement de la dépendance aux opioïdes (Eder et coll., 2005 ; Kraigher et coll., 2005 ; Mitchell et coll., 2004). Cette approche semble particulièrement utile comme alternative thérapeutique pour traiter les patients qui présentent des effets secondaires ou des symptômes résiduels de sevrage malgré la prise de méthadone (Kastelic et coll., 2008). Par ailleurs, le traitement de substitution dans les programmes utilisant l’héroïne est offert dans plusieurs pays et continue d’être une source de controverse (Gartry et coll., 2009), malgré les résultats généralement positifs des études randomisées et observationnelles (Dijkgraaf et coll., 2005 ; Haasen et coll., 2007 ; Symington, 2008 ; van den Brink et coll., 2003). Ces données suggèrent que cette approche pourrait être utile pour un sous-groupe de patients qui ne répondent pas aux traitements conventionnels.
La thérapie de remplacement de la nicotine (TRN) est couramment utilisée afin de traiter la dépendance à la nicotine et s’est avérée efficace pour traiter les symptômes de sevrage et augmenter le taux d’arrêt du tabagisme. L’effet de la TRN est toutefois mitigé sur le craving et l’effet hédonique de la cigarette (Shiffman et coll., 2006 ; Waters et coll., 2004), et les taux de rechute demeurent à près de 80 % après 12 mois de traitement (Stead et coll., 2008). Quant à la varénicline, elle s’avère une alternative valable à la TRN, mais est associée à des effets secondaires importants et à des taux de rechute significatifs (Cahill et coll., 2008). Les tentatives de développement d’une stratégie similaire pour la dépendance à la cocaïne à l’aide de stimulants sont décevantes dans l’ensemble et les études rapportent des taux de rechute élevés et une persistance de la consommation (Castells et coll., 2007). Les données sur la thérapie de remplacement pour la dépendance aux amphétamines et au cannabis demeurent limitées, quoique certains résultats encourageants aient été rapportés (Elkashef et coll., 2008 ; Tiihonen et coll., 2007 ; Levin et Kleber, 2008).
Blocage de la stimulation des récepteurs : antagonisme
Une alternative thérapeutique consiste à administrer une substance antagoniste, qui bloque les récepteurs ciblés par la drogue d’abus sans les activer, et par conséquent inhibe l’effet euphorisant. La naltrexone est un exemple bien connu d’antagoniste spécifique et est utilisée dans le traitement de la dépendance à l’héroïne. Malgré un intérêt théorique, le succès de cette avenue thérapeutique s’est avéré mitigé dans les essais cliniques. En effet, bien que la naltrexone puisse dans une certaine mesure diminuer la consommation d’héroïne, la rétention des sujets en traitement est problématique (Johansson et coll., 2006 ; Minozzi et coll., 2006). La naltrexone peut donc être un adjuvant thérapeutique intéressant pour des sujets hautement motivés chez qui l’abstinence totale procure des bénéfices intéressants, ce qui est notamment le cas de certains professionnels de la santé et des affaires ou d’individus sous probation (Ling et Wesson, 1984 ; O’Brien et Cornish, 2006 ; Roth et coll., 1997 ; Washton et coll., 1984). La version « dépôt » à effet prolongé pourrait partiellement répondre à la problématique de non-adhérence au traitement, mais la littérature demeure encore limitée à cet égard (Lobmaier et coll., 2008). Les individus traités avec la naltrexone demeurent aussi vulnérables au craving engendré par le stress et les stimuli associés à la drogue, ce qui pourrait être un des mécanismes à l’origine des taux élevés de non-adhérence thérapeutique (Hyman et coll., 2007).
Des études ont démontré l’efficacité de la mécamylamine, un antagoniste non compétitif des récepteurs de l’acétylcholine nicotinique, en combinaison avec la TRN pour le traitement du tabagisme. Cette médication diminue les effets renforçateurs de la nicotine alors que la TRN réduit les symptômes de sevrage (Lancaster et Stead, 2000 ; Galanter et Kleber, 2008). Des essais thérapeutiques ont aussi été menés pour identifier des molécules pouvant bloquer les effets de la cocaïne, soit en bloquant les sites de liaison au transporteur de la dopamine (c.-à-d. mazindole) ou en bloquant les récepteurs dopaminergiques (c.-à-d. ecopipam, antipsychotiques). Dans ces deux cas, les résultats se sont montrés décevants en matière de traitement de la dépendance à la cocaïne, tant pour diminuer de façon soutenue le craving que l’utilisation de cette substance (Margolin et coll., 1995 ; Stine et coll., 1995 ; Amato et coll., 2007 ; Haney et coll., 2001).
Blocage de l’effet euphorisant des substances
D’autres stratégies, définies comme aversives, ont été employées pour décourager l’individu dépendant de consommer, la molécule la plus connue dans cette catégorie étant le disulfiram. En inhibant de façon irréversible l’aldéhyde déshydrogénase, qui assure la transformation d’acétaldéhyde en acétate pouvant être éliminé par l’organisme, le disulfiram entraîne l’accumulation d’acétaldéhyde. Cette accumulation cause une pléiade de symptômes désagréables chez le patient qui consommerait de l’alcool tout en prenant du disulfiram (Galanter et Kleber, 2008). Bien que cette médication soit utile chez certains patients hautement motivés, l’adhérence au traitement est faible, nécessitant la supervision de l’administration ou entraînant tout simplement l’arrêt du traitement. Certains auteurs suggèrent que la combinaison avec d’autres traitements ciblant le craving pourrait permettre de maximiser le potentiel du disulfiram dans le traitement de l’alcoolisme (Suh et coll., 2006).
Les développements récents en immunologie, conjugués au manque de traitement pour la dépendance à certaines drogues comme la cocaïne et le PCP, ont mené au développement de stratégies ciblant l’intervention du système immunitaire contre la substance source d’abus, notamment par la fabrication d’anticorps se liant à la substance pour empêcher qu’elle ne passe la barrière hémato-encéphalique et qu’elle exerce ses effets (Orson et coll., 2008 ; Kinsey et coll., 2009). L’évaluation de cette avenue thérapeutique en est à ses débuts et il reste à prouver qu’elle s’avérera efficace pour traiter la toxicomanie.
Résumé et lacunes des traitements actuels
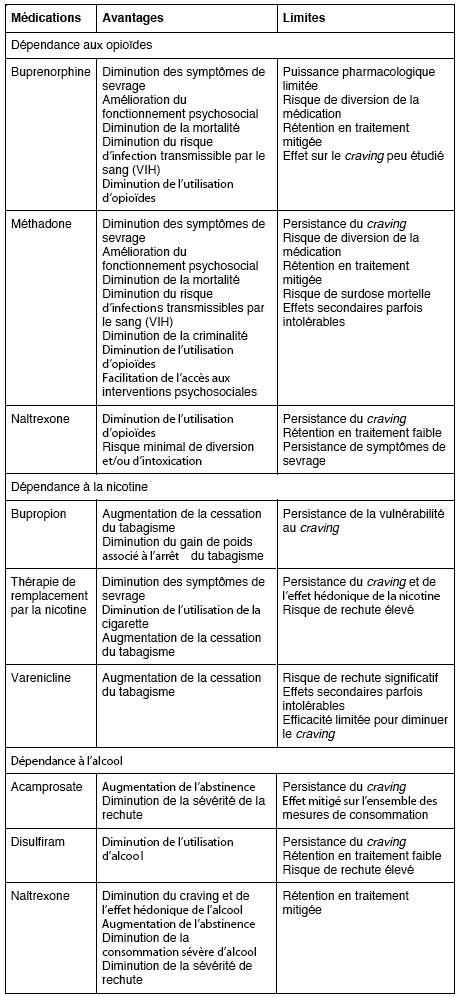
Pendant longtemps, la toxicomanie s’est résumée sur le plan neurobiologique à une sur ou sous-stimulation du fonctionnement des récepteurs auxquels se lie la substance abusée (p. ex. : opioïdes, cannabinoïdes). Il n’est donc pas étonnant de constater que les stratégies thérapeutiques se soient généralement limitées à cibler ces récepteurs. Ces approches ont contribué à soulager les symptômes de sevrage et à favoriser l’accès à des services psychosociaux adaptés. Toutefois, si pour un grand nombre d’individus les stratégies actuelles sont efficaces, les données soulignent, chez certains sous-groupes, leur efficacité mitigée à engendrer une diminution soutenue de la consommation et des symptômes associés à la toxicomanie, particulièrement le craving (Tableau 1). Il est possible que la description incomplète des phénomènes neurobiologiques en jeu puisse expliquer la difficulté que représente le traitement de certains symptômes et comportements qui sont au coeur même de la toxicomanie, comme la vulnérabilité prolongée à la rechute. Une meilleure compréhension de ces phénomènes a permis d’en dévoiler toute la complexité, tout en laissant émerger une multitude de nouvelles avenues thérapeutiques qui, bien qu’elles n’en soient qu’à leurs premiers pas, semblent fort prometteuses.
Tableau 1
Principaux avantages et limites des médications couramment utilisées dans le traitement de la dépendance aux substances (liste non exhaustive)
Évolution des neurosciences de la toxicomanie et des stratégies thérapeutiques associées
La dépendance à une substance est associée aux phénomènes de tolérance et de sevrage ainsi qu’à l’apparition de comportements nouveaux liés à la prise de drogue (APA, 2000). Ces comportements témoignent de la saillance particulière de la drogue par rapport aux stimuli naturels tels la nourriture, la sexualité et les rapports sociaux. La toxicomanie et les comportements de recherche de drogues qui suivent l’exposition répétée à une substance sont le résultat d’une interaction complexe qui survient entre des caractéristiques prémorbides et des modifications de la fonction neuronale. L’impact de cette interaction semble aller bien au-delà des modulations aiguës engendrées par l’utilisation ponctuelle d’une substance.
Dopamine et consommation compulsive de drogues
Un des facteurs communs aux substances qui peuvent engendrer une dépendance est le fait d’avoir des propriétés hédoniques. Cette caractéristique serait étroitement liée à la relâche de dopamine (DA) au sein des voies mésocorticolimbiques, lesquelles proviennent de l’aire tegmentale ventrale (ATV) et projettent au noyau accumbens (NAc), au cortex préfrontal (CPF) et à l’amygdale (Pierce et Kumaresan, 2006 ; Volkow et coll., 2009). Les propriétés hédoniques transitoires des substances ont longtemps été considérées comme l’élément central de la dépendance aux drogues. Bien qu’importante, particulièrement afin de favoriser la consommation récréative de substances, cette caractéristique peut difficilement expliquer l’ensemble des changements comportementaux associés à la toxicomanie, qui continuent d’être exprimés au-delà de l’effet transitoire des drogues. Des modifications profondes et durables du système dopaminergique semblent s’opérer, et la seule restauration des niveaux de dopamine ne semble d’ailleurs pas suffisante, tout comme les thérapies de substitution, pour pallier ces processus (Elkashef et coll., 2008 ; Poling et coll., 2006 ; Elkashef et coll., 2006).
Plusieurs études ont montré le rôle critique de la relâche de dopamine au sein du striatum et des structures mésocorticolimbiques dans l’apprentissage de nouveaux comportements (Wise, 2004). Les drogues ont les propriétés dopaminergiques nécessaires pour permettre l’acquisition de comportements qui engendrent la répétition de leur consommation, tout comme le font des stimuli tels que la nourriture et la sexualité (Schultz, 1998 ; 2001). Qui plus est, les substances provoquent une augmentation plus importante des niveaux de dopamine que ces stimuli biologiques, et contrairement à ces derniers, la libération de dopamine n’est pas soumise au phénomène d’habituation (Kalivas et O’Brien, 2008). Chaque exposition vient donc renforcer l’association entre prise de drogue et récompense, de même que consolider l’apprentissage des comportements permettant d’obtenir la substance en question. La relâche de dopamine vient également permettre d’associer certains stimuli à l’obtention de récompense initialement liée à la prise de drogue (Schultz, 1998).
Le développement accéléré des neurosciences a permis des progrès majeurs dans l’étude des mécanismes associés aux changements de comportements persistants liés à la toxicomanie. Il est maintenant établi qu’une exposition chronique à une drogue a des conséquences neurobiologiques plus complexes que la simple stimulation passagère des récepteurs dopaminergiques. Nous avons déjà mentionné que toute drogue a la capacité de se lier à des récepteurs particuliers pour avoir à court terme des effets psychologiques et physiques spécifiques. Or, l’activation de tels récepteurs serait aussi à l’origine de la modulation de différents systèmes de signalisation intracellulaire régulant l’expression des gènes. À titre d’exemple, la cascade de stimulations des récepteurs D1 provoque une augmentation de l’adénosine monophosphate cyclique (AMPc), de la protéine kinase A et de la protéine CREB (cAMP Response Element Binding protein). Cette augmentation vient influencer la transcription d’un ensemble de gènes suspectés d’avoir un rôle important dans la toxicomanie, tels que les gènes préprodynorphin, Homer et deltaFosB (Kalivas et O’Brien, 2008). DeltaFosB est la source de beaucoup d’intérêt puisque son expression est associée à une augmentation de l’effet renforçateur des drogues (Zachariou et coll., 2006 ; Kalivas et O’Brien, 2008 ; Nestler, 2008). Ce facteur de transcription est un candidat de choix pour expliquer le développement de la prise compulsive de drogues, d’autant plus que la neuroplasticité causée par d’autres gènes dont il module l’expression persisterait au-delà de sa propre dysrégulation (Norrholm et coll., 2003 ; Robinson et Kolb, 1999). L’expression de deltaFosB est modulée par plusieurs substances et pourrait donc avoir un rôle crucial dans les phénomènes neurobiologiques persistants en jeu dans la toxicomanie (Nestler, 2008). Pour l’instant, il n’y a pas d’inhibiteur de deltaFosB pouvant être administré chez l’humain, mais le développement d’une telle molécule apparaît comme particulièrement prometteur afin de cibler les processus neurobiologiques engendrés par l’exposition répétée à une substance et qui mènent à une perte de contrôle de la consommation. La perturbation prolongée des facteurs de transcription vient souligner toute l’importance des mécanismes épigénétiques, lesquels réfèrent à des processus qui provoquent des modifications de la fonction des gènes sans en modifier la séquence. La contribution de ces mécanismes aux modifications pathologiques prolongées survenant après la prise répétée de drogues est certainement trop complexe pour être entièrement résumée ici, mais elle a cependant fait l’objet de différentes revues de littérature (Renthal et Nestler, 2008). Par ailleurs, des facteurs qui influencent la croissance cellulaire, notamment le BDNF (brain-derived neurotrophic factor), sont impliqués dans plusieurs changements neurobiologiques et comportementaux associés à l’exposition aux drogues (Russo et coll., 2009). Le phénomène de sensibilisation aux effets comportementaux des drogues (Boileau et coll., 2006 ; Cornish et Kalivas, 2001), qui fait référence à l’augmentation progressive de la réponse comportementale (par exemple : augmentation de l’activité motrice chez l’animal ou psychomotrice chez l’humain) lors d’une exposition répétée à une drogue (particulièrement les stimulants), serait en effet en partie expliqué par la modulation du BDNF par les drogues (Russo et coll., 2009). Aussi, le BDNF est impliqué dans l’augmentation progressive des mesures de rechute dans les mois suivant le sevrage, phénomène nommé « incubation » (Lu et coll., 2004). Vu l’implication suggérée de l’incubation et de la sensibilisation comportementale aux drogues dans la toxicomanie, les facteurs de croissance neuronale pourraient donc aussi être ciblés afin de traiter cette problématique.
Rôle des projections glutamatergiques dans la rechute
Les phénomènes neurobiologiques décrits jusqu’ici permettent de comprendre certains des processus qui expliquent le passage d’une consommation sociale à une consommation compulsive de drogue. La stimulation des circuits dopaminergiques vient prendre en otage les structures en charge de la récompense, de la saillance motivationnelle et de l’apprentissage de nouveaux comportements. Comment peut-on expliquer que le cerveau ne met pas en place de processus qui permettraient d’inhiber cette tendance à consommer compulsivement la substance ? L’efficacité partielle des stratégies de restauration de la transmission dopaminergique pour prévenir la rechute vient suggérer que d’autres systèmes pourraient contribuer à ce phénomène. Des modèles animaux ont permis de jeter un nouvel éclairage et d’amener certaines pistes de réponses à cette question, notamment quant à la contribution de la neuroplasticité au sein des circuits glutamatergiques pour expliquer les comportements de recherche de drogue qui persistent malgré une période soutenue d’abstinence (Kalivas et coll., 2009 ; LaLumiere et Kalivas, 2008). Le glutamate compte pour près de 70 % de la neurotransmission cérébrale et des projections glutamatergiques sont retrouvées sur plusieurs composantes des circuits dopaminergiques en jeu dans la toxicomanie (ATV, NAc, CPF et amygdale). Cette localisation vient suggérer la pertinence de l’interaction entre glutamate et dopamine dans les mécanismes neurobiologiques sous-jacents à la dépendance aux drogues (Gass et Olive, 2008). Or, on a remarqué dans le NAc et le CPF, deux structures centrales dans la toxicomanie, une altération de la libération de glutamate, des récepteurs glutamatergiques et de la densité en épine dendritique à la suite d’une consommation répétée de substances (Kalivas et O’Brien, 2008 ; Gass et Olive, 2008). Aussi, la rechute provoquée par la drogue et les stimuli qui lui sont associés est liée à une augmentation des niveaux de glutamate dans le NAc, ce qui nécessite des projections glutamatergiques du CPF au corps du NAc (LaLumiere et Kalivas, 2008). Ces résultats ont contribué à l’élaboration d’un modèle de la toxicomanie caractérisé par la prise compulsive de drogue qui est expliquée par des processus dopaminergiques, auxquels succèdent, pendant l’abstinence, des comportements de recherche de drogue qui proviennent de processus largement glutamatergiques (Kalivas et O’Brien, 2008 ; Kalivas et coll., 2009 ; Knackstedt et Kalivas, 2009). Le lien entre les facteurs environnementaux à l’origine de la rechute et les mécanismes neurobiologiques sous-jacents à la dépendance aux drogues, bien qu’encore largement inexploré, semble vouloir être en voie d’être mieux caractérisé.
Tant les études cliniques que précliniques ont démontré que la rechute peut être causée par plusieurs facteurs, soit la prise de drogue, le stress ou l’exposition à des stimuli associés à la prise de drogue (Sinha, 2009 ; Aharonovich et coll., 2006 ; McKay et coll., 1999). Le système glutamatergique semble crucial quant aux processus neurobiologiques qui expliquent ces phénomènes. Les modèles animaux ont permis d’identifier ces processus, en imitant de façon relativement fidèle le développement d’une toxicomanie chez l’humain (Haney et Spealman, 2008 ; Stewart, 2008). Dans un de ces paradigme, l’animal contrôle la prise de substance (en appuyant sur un levier, ce qui entraîne l’administration d’une dose de drogue) et peut donc être étudié pendant les différentes phases de la dépendance : l’acquisition, la prise continue de drogue, le sevrage, l’abstinence et la rechute. La rechute, décrite dans ce modèle comme l’expression de comportements de recherche de drogue malgré l’absence d’administration de drogue à la suite de la pression sur le levier, peut être évoquée en soumettant l’animal à une variété de stimuli pendant la période d’abstinence. Ce modèle a permis de mettre en évidence les circuits impliqués dans la rechute, qui se superposent en partie selon le stimulus en cause. La recherche de substance suivant l’exposition à une drogue implique l’innervation dopaminergique du cortex préfrontal médian (CPFm) et de la coquille du NAc en provenance de l’aire tegmentale ventrale (ATV), mais également les projections glutamatergiques du CPFm au NAc et à l’ATV. Quant à la rechute liée à des stimuli associés à la prise de drogue, tant les projections dopaminergiques de l’ATV à l’amygdale basolatérale (ABL), au CPFm et au coeur du NAc que l’innervation glutamatergique du CPF et de l’ABL au NAc, et du CPF à l’ATV, sont impliquées (Feltenstein et See, 2008 ; Stewart, 2008). Peu importe la substance abusée, les études animales et humaines ont montré que le stress est aussi un des facteurs impliqués dans la rechute (Sinha et Li, 2007), parfois davantage que la prise de drogue elle-même (Shaham et coll., 1997). L’innervation dopaminergique n’y jouerait qu’un rôle de modulation, l’apport de la noradrénaline et de la corticolibérine au sein des projections du CPFm au corps du NAc via le noyau central de l’amygdale et le noyau latéral de la strie terminale étant plus central (Feltenstein et See, 2008).
La rechute semble donc modulée par des mécanismes glutamatergiques, particulièrement dans le contexte d’exposition à des stimuli associés à la substance. Peter Kalivas et son groupe ont même suggéré que les projections glutamatergiques corticostriatales seraient en quelque sorte la voie finale commune nécessaire dans le phénomène de la rechute, pouvant être la cible de nouvelles interventions dans le traitement de la toxicomanie (Kalivas et coll., 2009).
Craving : paradigme et application thérapeutique
Bien que les modèles animaux aient contribué significativement à préciser les processus neurobiologiques qui jouent un rôle dans la dépendance aux drogues, plus particulièrement la rechute, il n’en demeure pas moins que le développement de modèle permettant d’étudier ce phénomène chez l’humain demeure essentiel afin de valider les théories développées chez l’animal et d’évaluer de potentiels traitements ciblant, par exemple, le système glutamatergique. Phénomène bien connu en clinique, le craving a également été étudié systématiquement dans le milieu de la recherche. Tiffany et ses collaborateurs ont notamment contribué au développement d’échelles permettant de mesurer le désir de consommer, offrant ainsi un outil de recherche standardisé qui peut être plus aisément disséminé (Tiffany et coll., 1993). Or, la validité écologique de l’administration aléatoire de cet outil est limitée dans la plupart des situations expérimentales si le sujet n’est pas exposé à une situation susceptible de provoquer un épisode de craving. Le développement de paradigmes utilisant l’exposition à des contextes qui engendrent le craving a permis de confirmer la validité de ce phénomène sur le plan conceptuel. En effet, une variété de stimuli causent des symptômes physiques et psychologiques de façon constante et relativement prévisible (Carter et Tiffany, 1999). Bien que ce sujet dépasse nettement le cadre de cet article, certains groupes ont développé différents paradigmes qui utilisent notamment l’imagerie (Sinha, 2009) et la vidéo (Childress et coll., 1999) afin de provoquer le craving. Ainsi, les sujets sont exposés dans un environnement contrôlé à des scénarios, des images ou des vidéos étant source de stress ou qui rappellent la consommation de substance. Il est alors possible d’évaluer l’intensité du craving avant et après l’exposition aux stimuli, et de déterminer l’impact d’interventions spécifiques administrées à différents moments dans le protocole. L’émergence d’un modèle qui permet de reproduire assez fidèlement et de façon standardisée un phénomène répandu, tout en respectant les considérations éthiques suscitées par ce type de situation, est une étape cruciale dans le développement de nouvelles avenues thérapeutiques. Le fait que les mesures du craving provoqué par différents stimuli (par ex. : stress, stimuli associés à la prise de drogue) prédisent le risque de rechute (pour une revue, voir Sinha et Li, 2007) souligne tout le potentiel de ce paradigme afin d’étudier les processus neurobiologiques et les stratégies d’intervention liés à cette composante de la toxicomanie.
En utilisant ce même paradigme afin de provoquer le craving chez des sujets dépendants à la cocaïne, le groupe de Peter Kalivas a démontré la capacité de la N-acétylcystéine, une molécule restaurant les niveaux de glutamate, de diminuer le craving engendré par des images associées à la consommation de cocaïne (LaRowe et coll., 2007). Cette étude souligne le potentiel de la modulation du système glutamatergique dans le traitement du craving. D’autres systèmes pourraient également être ciblés à cet effet. La lofexidine, un agoniste des récepteurs alpha2-adrénergiques, a augmenté significativement l’abstinence et diminué la rechute dans une étude où elle était utilisée en combinaison avec la naltrexone chez des sujets dépendants à l’héroïne. Les patients sous la combinaison de médications avaient une réponse physiologique et des mesures de craving moins élevées à la suite d’une exposition (en laboratoire) à des stimuli liés au stress et à la prise de drogue que les patients uniquement sous naltrexone (Sinha et coll., 2007). Or, le même groupe a démontré la relative inefficacité de la naltrexone pour diminuer le craving dans cette population (Hyman et coll., 2007). Il est plutôt intriguant de constater que le craving est diminué par cette même molécule chez les patients dépendants à l’alcool (Mann et coll., 2009 ; Myrick et coll., 2008 ; Ooteman et coll., 2007), un polymorphisme du gène du récepteur opioïde de type mu étant un médiateur significatif de cet effet (McGeary et coll., 2006). Il est aussi intéressant de constater que l’acamprosate, une médication qualifiée « d’anticraving », diminue le craving engendré par l’exposition à l’alcool, mais se montre peu efficace à traiter le craving provoqué par un stimulus associé à la consommation d’alcool (Hammarberg et coll., 2009 ; Ooteman et coll., 2007). Différents facteurs semblent moduler l’effet des interventions sur le craving, notamment la substance utilisée, certains polymorphismes et le type de stimulus utilisé. L’efficacité des interventions thérapeutiques pourrait être optimisée en personnalisant le choix de la médication à l’aide de ces facteurs, qui devront dans un premier temps être mieux caractérisés.
Le système cannabinoïde comme cible de traitement du craving et de la rechute
La modulation directe des systèmes dopaminergique et glutamatergique, qui comme nous l’avons expliqué sont largement responsables de la vulnérabilité chronique à la rechute, s’est avérée jusqu’à maintenant inconsistante comme stratégie de traitement du craving. Une des explications pourrait provenir du fait que cette stratégie ne permet pas de moduler spécifiquement la transmission au sein des structures impliquées dans le craving et la rechute. Une stratégie alternative serait donc de moduler indirectement, mais de façon plus spécifique, la transmission glutamatergique et/ou dopaminergique au sein du système mésocorticolimbique, site où se joue la neuroplasticité en cause dans la toxicomanie. Le système endocannabinoïde, un régulateur critique de la fonction synaptique, s’avère à notre avis une cible prometteuse à cet égard (Katona et Freund, 2008).
Le système endocannabinoïde est constitué de plusieurs ligands, enzymes et récepteurs. Le récepteur de type CB1 (CB1R) est reconnu comme l’élément principal qui explique l’effet psychoactif du THC, principal constituant du cannabis et agoniste des récepteurs CB1. Or, ces récepteurs cannabinoïdes sont positionnés au sein de structures critiques pour la toxicomanie en général, et plus spécifiquement le craving et la rechute. Ils sont localisés sur les neurones glutamatergiques du système mésocorticolimbique impliqués dans la rechute et le craving, et ont la capacité de moduler leur fonctionnement (Ferraro et coll., 2001 ; Huang et coll., 2001 ; Lopez-Moreno et coll., 2008 ; Mereu et coll., 2003 ; Shen et coll., 1996). Leur disposition sur les interneurones GABAergiques et les neurones glutamatergiques qui régulent les neurones dopaminergiques au niveau mésolimbique est particulièrement intriguante (Riegel et Lupica, 2004 ; Lupica et Riegel, 2005). Elle expliquerait notamment la capacité des agonistes des récepteurs CB1, tel le THC, d’augmenter indirectement les niveaux de dopamine striatal et mésocorticolimbique qui modulent la cognition et la motivation (Fadda et coll., 2006 ; Malone et Taylor, 1999 ; Bossong et coll., 2008). De plus, les récepteurs opioïdes de type mu sont colocalisés avec les récepteurs CB1 sur les neurones qui projettent du NAc et du striatum dorsal, ceux-ci modulant le renforcement, les comportements dirigés et l’acquisition de comportements en jeu dans la toxicomanie ; l’interaction des récepteurs opioïdes et CB1 modifierait aussi la relâche de dopamine dans le NAc (Rodriguez et coll., 2001 ; Schoffelmeer et coll., 2006).
Faisant écho à ces caractéristiques du système endocannabinoïde, le THC peut non seulement en soi devenir source de dépendance comme les autres substances qui augmentent la dopamine au niveau striatal (Cooper et Haney, 2009), mais peut dans certaines circonstances être associé à la prise d’autres substances telle l’héroïne (Ellgren et coll., 2007 ; Spano et coll., 2007). L’effet délétère de certains agonistes des récepteurs CB1, tel le THC, sur les comportements associés à la toxicomanie suggère que des antagonistes de ces mêmes récepteurs pourraient être utilisés à des fins thérapeutiques. Le rimonabant, un antagoniste synthétique des récepteurs CB1, s’est d’ailleurs avéré efficace afin d’augmenter le taux d’arrêt du tabagisme. L’émergence de symptômes dépressifs et d’idées suicidaires secondaires à cette médication a soulevé d’importantes inquiétudes quant à l’utilisation répandue éventuelle de cette substance (Cahill et Ussher, 2007). Une étude récente aurait toutefois montré des taux relativement faibles de ce type d’effets secondaires, que ce soit lorsque le rimonabant est administré seul ou en combinaison avec la thérapie de remplacement de la nicotine (Rigotti et coll., 2009). Or, des antagonistes des récepteurs CB1 pourraient être utilisés dans le traitement de la dépendance à d’autres substances. Déjà, plusieurs groupes ont démontré le rôle potentiel d’antagonistes des récepteurs CB1 dans la modulation des phénomènes neurobiologiques en jeu dans la dépendance à la cocaïne, l’alcool, la kétamine et aux opioïdes (Alvarez-Jaimes et coll., 2008 ; Cippitelli et coll., 2005 ; Economidou et coll., 2006 ; Li et coll., 2008 ; Li et coll., 2009 ; Xi et coll., 2008). Ces modulateurs du système cannabinoïde s’avèrent efficaces afin d’éviter la rechute induite par l’exposition à la drogue ou à un stimulus associé (mais non par le stress) dans les études animales (Fattore et coll., 2007a). Ces résultats prometteurs viennent s’ajouter à l’ensemble des évidences disponibles soutenant l’importance du système endocannabinoïde dans le phénomène de la rechute et des comportements de recherche de drogues associés à la toxicomanie, et le potentiel que présentent les stratégies visant ce système de transmission (Fattore et coll., 2007a ; Fattore et coll., 2007b ; Maldonado et coll., 2006 ; Solinas et coll., 2007 ; Wiskerke et coll., 2008). L’impact de déterminants génétiques du système endocannabinoïde sur le craving (Haughey et coll., 2008) suggère que ce système pourrait être la cible d’intervention pour moduler le risque de rechute. La disponibilité d’un paradigme valide afin d’évaluer le craving en laboratoire rend d’autant plus attrayante cette stratégie. Un modulateur du système cannabinoïde pourrait donc hypothétiquement diminuer le risque de rechute en atténuant le craving induit par l’exposition à la drogue ou à un stimulus y étant associé. Une telle médication pourrait alors être intégrée à un programme de traitement incluant des interventions psychologiques qui viserait à permettre au patient de développer une résilience aux stimuli environnementaux qui sont souvent à l’origine de la perpétuation du cycle destructeur de la consommation.
Conclusion
La toxicomanie a été longtemps conceptualisée sur le plan neurobiologique par un modèle ne prenant principalement en compte que la modulation à court terme des différents systèmes de neurotransmission. Les stratégies thérapeutiques émanant de cette théorie, bien qu’efficaces pour traiter les symptômes d’intoxication et de sevrage, ont montré certaines limites dans leur capacité à favoriser la reprise de contrôle ou l’arrêt de la consommation de façon durable pour une proportion significative de patients. Plus récemment, la compréhension des modifications neurobiologiques prolongées expliquant la dépendance aux drogues a mis en évidence le rôle, l’interaction et la plasticité de différents systèmes de neurotransmission au sein des structures mésocorticolimbiques, de même que certains processus épigénétiques à l’origine de changements fonctionnels durables au sein de ces composantes cérébrales. La modulation de ces systèmes s’avère particulièrement prometteuse dans le développement de nouvelles stratégies de traitement biologique de la toxicomanie. De nouvelles interventions pourraient potentialiser l’effet thérapeutique des traitements existants ou représenter des alternatives essentielles à ceux-ci. Pour atteindre cet objectif, l’attention devra être réorientée vers les modifications neurobiologiques à long terme associées à l’utilisation chronique d’une drogue, compte tenu des évidences grandissantes à l’appui de leur rôle au coeur même de la toxicomanie.
Plusieurs aspects n’ont pas été explorés dans cet article, bien qu’étant cruciaux dans l’amélioration des services offerts aux toxicomanes. Par exemple, le traitement des adolescents, dont la maturation cérébrale n’est pas complétée, pourrait être la cible d’intervention spécifique. La flexibilité relative du neurodéveloppement à ce stade pourrait s’avérer une limite thérapeutique importante, mais semble également être une fenêtre privilégiée d’intervention. Aussi, l’étude plus approfondie des changements neurobiologiques qui sont engendrés par les interventions psychosociales permettrait de mieux caractériser leur impact et possiblement de mieux adapter et personnaliser ces approches afin de répondre de façon plus adéquate aux besoins de l’individu aux prises avec un problème de consommation.
Il apparaît important à ce stade de considérer avec ouverture différentes approches vu l’ampleur de la problématique. L’utilisation de modulateurs du système cannabinoïde dans le traitement de la toxicomanie, alors que le cannabis a récemment été source d’attention considérable pour le risque de dépendance qui y est associé, pourrait s’avérer à notre avis un exemple judicieux d’une telle ouverture.
Appendices
Remerciements
La préparation de cet article a été soutenue par une bourse de la Fondation du Centre hospitalier de l’Université de Montréal (DJA).
Bibliographie
- Aharonovich, E., Garawi, F., Bisaga, A., Brooks, D., Raby, W. N., Rubin, E., Nunes, E. V. et Levin, F. R. (2006). « Concurrent cannabis use during treatment for comorbid ADHD and cocaine dependence: effects on outcome ». American Journal of Drug and Alcohol Abuse. 32 (4), p. 629-635.
- Alvarez-Jaimes, L., Polis, I. et Parsons, L. H. (2008). « Attenuation of cue-induced heroin-seeking behavior by cannabinoid CB1 antagonist infusions into the nucleus accumbens core and prefrontal cortex, but not basolateral amygdala ». Neuropsychopharmacology. 33 (10), p. 2483-2493.
- Amato, L., Davoli, M., Minozzi, S., Ali, R. et Ferri, M. (2005). « Methadone at tapered doses for the management of opioid withdrawal ». Cochrane Database of Systematic Reviews. (3), CD003409.
- Amato, L., Minozzi, S., Pani, P. P. et Davoli, M. (2007). « Antipsychotic medications for cocaine dependence ». Cochrane Database of Systematic Reviews. (3), CD006306.
- American Psychiatric Association (APA). (2000). Diagnostic and Statistical Manual of Mental Disorders. Fourth Edition. Text Revision. Washington DC : American Psychiatric Association, 943 p.
- Bao, Y. P., Liu, Z. M., Epstein, D. H., Du, C., Shi, J. et Lu, L. (2009). « A meta-analysis of retention in methadone maintenance by dose and dosing strategy ». American Journal of Drug Alcohol Abuse. 35 (1), p. 28-33.
- Boileau, I., Dagher, A., Leyton, M., Gunn, R. N., Baker, G. B., Diksic, M. et Benkelfat, C. (2006). « Modeling sensitization to stimulants in humans: an [11C]raclopride/positron emission tomography study in healthy men ». Archives of General Psychiatry. 63 (12), p. 1386-1395.
- Bossong, M. G., van Berckel, B. N., Boellaard, R., Zuurman, L., Schuit, R. C., Windhorst, A. D., van Gerven, J. M., Ramsey, N. F., Lammertsma, A. A. et Kahn, R. S. (2008). « Delta9-tetrahydrocannabinol induces dopamine release in the human striatum ». Neuropsychopharmacology. 34 (3), p. 759-766.
- Cahill, K. et Ussher, M. (2007). « Cannabinoid type 1 receptor antagonists (rimonabant) for smoking cessation ». Cochrane Database of Systematic Reviews. (4), CD005353.
- Cahill, K., Stead, L. F. et Lancaster, T. (2008). « Nicotine receptor partial agonists for smoking cessation ». Cochrane Database of Systematic Reviews. (3), CD006103.
- Carter, B. L. et Tiffany, S. T. (1999). « Meta-analysis of cue-reactivity in addiction research ». Addiction. 94 (3), p. 327-340.
- Castells, X., Casas, M., Vidal, X., Bosch, R., Roncero, C., Ramos-Quiroga, J. A. et Capella, D. (2007). « Efficacy of central nervous system stimulant treatment for cocaine dependence: a systematic review and meta-analysis of randomized controlled clinical trials ». Addiction. 102 (12), p. 1871-1887.
- Childress, A. R., Mozley, P. D., McElgin, W., Fitzgerald, J., Reivich, M. et O’Brien, C. P. (1999). «Limbic activation during cue-induced cocaine craving». American Journal of Psychiatry. 156 (1), p. 11-18.
- Cippitelli, A., Bilbao, A., Hansson, A. C., del Arco, I., Sommer, W., Heilig, M., Massi, M., Bermudez-Silva, F. J., Navarro, M., Ciccocioppo, R. et de Fonseca, F. R. (2005). « Cannabinoid CB1 receptor antagonism reduces conditioned reinstatement of ethanol-seeking behavior in rats ». European Journal of Neuroscience. 21 (8), p. 2243-2251.
- Connock, M., Juarez-Garcia, A., Jowett, S., Frew, E., Liu, Z., Taylor, R. J., Fry-Smith, A., Day, E., Lintzeris, N., Roberts, T., Burls, A. et Taylor, R. S. (2007). « Methadone and buprenorphine for the management of opioid dependence: a systematic review and economic evaluation ». Health Technology Assessment. 11 (9), p. 1-171, iii-iv.
- Cooper, Z. D. et Haney, M. (2009). « Actions of delta-9-tetrahydrocannabinol in cannabis: relation to use, abuse, dependence ». International Review of Psychiatry. 21 (2), p. 104-112.
- Cornish, J. L. et Kalivas, P. W. (2001). « Repeated cocaine administration into the rat ventral tegmental area produces behavioral sensitization to a systemic cocaine challenge ». Behavioral Brain Research. 126 (1-2), p. 205-209.
- Dijkgraaf, M. G., van der Zanden, B. P., de Borgie, C. A., Blanken, P., van Ree, J. M. et van den Brink, W. (2005). « Cost utility analysis of co-prescribed heroin compared with methadone maintenance treatment in heroin addicts in two randomised trials ». British Medical Journal. 330 (7503), p. 1297.
- Dole, V. P. et Nyswander, M. (1965). « A medical treatment for diacetylmorphine (heroin) addiction. A clinical trial with methadone hydrochloride ». Journal of the American Medical Association. 193 (8), p. 646-650.
- Eap, C. B., Buclin, T. et Baumann, P. (2002). « Interindividual variability of the clinical pharmacokinetics of methadone: implications for the treatment of opioid dependence ». Clinical Pharmacokinetics. 41 (14), p. 1153-1193.
- Economidou, D., Mattioli, L., Cifani, C., Perfumi, M., Massi, M., Cuomo, V., Trabace, L. et Ciccocioppo, R. (2006). « Effect of the cannabinoid CB1 receptor antagonist SR-141716A on ethanol self-administration and ethanol-seeking behaviour in rats ». Psychopharmacology (Berl). 183 (4), p. 394-403.
- Eder, H., Jagsch, R., Kraigher, D., Primorac, A., Ebner, N. et Fischer, G. (2005). « Comparative study of the effectiveness of slow-release morphine and methadone for opioid maintenance therapy ». Addiction. 100 (8), p. 1101-1109.
- Elkader, A. et Sproule, B. (2005). « Buprenorphine: clinical pharmacokinetics in the treatment of opioid dependence ». Clinical Pharmacokinetics. 44 (7), p. 661-680.
- Elkashef, A., Fudala, P. J., Gorgon, L., Li, S. H., Kahn, R., Chiang, N., Vocci, F., Collins, J., Jones, K., Boardman, K. et Sather, M. (2006). « Double-blind, placebo-controlled trial of selegiline transdermal system (STS) for the treatment of cocaine dependence ». Drug and Alcohol Dependence. 85 (3), p. 191-197.
- Elkashef, A., Vocci, F., Hanson, G., White, J., Wickes, W. et Tiihonen, J. (2008). « Pharmacotherapy of methamphetamine addiction: an update ». Substance Abuse. 29 (3), p. 31-49.
- Elkashef, A. M., Rawson, R. A., Anderson, A. L., Li, S. H., Holmes, T., Smith, E. V., Chiang, N., Kahn, R., Vocci, F., Ling, W., Pearce, V. J., McCann, M., Campbell, J., Gorodetzky, C., Haning, W., Carlton, B., Mawhinney, J. et Weis, D. (2008). « Bupropion for the treatment of methamphetamine dependence ». Neuropsychopharmacology. 33 (5), p. 1162-1170.
- Ellgren, M., Spano, S. M. et Hurd, Y. L. (2007). « Adolescent cannabis exposure alters opiate intake and opioid limbic neuronal populations in adult rats ». Neuropsychopharmacology. 32 (3), p. 607-615.
- Fadda, P., Scherma, M., Spano, M. S., Salis, P., Melis, V., Fattore, L. et Fratta, W. (2006). « Cannabinoid self-administration increases dopamine release in the nucleus accumbens ». NeuroReport. 17 (15), p. 1629-1632.
- Fattore, L., Spano, M. S., Deiana, S., Melis, V., Cossu, G., Fadda, P. et Fratta, W. (2007a). « An endocannabinoid mechanism in relapse to drug seeking: a review of animal studies and clinical perspectives ». Brain Research Reviews. 53 (1), p. 1-16.
- Fattore, L., Fadda, P. et Fratta, W. (2007b). « Endocannabinoid regulation of relapse mechanisms ». Pharmacology Research. 56 (5), p. 418-427.
- Feltenstein, M. W. et See, R. E. (2008). « The neurocircuitry of addiction: an overview ». British Journal of Pharmacology. 154 (2), p. 261-274.
- Ferraro, L., Tomasini, M. C., Gessa, G. L., Bebe, B. W., Tanganelli, S. et Antonelli, T. (2001). «The cannabinoid receptor agonist WIN 55,212-2 regulates glutamate transmission in rat cerebral cortex: an in vivo and in vitro study ». Cerebral Cortex. 11 (8), p. 728-733.
- Galanter, M. et Kleber, H. D. (2008). The American Psychiatric Publishing Textbook of Substance Abuse Treatment. Washington, D.C. : American Psychiatric Publishing, 752 p.
- Garrido, M. J. et Troconiz, I. F. (1999). « Methadone: a review of its pharmacokinetic/pharmacodynamic properties ». Journal of Pharmacological and Toxicological Methods. 42 (2), p. 61-66.
- Gartry, C. C., Oviedo-Joekes, E., Laliberte, N. et Schechter, M. T. (2009). « NAOMI: The trials and tribulations of implementing a heroin assisted treatment study in North America ». Harm Reduction Journal. 6, p. 2.
- Gass, J. T. et Olive, M. F. (2008). « Glutamatergic substrates of drug addiction and alcoholism ». Biochemical Pharmacology. 75 (1), p. 218-265.
- Gibson, A., Degenhardt, L., Mattick, R. P., Ali, R., White, J. et O’Brien, S. (2008). « Exposure to opioid maintenance treatment reduces long-term mortality ». Addiction. 103 (3), p. 462-468.
- Gossop, M., Stewart, D., Browne, N. et Marsden, J. (2002). « Factors associated with abstinence, lapse or relapse to heroin use after residential treatment: protective effect of coping responses ». Addiction. 97 (10), p. 1259-1267.
- Haasen, C., Verthein, U., Degkwitz, P., Berger, J., Krausz, M. et Naber, D. (2007). « Heroin-assisted treatment for opioid dependence: randomised controlled trial ». British Journal of Psychiatry. 191 (1), p. 55-62.
- Hammarberg, A., Jayaram-Lindstrom, N., Beck, O., Franck, J. et Reid, M. S. (2009). « The effects of acamprosate on alcohol-cue reactivity and alcohol priming in dependent patients: a randomized controlled trial ». Psychopharmacology (Berl).205 (1), p. 53-62.
- Haney, M., Ward, A. S., Foltin, R. W. et Fischman, M. W. (2001). « Effects of ecopipam, a selective dopamine D1 antagonist, on smoked cocaine self-administration by humans ». Psychopharmacology (Berl). 155 (4), p. 330-337.
- Haney, M. et Spealman, R. (2008). « Controversies in translational research: drug self-administration ». Psychopharmacology (Berl). 199 (3), p. 403-419.
- Haughey, H. M., Marshall, E., Schacht, J. P., Louis, A. et Hutchison, K. E. (2008). « Marijuana withdrawal and craving: influence of the cannabinoid receptor 1 (CNR1) and fatty acid amide hydrolase (FAAH) genes ». Addiction. 103 (10), p. 1678-1686.
- Hopwood, M., Southgate, E., Kippax, S., Bammer, G., Isaac-Toua, G. et MacDonald, M. (2003). « The injection of methadone syrup in New South Wales: patterns of use and increased harm after partial banning of injecting equipment ». Australian and New Zealand Journal of Public Health. 27 (5), p. 551-555.
- Huang, C. C., Lo, S. W. et Hsu, K. S. (2001). « Presynaptic mechanisms underlying cannabinoid inhibition of excitatory synaptic transmission in rat striatal neurons ». The Journal of Physiology. 532 (Pt 3), p. 731-748.
- Hyman, S. M., Fox, H., Hong, K. I., Doebrick, C. et Sinha, R. (2007). « Stress and drug-cue-induced craving in opioid-dependent individuals in naltrexone treatment ». Experimental and Clinical Psychopharmacology. 15 (2), p. 134-143.
- Ilgen, M., Jain, A., Kim, H. M. et Trafton, J. A. (2008). « The effect of stress on craving for methadone depends on the timing of last methadone dose ». Behaviour Research and Therapy. 46 (10), p. 1170-1175.
- Johansson, B. A., Berglund, M. et Lindgren, A. (2006). « Efficacy of maintenance treatment with naltrexone for opioid dependence: a meta-analytical review ». Addiction. 101 (4), p. 491-503.
- Johansson, B. A., Berglund, M. et Lindgren, A. (2007). « Efficacy of maintenance treatment with methadone for opioid dependence: a meta-analytical study ». Nordic Journal of Psychiatry. 61 (4), p. 288-295.
- Kalivas, P. W. et O’Brien, C. (2008). « Drug addiction as a pathology of staged neuroplasticity ». Neuropsychopharmacology. 33 (1), p. 166-180.
- Kalivas, P. W., Lalumiere, R. T., Knackstedt, L. et Shen, H. (2009). « Glutamate transmission in addiction ». Neuropharmacology. 56 (Suppl 1), p. 169-173.
- Kastelic, A., Dubajic, G. et Strbad, E. (2008). « Slow-release oral morphine for maintenance treatment of opioid addicts intolerant to methadone or with inadequate withdrawal suppression ». Addiction. 103 (11), p. 1837-1846.
- Katona, I. et Freund, T. F. (2008). « Endocannabinoid signaling as a synaptic circuit breaker in neurological disease ». Nature Medecine. 14 (9), p. 923-930.
- Kinsey, B. M., Jackson, D. C. et Orson, F. M. (2009). « Anti-drug vaccines to treat substance abuse ». Immunology & Cell Biology. 87 (4), p. 309-314.
- Kleber, H. D., Weiss, R. D., Anton, R. F., Jr., George, T. P., Greenfield, S. F., Kosten, T. R., O’Brien, C. P., Rounsaville, B. J., Strain, E. C., Ziedonis, D. M., Hennessy, G., Connery, H. S., McIntyre, J. S., Charles, S. C., Anzia, D. J., Cook, I. A., Finnerty, M. T., Johnson, B. R., Nininger, J. E., Summergrad, P., Woods, S. M., Yager, J., Pyles, R., Cross, C. D., Peele, R., Shemo, J. P., Lurie, L., Walker, R. D., Barnovitz, M. A., Gray, S. H., Saxena, S., Tonnu, T., Kunkle, R., Albert, A. B., Fochtmann, L. J., Hart, C., Regier, D., Work Group on Substance Use, Disorders, American Psychiatric, Association et Steering Committee on Practice, Guidelines. (2007). « Treatment of patients with substance use disorders, second edition. American Psychiatric Association ». American Journal of Psychiatry. 164 (4 Suppl), p. 5-123.
- Knackstedt, L. A. et Kalivas, P. W. (2009). « Glutamate and reinstatement ». Current Opinion in Pharmacology. 9 (1), p. 59-64.
- Kraigher, D., Jagsch, R., Gombas, W., Ortner, R., Eder, H., Primorac, A. et Fischer, G. (2005). « Use of slow-release oral morphine for the treatment of opioid dependence ». European Addiction Research. 11 (3), p. 145-151.
- LaLumiere, R. T. et Kalivas, P. W. (2008). « Glutamate release in the nucleus accumbens core is necessary for heroin seeking ». Journal of Neuroscience. 28 (12), p. 3170-3177.
- Lancaster, T. et Stead, L. F. (2000). « Mecamylamine (a nicotine antagonist) for smoking cessation ». Cochrane Database of Systematic Reviews. (2), CD001009.
- Langleben, D. D., Ruparel, K., Elman, I., Busch-Winokur, S., Pratiwadi, R., Loughead, J., O’Brien, C. P. et Childress, A. R. (2008). « Acute effect of methadone maintenance dose on brain FMRI response to heroin-related cues ». American Journal of Psychiatry. 165 (3), p. 390-394.
- LaRowe, S. D., Myrick, H., Hedden, S., Mardikian, P., Saladin, M., McRae, A., Brady, K., Kalivas, P. W. et Malcolm, R. (2007). « Is cocaine desire reduced by N-acetylcysteine? ». American Journal of Psychiatry. 164 (7), p. 1115-1117.
- Levin, F. R. et Kleber, H. D. (2008). « Use of dronabinol for cannabis dependence: two case reports and review ». American Journal Addictions. 17 (2), p. 161-164.
- Li, F., Fang, Q., Liu, Y., Zhao, M., Li, D., Wang, J. et Lu, L. (2008). « Cannabinoid CB(1) receptor antagonist rimonabant attenuates reinstatement of ketamine conditioned place preference in rats ». European Journal of Pharmacology. 589 (1-3), p. 122-126.
- Li, X., Hoffman, A. F., Peng, X. Q., Lupica, C. R., Gardner, E. L. et Xi, Z. X. (2009). « Attenuation of basal and cocaine-enhanced locomotion and nucleus accumbens dopamine in cannabinoid CB1-receptor-knockout mice ». Psychopharmacology (Berl). 204 (1), p. 1-11.
- Ling, W. et Wesson, D. R. (1984). « Naltrexone treatment for addicted health-care professionals: a collaborative private practice experience ». Journal of Clinical Psychiatry. 45 (9 Pt 2), p. 46-48.
- Lobmaier, P., Kornor, H., Kunoe, N. et Bjorndal, A. (2008). « Sustained-release naltrexone for opioid dependence ». Cochrane Database of Systematic Reviews. (2), CD006140.
- Lopez-Moreno, J. A., Gonzalez-Cuevas, G., Moreno, G. et Navarro, M. (2008). « The pharmacology of the endocannabinoid system: functional and structural interactions with other neurotransmitter systems and their repercussions in behavioral addiction ». Addiction Biology. 13 (2), p. 160-187.
- Lott, D. C., Strain, E. C., Brooner, R. K., Bigelow, G. E. et Johnson, R. E. (2006). « HIV risk behaviors during pharmacologic treatment for opioid dependence: a comparison of levomethadyl acetate [corrected] buprenorphine, and methadone ». Journal of Substance Abuse Treatment. 31 (2), p. 187-194.
- Lu, L., Grimm, J. W., Hope, B. T. et Shaham, Y. « Incubation of cocaine craving after withdrawal: a review of preclinical data ». Neuropharmacology, 47 (Suppl 1), p. 214-226.
- Lupica, C. R. et Riegel, A. C. (2005). « Endocannabinoid release from midbrain dopamine neurons: a potential substrate for cannabinoid receptor antagonist treatment of addiction ». Neuropharmacology. 48 (8), p. 1105-1116.
- Lutfy, K. et Cowan, A. (2004). « Buprenorphine: a unique drug with complex pharmacology ». Current Neuropharmacology. 2 (4), p. 395-402.
- Magura, S., Lee, S. J., Salsitz, E. A., Kolodny, A., Whitley, S. D., Taubes, T., Seewald, R., Joseph, H., Kayman, D. J., Fong, C., Marsch, L. A. et Rosenblum, A. (2007). « Outcomes of buprenorphine maintenance in office-based practice ». Journal of Addictive Diseases. 26 (2), p. 13-23.
- Maldonado, R., Valverde, O. et Berrendero, F. (2006). « Involvement of the endocannabinoid system in drug addiction ». Trends in Neurosciences. 29 (4), p. 225-232.
- Malone, D. T. et Taylor, D. A. (1999). « Modulation by fluoxetine of striatal dopamine release following Delta9-tetrahydrocannabinol: a microdialysis study in conscious rats ». British Journal of Pharmacology. 128 (1), p. 21-26.
- Mann, K., Kiefer, F., Smolka, M., Gann, H., Wellek, S. et Heinz, A. (2009). « Searching for responders to acamprosate and naltrexone in alcoholism treatment: rationale and design of the predict study ». Alcohol: Clinical and Experimental Research. 33 (4), p. 674-683.
- Margolin, A., Avants, S. K. et Kosten, T. R. (1995). « Mazindol for relapse prevention to cocaine abuse in methadone-maintained patients ». American Journal of Drug and Alcohol Abuse. 21 (4), p. 469-481.
- Mattick, R. P. et Hall, W. (1996). « Are detoxification programmes effective ? ». Lancet. 347 (8994), p. 97-100.
- Mattick, R. P., Breen, C., Kimber, J. et Davoli, M. (2003). « Methadone maintenance therapy versus no opioid replacement therapy for opioid dependence ». Cochrane Database of Systematic Reviews. (2), CD002209.
- Mattick, R. P., Kimber, J., Breen, C. et Davoli, M. (2008). « Buprenorphine maintenance versus placebo or methadone maintenance for opioid dependence ». Cochrane Database of Systematic Reviews. (2), CD002207.
- McGeary, J. E., Monti, P. M., Rohsenow, D. J., Tidey, J., Swift, R. et Miranda, R., Jr. (2006). « Genetic moderators of naltrexone’s effects on alcohol cue reactivity ». Alcohol: Clinical and Experimental Research. 30 (8), p. 1288-1296.
- McKay, J. R., Alterman, A. I., Rutherford, M. J., Cacciola, J. S. et McLellan, A. T. (1999). « The relationship of alcohol use to cocaine relapse in cocaine dependent patients in an aftercare study ». Journal of Studies on Alcohol. 60 (2), p. 176-180.
- Mereu, G., Fa, M., Ferraro, L., Cagiano, R., Antonelli, T., Tattoli, M., Ghiglieri, V., Tanganelli, S., Gessa, G. L. et Cuomo, V. (2003). « Prenatal exposure to a cannabinoid agonist produces memory deficits linked to dysfunction in hippocampal long-term potentiation and glutamate release ». Proceedings of the National Academy of Sciences U S A. 100 (8), p. 4915-4920.
- Millson, P., Challacombe, L., Villeneuve, P. J., Strike, C. J., Fischer, B., Myers, T., Shore, R. et Hopkins, S. (2007). « Reduction in injection-related HIV risk after 6 months in a low-threshold methadone treatment program ». AIDS Education and Prevention. 19 (2), p. 124-136.
- Minozzi, S., Amato, L., Vecchi, S., Davoli, M., Kirchmayer, U. et Verster, A. (2006). « Oral naltrexone maintenance treatment for opioid dependence ». Cochrane Database of Systematic Reviews. (1), CD001333.
- Mitchell, T. B., White, J. M., Somogyi, A. A. et Bochner, F. (2004). « Slow-release oral morphine versus methadone: a crossover comparison of patient outcomes and acceptability as maintenance pharmacotherapies for opioid dependence ». Addiction. 99 (8), p. 940-945.
- Myrick, H., Anton, R. F., Li, X., Henderson, S., Randall, P. K. et Voronin, K. (2008). « Effect of naltrexone and ondansetron on alcohol cue-induced activation of the ventral striatum in alcohol-dependent people ». Archives of General Psychiatry. 65 (4), p. 466-475.
- Nestler, E. J. (2008). « Transcriptional mechanisms of addiction: role of DeltaFosB ». Philosophical Transactions of the Royal Society of London Series B: Biology Sciences. 363 (1507), p. 3245-3255.
- Norrholm, S. D., Bibb, J. A., Nestler, E. J., Ouimet, C. C., Taylor, J. R. et Greengard, P. (2003). « Cocaine-induced proliferation of dendritic spines in nucleus accumbens is dependent on the activity of cyclin-dependent kinase-5 ». Neuroscience. 116 (1), p. 19-22.
- Ntais, C., Pakos, E., Kyzas, P. et Ioannidis, J. P. (2005). « Benzodiazepines for alcohol withdrawal ». Cochrane Database of Systematic Reviews. (3), CD005063.
- O’Brien, C. et Cornish, J. W. (2006). « Naltrexone for probationers and parolees ». Journal of Substance Abuse Treatment. 31 (2), p. 107-111.
- Ooteman, W., Koeter, M. W., Verheul, R., Schippers, G. M. et van den Brink, W. (2007). « The effect of naltrexone and acamprosate on cue-induced craving, autonomic nervous system and neuroendocrine reactions to alcohol-related cues in alcoholics ». European Neuropsychopharmacology. 17 (8), p. 558-566.
- Orson, F. M., Kinsey, B. M., Singh, R. A., Wu, Y., Gardner, T. et Kosten, T. R. (2008). « Substance abuse vaccines ». Annals of the New York Academy of Sciences. 1141, p. 257-269.
- Pierce, R. C. et Kumaresan, V. (2006). « The mesolimbic dopamine system: the final common pathway for the reinforcing effect of drugs of abuse ? ». Neuroscience & Biobehavioral Reviews. 30 (2), p. 215-238.
- Poling, J., Oliveto, A., Petry, N., Sofuoglu, M., Gonsai, K., Gonzalez, G., Martell, B. et Kosten, T. R. (2006). « Six-month trial of bupropion with contingency management for cocaine dependence in a methadone-maintained population ». Archives of General Psychiatry. 63 (2), p. 219-228.
- Renthal, W. et Nestler, E. J. (2008). « Epigenetic mechanisms in drug addiction ». Trends in Molecular Medecine. 14 (8), p. 341-350.
- Riegel, A. C. et Lupica, C. R. (2004). « Independent presynaptic and postsynaptic mechanisms regulate endocannabinoid signaling at multiple synapses in the ventral tegmental area ». Journal of Neuroscience. 24 (49), p. 11070-11078.
- Rigotti, N. A., Gonzales, D., Dale, L. C., Lawrence, D. et Chang, Y. (2009). « A randomized controlled trial of adding the nicotine patch to rimonabant for smoking cessation: efficacy, safety and weight gain ». Addiction. 104 (2), p. 266-276.
- Robinson, T. E. et Kolb, B. (1999). « Morphine alters the structure of neurons in the nucleus accumbens and neocortex of rats ». Synapse. 33 (2), p. 160-162.
- Roche, A., McCabe, S. et Smyth, B. P. (2008). « Illicit methadone use and abuse in young people accessing treatment for opiate dependence ». European of Addiction Research. 14 (4), p. 219-225.
- Rodriguez, J. J., Mackie, K. et Pickel, V. M. (2001). « Ultrastructural localization of the CB1 cannabinoid receptor in mu-opioid receptor patches of the rat Caudate putamen nucleus ». Journal of Neuroscience. 21 (3),p. 823-833.
- Roth, A., Hogan, I. et Farren, C. (1997). « Naltrexone plus group therapy for the treatment of opiate-abusing health-care professionals ». Journal of Substance Abuse Treatment. 14 (1), p. 19-22.
- Russo, S. J., Mazei-Robison, M. S., Ables, J. L. et Nestler, E. J. (2009). « Neurotrophic factors and structural plasticity in addiction ». Neuropharmacology. 56 (Suppl 1), p. 73-82.
- Schoffelmeer, A. N., Hogenboom, F., Wardeh, G. et De Vries, T. J. (2006). « Interactions between CB1 cannabinoid and mu opioid receptors mediating inhibition of neurotransmitter release in rat nucleus accumbens core ». Neuropharmacology. 51 (4), p. 773-781.
- Schultz, W. (1998). « Predictive reward signal of dopamine neurons ». Journal of Neurophysiology. 80 (1), p. 1-27.
- Schultz, W. (2001). « Reward signaling by dopamine neurons ». The Neuroscientist. 7 (4), p. 293-302.
- Shaham, Y., Funk, D., Erb, S., Brown, T. J., Walker, C. D. et Stewart, J. (1997). « Corticotropin-releasing factor, but not corticosterone, is involved in stress-induced relapse to heroin-seeking in rats ». The Journal of Neuroscience. 17 (7), p. 2605-2614.
- Shen, M., Piser, T. M., Seybold, V. S. et Thayer, S. A. (1996). « Cannabinoid receptor agonists inhibit glutamatergic synaptic transmission in rat hippocampal cultures ». The Journal of Neuroscience. 16 (14), p. 4322-4334.
- Shiffman, S., Ferguson, S. G. et Gwaltney, C. J. (2006). « Immediate hedonic response to smoking lapses: relationship to smoking relapse, and effects of nicotine replacement therapy ». Psychopharmacology (Berl). 184 (3-4), p. 608-618.
- Simojoki, K., Vorma, H. et Alho, H. (2008). « A retrospective evaluation of patients switched from buprenorphine (Subutex) to the buprenorphine/naloxone combination (Suboxone) ». Substance Abuse Treatment, Prevention and Policy. 3, p. 16.
- Sinha, R., Kimmerling, A., Doebrick, C. et Kosten, T. R. (2007). « Effects of lofexidine on stress-induced and cue-induced opioid craving and opioid abstinence rates: preliminary findings ». Psychopharmacology (Berl). 190 (4), p. 569-574.
- Sinha, R. et Li, C. S. (2007). « Imaging stress- and cue-induced drug and alcohol craving: association with relapse and clinical implications ». Drug and Alcohol Review. 26 (1), p. 25-31.
- Sinha, R. (2009). « Modeling stress and drug craving in the laboratory: implications for addiction treatment development ». Addiction Biology. 14 (1), p. 84-98.
- Solinas, M., Yasar, S. et Goldberg, S. R. (2007). « Endocannabinoid system involvement in brain reward processes related to drug abuse ». Pharmacological Research. 56 (5), p. 393-405.
- Spano, M. S., Ellgren, M., Wang, X. et Hurd, Y. L. (2007). « Prenatal cannabis exposure increases heroin seeking with allostatic changes in limbic enkephalin systems in adulthood ». Biological Psychiatry. 61 (4), p. 554-563.
- Stead, L. F., Perera, R., Bullen, C., Mant, D. et Lancaster, T. (2008). « Nicotine replacement therapy for smoking cessation ». Cochrane Database of Systematic Reviews. (1), CD000146.
- Stewart, J. (2008). « Review. Psychological and neural mechanisms of relapse ». Philosophical Transactions of the Royal Society of London series B: Biological Sciences. 363 (1507), p. 3147-3158.
- Stine, S. M., Krystal, J. H., Kosten, T. R. et Charney, D. S. (1995). « Mazindol treatment for cocaine dependence ». Drug and Alcohol Dependence. 39 (3), p. 245-252.
- Suh, J. J., Pettinati, H. M., Kampman, K. M. et O’Brien, C. P. (2006). « The status of disulfiram: a half of a century later ». Journal of Clinical Psychopharmacology. 26 (3), p. 290-302.
- Symington, A. (2008). « Results of the first North American prescription heroin study are promising ». HIV/AIDS Policy and Law Review. 13 (2-3), p. 11-12.
- Tiffany, S.T., Fields, L., Singleton, E., Haertzen, C. et Henningfield, J.E. (1993). The development of a heroin craving questionnaire (unpublished manuscript). West Lafayette, Indiana : Purdue University [s. p.].
- Tiihonen, J., Kuoppasalmi, K., Fohr, J., Tuomola, P., Kuikanmaki, O., Vorma, H., Sokero, P., Haukka, J. et Meririnne, E. (2007). « A comparison of aripiprazole, methylphenidate, and placebo for amphetamine dependence ». American Journal of Psychiatry. 164 (1), p. 160-162.
- UNODC. (2008). World Drug Report 2008. Vienna, Austria : United Nations Office on Drugs and Crime, 303 p.
- van den Brink, W., Hendriks, V. M., Blanken, P., Koeter, M. W., van Zwieten, B. J. et van Ree, J. M. (2003). « Medical prescription of heroin to treatment resistant heroin addicts: two randomised controlled trials ». British Medical Journal. 327 (7410), p. 310.
- Volkow, N. D., Fowler, J. S., Wang, G. J., Baler, R. et Telang, F. (2009). « Imaging dopamine’s role in drug abuse and addiction ». Neuropharmacology. 56 (Suppl 1), p. 3-8.
- Waldvogel, D., Figner, B. et Eich, D. (2005). « Illicit methadone injecting during methadone maintenance treatment in a specialised out-patient clinic ». Swiss Medical Weekly. 135 (43-44), p. 644-646.
- Walsh, S. L. et Eissenberg, T. (2003). « The clinical pharmacology of buprenorphine: extrapolating from the laboratory to the clinic ». Drug and Alcohol Dependence. 70 (2 Suppl), p. S13-27.
- Washton, A. M., Pottash, A. C. et Gold , M. S. (1984). « Naltrexone in addicted business executives and physicians ». Journal of Clinical Psychiatry. 45 (9 Pt 2), p. 39-41.
- Waters, A. J., Shiffman, S., Sayette, M. A., Paty, J. A., Gwaltney, C. J. et Balabanis, M. H. (2004). « Cue-provoked craving and nicotine replacement therapy in smoking cessation ». Journal of Consulting and Clinical Psychology. 72 (6), p. 1136-1143.
- Winstock, A. R., Lea, T. et Sheridan, J. (2008). « Prevalence of diversion and injection of methadone and buprenorphine among clients receiving opioid treatment at community pharmacies in New South Wales, Australia ». International Journal of Drug Policy. 19 (6), p. 450-458.
- Wise, R. A. (2004). « Dopamine, learning and motivation ». National Reviews Neuroscience. 5 (6), p. 483-494.
- Wiskerke, J., Pattij, T., Schoffelmeer, A. N. et De Vries, T. J. (2008). « The role of CB1 receptors in psychostimulant addiction ». Addictive Biology. 13 (2), p. 225-238.
- Xi, Z. X., Spiller, K., Pak, A. C., Gilbert, J., Dillon, C., Li, X., Peng, X. Q. et Gardner, E. L. (2008). « Cannabinoid CB1 receptor antagonists attenuate cocaine’s rewarding effects: experiments with self-administration and brain-stimulation reward in rats ». Neuropsychopharmacology. 33 (7), p. 1735-1745.
- Zachariou, V., Bolanos, C. A., Selley, D. E., Theobald, D., Cassidy, M. P., Kelz, M. B., Shaw-Lutchman, T., Berton, O., Sim-Selley, L. J., Dileone, R. J., Kumar, A. et Nestler, E. J. (2006). « An essential role for DeltaFosB in the nucleus accumbens in morphine action ». Nature Neuroscience. 9 (2), p. 205-211.
List of figures
Figure 1
Différentes phases du cycle de la toxicomanie et quelques exemples d’interventions associées
List of tables
Tableau 1
Principaux avantages et limites des médications couramment utilisées dans le traitement de la dépendance aux substances (liste non exhaustive)