

Abstracts
Résumé
Le Parc national de la Salonga (PNS) et ses ressources biologiques revêtent une importance considérable tant sur le plan environnemental qu'économique pour les communautés riveraines qui dépendent de ses ressources pour leur survie. Cette étude vise à quantifier l'impact des activités humaines sur les forêts primaires de la partie sud du PNS en utilisant la classification d'images Landsat des années 2002, 2007, 2014 et 2020, combinée aux outils d'analyse de l'écologie du paysage. Les résultats ont révélé que le paysage reste prédominé par les forêts primaires entre 2002 et 2020, bien que leur superficie ait constamment diminué au profit des forêts secondaires et des zones agricoles. En effet, la superficie des forêts primaires est passée de 12 636,82 km² en 2002 à 11 266 km² en 2020. La perte totale des forêts primaires au cours de cette période s'élève à 10,4 %, soit un taux annuel de déforestation de 0,6 %. Le nombre total de fragments de la forêt primaire a également légèrement diminué, passant de 525 142 en 2002 à 524 584 en 2020. De plus, les résultats ont également révélé que la proportion de forêts primaires augmente à mesure que l'on s'éloigne des villages, indiquant un impact significatif des zones villageoises sur la dégradation forestière. L'anthropisation de la partie sud du PNS se manifeste clairement par la réduction des surfaces de forêts primaires, principalement due aux activités agricoles. Ces résultats soulignent la nécessité d'un suivi régulier des activités humaines au sein du PNS, ainsi que le développement d'une stratégie de gestion intégrée pour préserver cette zone d'une importance écologique cruciale.
Mots-clés :
- Parc national de la Salonga,
- changements anthropiques,
- déforestation,
- télédétection/SIG,
- approche paysagère
Abstract
The Salonga National Park (PNS) and its biological resources hold significant environmental and economic importance for the surrounding communities that rely on its resources for survival. This study aims to quantify the impact of human activities on the primary forests of the southern part of the PNS using Landsat image classification from the years 2002, 2007, 2014, and 2020, combined with landscape ecology analysis tools. The results revealed that the landscape remained predominantly covered by primary forests between 2002 and 2020, although their area consistently decreased in favor of secondary forests and agricultural zones. Specifically, the area of primary forests decreased from 12,636.82 km² in 2002 to 11,266 km² in 2020. The total loss of primary forests during this period amounted to 10.4%, with an annual deforestation rate of 0.6%. The total number of primary forest fragments also slightly decreased, from 525,142 in 2002 to 524,584 in 2020. Additionally, the results indicated that the proportion of primary forests increased as the distance from villages increased, suggesting a significant impact of village areas on forest degradation. The anthropogenic impact on the southern part of the PNS is clearly evidenced by the reduction in primary forest areas, primarily due to agricultural activities. These findings underscore the necessity for regular monitoring of human activities within the PNS and the development of an integrated management strategy to preserve this ecologically crucial area.
Keywords:
- Salonga National Park,
- anthropogenic changes,
- deforestation,
- remote sensing/GIS,
- landscapes approaches
Article body
Introduction
Les forêts tropicales, autrefois couvrant 12 % de la surface terrestre, occupent moins de 6 % depuis le début du 21e siècle (Brandon, 2014). Malgré cette réduction, elles hébergent environ les deux tiers de la biodiversité mondiale et procurent des avantages essentiels au bien-être humain à tous les niveaux — local, régional et global (Gardner et al., 2009). Cependant, les activités humaines ont entraîné une déforestation et une dégradation des forêts, provoquant un taux d'extinction des espèces 100 fois supérieur au rythme naturel au cours du siècle dernier (Ceballos et al., 2015). Ainsi, les aires protégées sont cruciales pour la conservation de la biodiversité des forêts tropicales face à cette menace (Havyarimana et al., 2017). Pourtant, seulement environ 10 % du biome des forêts tropicales se trouve dans des zones strictement protégées (Schmitt et al., 2009). De plus, la viabilité à long terme de ces réserves est fortement influencée par les activités humaines dans les zones adjacentes (Wittemyer et al., 2008). L'avenir des espèces forestières tropicales reste donc incertain, d'autant plus que peu de régions des tropiques ont échappé aux impacts humains (Kareiva et al., 2007). En Afrique subsaharienne, les forêts du bassin du Congo qui constituent le deuxième plus grand écosystème forestier tropical après l'Amazonie, n'échappe pas à cette tendance.
En effet, ce bassin soutient les moyens de subsistance de 60 millions de personnes qui y vivent ou résident à proximité (de Wasseige, 2014). Cependant, les activités humaines dans cette région entraînent une déforestation représentant 0,15 % de la surface forestière du bassin du Congo, contre 0,51 % en Amérique tropicale et 0,58 % en Asie tropicale (Bellassen et Gitz, 2008). En République Démocratique du Congo (RDC), qui possède environ 152 millions d’hectares de forêts (60 % du territoire), ces forêts sont capables de stocker 140 Gt (Gigatonne) de CO2 (Potapov et al., 2012). Cependant, elles sont menacées principalement par l’agriculture sur brûlis et l’exploitation pour le bois de chauffe (Misser, 2013 ; Useni et al., 2017).
Pour conserver sa riche biodiversité, la RDC a établi un réseau de 51 aires protégées couvrant environ 11 % du territoire national (Pélissier et al., 2015). Ces aires protégées font face à des problèmes récurrents de gestion et de durabilité des ressources (Doumenge et al., 2015). De nombreuses études révèlent que les menaces sur les aires protégées sont principalement dues aux activités anthropiques (Kyale et al., 2019 ; Useni et al., 2020 ; Mangaza et al., 2022). Les pratiques agricoles non durables, l’exploitation forestière industrielle et artisanale, ainsi que la chasse, sont identifiées comme les principaux moteurs de la déforestation en RDC. Ces transformations se manifestent par des changements dans la structure du paysage et une perte progressive de biodiversité (Kalambay et al., 2016).
La pression sur la diversité biologique est particulièrement forte dans les zones à populations pauvres. Le défi de la gestion durable des aires protégées est lié à la compréhension de la dynamique des paysages, afin de trouver un compromis entre la préservation de l’environnement et la survie des communautés riveraines (Potapov et al., 2012 ; Wabasa, 2017 ; Useni et al., 2020). Cette question est cruciale pour le Parc national de la Salonga (PNS) en RDC, le plus grand parc forestier d’Afrique (36 000 km²).
Des études antérieures ont examiné l'impact des activités humaines dans le PNS sur les ressources naturelles via des enquêtes ou inventaires (WCS, 2014 ; Wabasa, 2017 ; Tingu et al., 2019). Cependant, peu ont abordé les changements dans la dynamique paysagère des forêts primaires du PNS qui sont des écosystèmes naturels préservés de toute perturbation significative d'origine humaine depuis plusieurs siècles, favorisant l'émergence d'une biodiversité mature et stable. Les avancées en imagerie satellite et en apprentissage automatique facilitent désormais cette analyse, permettant une étude plus précise des changements environnementaux au fil du temps (Crowley et Cardille, 2020). Bien que cette approche ait été utilisée dans diverses aires protégées en RDC (Kalambay et al., 2016 ; Kisangala et al., 2019 ; Kyale et al., 2019 ; Useni et al., 2020) et en Afrique (Soulama et al., 2015), son application dans la Salonga a été entravée par des nuages, nécessitant des corrections d'images significatives. Malgré ces défis, de telles analyses sont cruciales pour mieux comprendre l'impact des activités humaines dans la région et orienter les décisions de gestion à long terme des ressources naturelles.
Cette étude vise à quantifier la dynamique spatio-temporelle du paysage dans le bloc sud du PNS entre 2002 et 2020, et à analyser l'impact des villages sur cette évolution. Cet objectif permet de tester l'hypothèse selon laquelle le développement des activités agricoles dans le PNS entraîne une modification de la structure spatiale par la régression de la couverture de forêt primaire. De plus, l'étude vérifie si la présence des villages contribue à cette régression, avec une diminution progressive de la disparition de la forêt primaire à mesure que l'on s'éloigne des zones habitées.
Méthodologie
Site d’étude
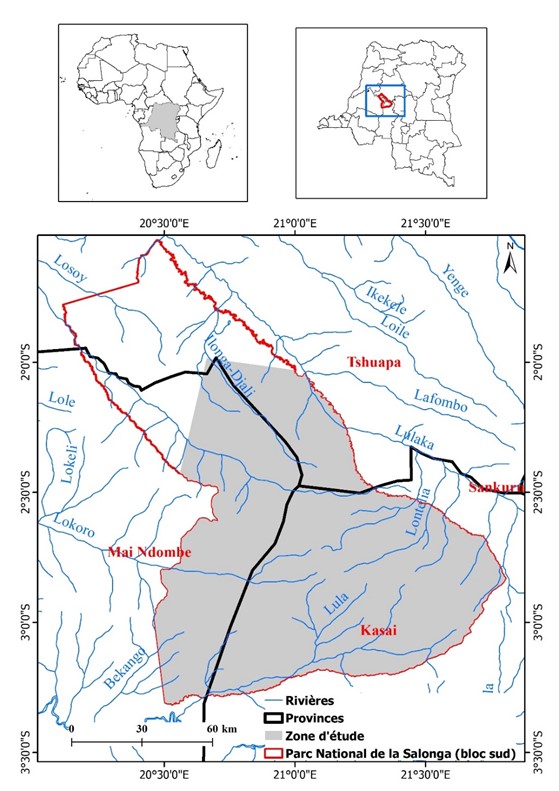
Le Parc national de la Salonga (PNS), créé en 1970, est situé entre les latitudes 1° et 3°30' Sud et les longitudes 20° et 23° Est (figure 1). Il est constitué de deux blocs de grandes forêts ombrophiles et s’étend sur 36 000 km2 (Van Krunkelsven et Draulans, 2000). Ce parc se localise dans les provinces du Sankuru, de Maindombe, du Kasaï et de la Tshuapa. On trouve à l’intérieur neuf villages dont un dans le bloc nord (Kinki Botonganji) et huit dans le bloc sud (Iyamba, Luapa, Ila, Nsese, Ediki, Bosanja, Isambo Botongola and Bokumu Beke (Ibesoa, 2008).
La majeure partie du parc est occupée par de basses terres inondables et traversée au nord par les rivières Lomela et Salonga, qui se jettent dans la Ruki, affluent du fleuve Congo. Au sud, les principales rivières sont la Lukenie et le Sankuru, affluents du Kasaï, qui se jettent dans le fleuve Congo à Kwamouth. Les précipitations mensuelles, de l’ordre de 2100 mm au nord et de 1700 mm au sud, sont bien réparties tout au long de l’année, avec une diminution légère entre juin et août (Reinartz et al., 2006). La forêt semi-décidue constitue le type de forêt prédominant sur les terres sèches du PNS, dominées par les peuplements de Scorodophloeus zenkeri et de Gilbertiodendron dewevrei. Les forêts secondaires de tous âges y sont moins fréquentes (Van Krunkelsven et Draulans, 2000).
Le parc regorge d’importantes espèces animales endémiques : Afropavo congensis, Pan paniscus, Loxodonta cyclotis et Mecistops cataphractus. La population riveraine du PNS, de plus en plus croissante, vit dans 700 villages qui ont des populations comprises entre 300 et 500 habitants (Wabasa, 2017). La pratique de l’agriculture sur brûlis, la pêche illicite dans les cours d’eau traversant le parc, la chasse de la faune pour la consommation familiale et la commercialisation, la recherche de bois d’énergie pour la préparation des mets constituent les activités principales réalisées dans le PNS (WCS, 2014 ; Tingu et al., 2019). Ces activités mettent en péril les services écologiques, sociaux et économiques du parc à l'étude (UICN – PACO, 2010).
Figure 1
Localisation du Bloc Sud du PNS
Méthodes
Données utilisées
La zone d’étude se situe dans une région tropicale humide où il pleut toute l’année. Cependant, entre janvier et mars, il y a des périodes de courte sécheresse. C’est pendant cette période que nous avons sélectionné nos données. Pour détecter le changement de l’occupation du sol, le choix des images satellitaires conditionne le succès des résultats à obtenir (Lagabrielle et al. , 2005). La dynamique paysagère dans le bloc sud du PNS a été appréciée à partir de l’analyse d’une image Landsat 5 (TM) de 2002, une image Landsat 7 (ETM+) de 2007 et deux images Landsat 8 (OLI)[1] de 2014 et 2020 dont les caractéristiques sont présentées dans le tableau 1. Les quatre images, de 30 mètres de résolution, ont été acquises entre les mois de janvier et mars, pendant les périodes les moins pluvieuses pour chaque année considérée dans cette étude afin de mettre en évidence les différences spectrales plus grandes dans les classes d’occupation du sol et de minimiser l’impact des nuages (Conghe et al., 2001). Ces images ont été téléchargées à partir du site de United States Geological Survey (USGS)[2].
Tableau 1
Caractéristiques des images Landsat TM, ETM + et OLI

λ est la longueur d'onde (Farah et al. , 2021).
Traitement des données
Pour améliorer la qualité des images Landsat acquises, les corrections radiométriques ont été effectuées à l’aide du Fast Line-of-Sight Atmospheric Analysis of Spectral Hypercubes (FLAASH). Ce module avancé de correction atmosphérique utilise l'algorithme de transfert radiatif MODTRAN4, basé sur l'équation standard de radiance spectrale pour chaque pixel du capteur (Anderson et al., 2002). Il s'applique à la gamme de longueurs d'onde du rayonnement solaire, en excluant l'émission thermique, et suppose une surface lambertienne et plane ou ses équivalents. Ce module intègre les effets du rayonnement réfléchi par la surface terrestre et diffusé par l'atmosphère jusqu'au capteur, offrant ainsi une correction précise des influences atmosphériques sur les données spectrales. La différence entre ces deux radiances résulte de l'effet de contiguïté, un mélange spatial de radiance entre pixels adjacents causé par la diffusion atmosphérique (López-Serrano et al., 2016). Cette correction radiométrique a été réalisée à l’aide du logiciel ENVI 5.3.
De plus, concernant les corrections géométriques, les images Landsat de 2002, 2007 et 2014 ont ensuite été soumises à un processus d'ortho-rectification en référence à l'image Landsat de 2020 afin de corriger les distorsions géométriques inhérentes aux images satellitaires, permettant ainsi l'obtention d'images géoréférencées avec une précision accrue (Saito et Miura, 2019). Le processus d'ortho-rectification a nécessité l'application de l'algorithme Random Sample Consensus (RANSAC) pour identifier les points de contrôle correspondants entre l'image Landsat et les données de terrain, le calcul des transformations nécessaires, l'application de ces transformations, la correction des effets topographiques, la validation du processus, et enfin la génération de l'image ortho-rectifiée (Lee et Yum, 2019).
Traitement des images
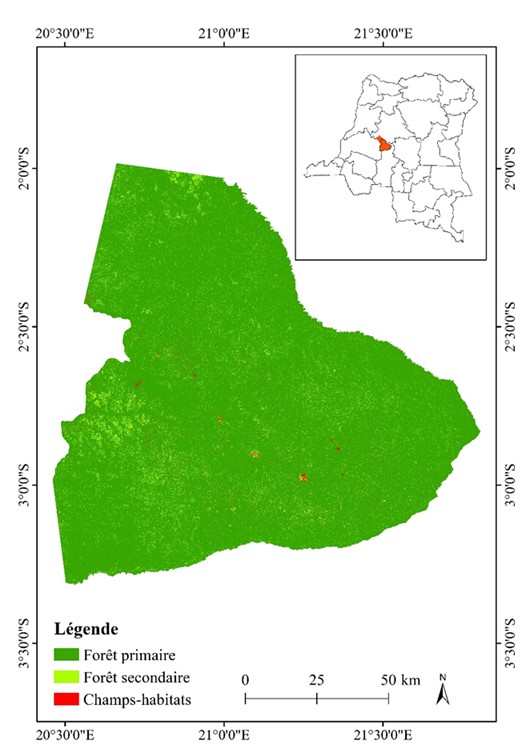
En vue de réaliser la classification des images Landsat, des compositions colorées fausses couleurs ont été effectuées pour une analyse plus efficace et une interprétation visuelle plus aisée des résultats (Phan et al., 2020). Pour les images Landsat TM et ETM+, la composition colorée a été réalisée en associant les bandes proches infra-rouge (PIR), rouge (R), et vert (V) (Caloz et Collet, 2001), tandis que pour l'image Landsat OLI, les bandes moyen infra-rouge (MIR), proche infra-rouge (PIR), et rouge (R) ont été combinées (Issiaka et al., 2016). Les canaux proches infra-rouge et rouge ont été préférés en raison de leur capacité à discriminer efficacement la végétation (Useni et al., 2020). Pour une visualisation optimale, un ajustement des étendues de couleur et des contrastes a été réalisé. Par la suite, trois classes d'occupation du sol dominantes dans le PNS ont été retenues, à savoir la forêt primaire, la forêt secondaire, les champs-habitats (figure 2, tableau 2). Les zones d'entraînement des différentes classes d’occupation du sol ont été déterminées lors des travaux de terrain réalisés entre janvier et février 2020. Pour les zones inaccessibles du domaine de chasse, des images à haute résolution obtenues via des images d'archives de Google Earth Pro ont été utilisées (Phan et al., 2020). Ces zones d’entraînement ont été collectées à l’aide d’un GPS de type Garmin 64 st (précision de plus ou moins 3 mètres).
Ces zones d'entraînement ont été ensuite représentées sous forme de points sur l'image composite, se répartissant en 350 pour la forêt primaire, 250 pour la forêt secondaire, 150 pour les champs-habitats indépendamment sur toutes les images. Ces aires d'entraînement ont été utilisées pour effectuer une classification supervisée en utilisant l'algorithme du maximum de vraisemblance (Caloz et Collet, 2001). Cet algorithme est crucial, car il offre une méthode statistiquement solide pour assigner les pixels aux classes d’occupation du sol en se fondant sur la distribution de probabilité des données d'entraînement, ce qui augmente la précision et la fiabilité des résultats de classification (Otukei et Blaschke, 2010). Ces analyses ont été réalisées sous ArcGIS 10.8.1.
Tableau 2
Description des classes d'occupation du sol

Figure 2
Localisation des classes d'occupation du sol du bloc sud du parc national de la Salonga
Validation de la classification
Après les travaux de classification, des missions de terrain ont été effectuées dans la zone d’étude, leurs validations ayant été effectuées du 15 juillet au 20 août 2020. Ensuite, ces points ont été utilisés pour vérifier la qualité des classifications des images Landsat. Cette qualité des classifications a été évaluée à travers la précision globale, qui caractérise la proportion de pixels correctement classés, et le coefficient Kappa, qui correspond au rapport entre les pixels correctement classés et l'ensemble des pixels considérés (Hubert-Moy et al. , 2001). Cet indice a été critiqué par Pontius et Millones (2011) en raison de sa tendance à surestimer l'accord au hasard, pouvant conduire à une sous-estimation de la précision de la classification. Néanmoins, son utilisation dans cette étude a été motivée par la capacité à tester les différences significatives entre deux coefficients indépendants, le positionnant ainsi comme une mesure essentielle d'évaluation de la précision (Congalton et Green, 2009). Sa valeur varie généralement entre 0 et 1 (Lillesand et al. , 2008). Un coefficient Kappa supérieur à 0,80 indique un accord fort (presque parfait), une valeur entre 0,40 et 0,80 indique un accord modéré à substantiel, tandis qu'une valeur inférieure à 0,40 indique un accord faible à médiocre (Congalton et Green, 2009). Cette analyse a été réalisée sous le logiciel ArcGIS 10.8.1., qui a été également utilisé pour la mise en page cartographique.
Analyse de la dynamique paysagère
Trois indices de structure spatiale, qui renseignent sur la fragmentation du paysage entre deux dates (Barima et al., 2009), ont été calculés pour déterminer les rapports existants entre la structure paysagère et les processus écologiques de manière quantifiable. Il s’agit du nombre de taches, de l’aire totale des taches et de l’indice de perturbation défini comme le rapport de l’aire cumulative des classes anthropiques dans le paysage et de l’aire cumulative des classes naturelles. La quantification du changement spatio-temporel intervenu entre classes d’occupation du sol a été réalisée à l’aide de la matrice de transition (Mama et al., 2013). Le rapport entre la somme des valeurs en dehors de la diagonale et la valeur sur la diagonale de la matrice de transition, a été utilisé pour exprimer les dynamiques de conversion de chaque type de classe d’occupation du sol (Bogaert et al., 2014), par le moyen de leur stabilité. Enfin, à partir de l’aire et du nombre de taches, les processus de transformation spatiale ont été identifiés à l’aide d’un arbre de décision de Bogaert et al. (2004). La différenciation entre la dissection et la fragmentation a été effectuée en utilisant la valeur t théorique = 0.75, où t calculé (le rapport entre l’aire finale et l’aire initiale) ≤ 0.75 indique la fragmentation et t > 0.75 suggère la dissection (de Haulleville et al., 2018).
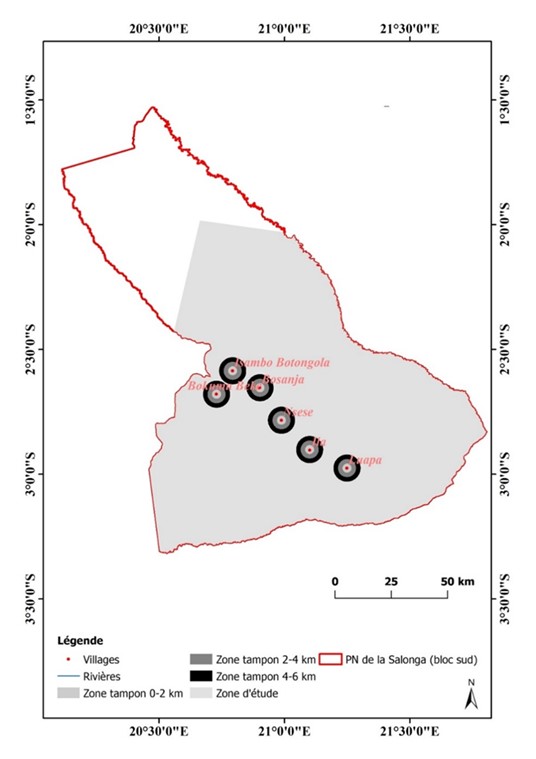
Afin de quantifier le changement de composition du paysage autour de six villages se trouvant au sein du PNS, la proportion de la forêt primaire et de la forêt secondaire a été calculée dans les zones tampons comprises entre 0-2 km, 2-4 km et 4-6 km du centre géométrique (centroïde) des villages retenus (Useni et al., 2018 ; Useni et al., 2020b). Ces rayons couvrent des distances proches (0-2 km), moyennement éloignées (2-4 km) et éloignées (4-6 km) des villages se trouvant au sein du parc (figure 3). Le rayon maximal de 6 km a été fixé aléatoirement pour déterminer l’impact des villages sur le paysage forestier. Le test de Kolmogorov-Smirnov a été utilisé pour évaluer l’ajustement de données à la loi normale (Steinskog et al., 2007). Ensuite une transformation Box-Cox a été réalisée pour normaliser les données afin d’utiliser l’analyse de la variance (ANOVA) pour évaluer l’effet année et l’effet de la distance sur la proportion des forêts primaires et des forêts secondaires (Khoji et al., 2022). Ces analyses statistiques ont été réalisées sous le logiciel R, avec une valeur seuil de probabilité de 5%.
Figure 3
Différentes zones tampons selon la distance autour des villages
Résultats
Précision des classifications
Le tableau 3 présente les matrices de confusion. Il en résulte des valeurs de précision globale variant entre 84,0% et 91,5%, et celles du coefficient Kappa entre 0,79 et 0,89. En général, ces résultats suggèrent une bonne discrimination entre les différentes classes d'occupation du sol, ce qui les rend appropriés pour une analyse. Cependant, des confusions subsistent : la matrice de confusion a montré que 6%, 2%, 9%, et 3% des pixels de forêt secondaire ont été confondus avec la forêt primaire en 2002, 2007, 2014, et 2020 respectivement. De plus, 10%, 11%, 8%, et 11% des pixels de forêt primaire ont été confondus avec la forêt secondaire sur ces mêmes périodes. Des confusions ont également été observées entre les forêts secondaires et les champs-habitats. Cette confusion peut être attribuée d’une part à la similitude des signatures spectrales et des structures forestières entre les forêts secondaires matures et les forêts primaires. D'autre part, les similitudes entre les cultures et les forêts secondaires qui sont encore jeunes contribuent également à ces erreurs de classification.
Tableau 3
Précision des classifications supervisées des images Landsat de 2002, 2007, 2014 et 2020 appuyée sur l'algorithme du maximum de vraisemblance

Les valeurs des colonnes réfèrent aux données classifiées et celles des lignes aux données de référence (terrain). Les paramètres de précisions sont en pourcentage (%).
Évolution paysagère et conversion des classes d’occupation du sol dans le bloc sud du PNS
Les proportions des classes d’occupation du sol du paysage du bloc sud du PNS permettent de constater des évolutions différentes durant la période de 2002 à 2020. Les états de l’occupation du sol (figure 4) permettent d’observer que la forêt secondaire et les champs-habitats sont caractérisés par une dynamique progressive, notamment vers le centre sud, contrairement à la forêt primaire dans le paysage du PNS, quoique dominante, qui affiche une dynamique régressive.
En 18 ans, la matrice paysagère reste dominée par les forêts primaires, quoique progressivement dégradées en forêts secondaires et en champs-habitats (tableau 4). En effet, il est observé sur cette période, une perte totale de 10,8% (soit annuellement 0,6%) des superficies de forêts primaires, dont 0,5% entre 2002 et 2007 ; 3,7% entre 2007 et 2014 et 6,9% entre 2014 et 2020. Ainsi, le taux annuel de perte de forêt primaire est de 0,09% entre 2002 et 2007 ; 0,5% entre 2007 et 2014, et 0,9% entre 2014 et 2020, correspondant respectivement à 62,85 km 2 , 463,41 km 2 et 843,68km 2 des forêts). On assiste à un gain de 13,5% des forêts secondaires et 9,3% des champs-habitats par an. Le taux annuel de déforestation (perte de forêts primaires et secondaires confondues) est évalué à 0,006%.
Très peu de changement est observé entre la période de 2002 à 2007, caractérisée par la conversion de 0,40 % et 0,08 % des forêts primaires, respectivement en forêts secondaires et champs-habitats. Sur la période de 2007 à 2014, il a été noté 3,52 % de superficies de forêts primaires converties en forêts secondaires ; ces dernières ont été converties à hauteur de 0,04% en champs-habitats. Au fil du temps, entre 2014 et 2020, il a été observé une perte nette de 6,42% des forêts primaires, converties en forêts secondaires à 6,02 % et 0,40 % en champs-habitats. Bien plus, 0,77 % des forêts secondaires se sont dégradées au profit des champs-habitats, durant cette même période.
L’analyse de l’indice de stabilité montre des valeurs élevées entre 2002-2007 caractéristiques d’une stabilité des classes dans le paysage alors que les périodes 2007-2014 et 2014-2020 montrent des valeurs faibles indiquant une instabilité des classes, synonyme de l’impact humain sur le paysage (tableau 4).
Figure 4
Cartes de l’occupation du sol dans le bloc sud du PNS en 2002, 2007, 2014 et 2020
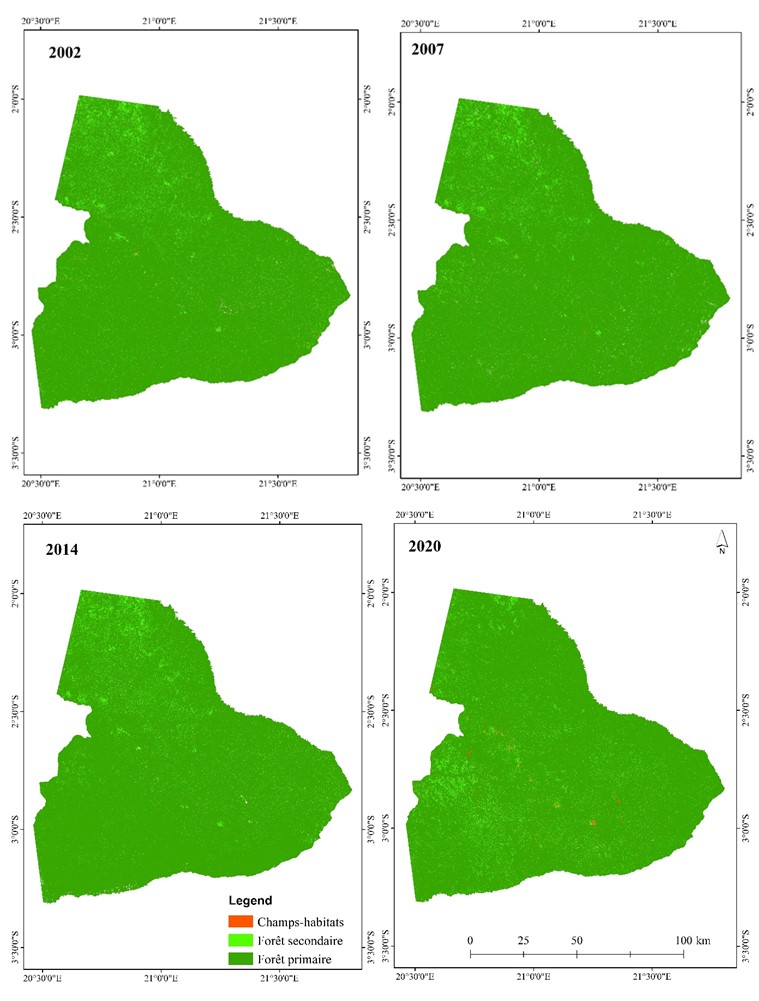
Tableau 4
Matrices de transition (en pourcentage) des classes d’occupation du sol issue de la classification des images Landsat de 2002, 2007, 2014 et 2020

Images Landsat de 2002, 2007, 2014 et 2020 observées au sein du Parc National de la Salonga. 1%= 131,4 km2.
Dynamique structurale du paysage du PNS
Entre les périodes 2002-2007 et 2014-2020, la suppression a été le processus caractéristique des forêts primaires, car il est noté une diminution simultanée de l’aire totale et du nombre de taches (tableau 5). En revanche entre 2007 et 2014, la dissection a été le processus de transformation spatiale dominant des forêts primaires d’autant plus que la diminution de l’aire totale des taches est accompagnée de l’augmentation du nombre de taches (tableau 4). Par ailleurs, durant les périodes 2002-2007, 2007-2014 et 2014-2020, le processus de transformation spatiale des forêts secondaires et des champs-habitats a été la création, étant donné qu’il est observé l’augmentation du nombre de taches et de leur aire totale de façon simultanée (tableau 4).
Entre 2002 et 2020, il a été noté une multiplication par près de 12 des valeurs de l'indice de perturbation , qui sont ainsi passées de 0,0014 en 2002 à 0,017 en 2020, traduisant ainsi l’importance du phénomène de dégradation du milieu et donc de l’anthropisation des classes d’occupation naturelle.
Tableau 5
Valeurs du nombre de taches (n) et de l’aire totale (at) en km2 par année et par classe d’occupation du sol dans le bloc Sud du Parc National de la Salonga en 2002, 2007, 2014 et 2020

Les processus de transformation spatiale (PTS) ont été identifiés sur la base de l’arbre de décision (12).
Dynamique de l’occupation du sol autour des villages ciblés dans la partie sud du PNS
L’analyse de la variance montre une différence significative au niveau de la proportion des classes d’occupations du sol autour des villages, entre rayons (p<0,01). Au cours des quatre dates retenues, quel que soit le village (figure 5), la proportion de la forêt primaire a diminué (p<0,05), passant en moyenne de 97,2% en 2002 à 74,0% en 2020 dans la zone tampon comprise entre 0 et 2 km ; de 99% à 92% dans la zone tampon comprise entre 2 et 4 km et de 99,2% à 94,0% dans la zone tampon comprise entre 4 à 6 km (p<0,001). Pendant la même période, la proportion de la forêt secondaire a augmenté (p<0,01), passant de 2,3% à 14,2% dans la zone tampon comprise entre 0 et 2 km ; 0,7% à 5,5 % dans celle comprise entre 2 et 4 km et de 0,7 % à 5,2% dans la zone tampon comprise entre 4 et 6 km (p<0,05). Globalement, la proportion de la forêt secondaire a, quant à elle, augmenté, particulièrement entre 2007 et 2014. De plus, le complexe champs-habitats a augmenté autour des villages (p<0,05) entre 2002 et 2020 tout en diminuant en s’éloignant des villages (p<0,05), passant de 0,37% à 11,87% entre 0 et 2 km, 0,11% à 1,64% entre 2 et 4 km, et 0,02% à 0,61% entre 4 et 6 km. Globalement, ces résultats suggèrent une transformation de l'occupation du sol autour des villages au fil du temps, caractérisée par une diminution de la forêt primaire puis une augmentation de la forêt secondaire et du complexe champs-habitats.
Figure 5
Proportion des forêts primaires et secondaires ainsi que le complexe champs-habitats

Figure

Évolution de la proportion de la forêt primaire (en haut) et de la forêt secondaire (au milieu) et du complexe champs-habitats à différentes distances par rapport aux villages villages (Iyamba, Ila, Nsese, Bosanja, Isambo Botongola, Bokumu Beke) entre 2000 et 2020 sur base de la classification des images Landsat de 2002, 2007, 2014 et 2020. La zone tampon comprise entre 0 et 2 km a une surface de 12,5 km2, celle comprise entre 2 et 4 km a une surface de 37,6 km2 et celle comprise entre 4 et 6 km une surface de 62,7 km2.
Discussion
Approche méthodologique
L'utilisation des images Landsat de 2002 à 2020 a permis de quantifier la dynamique des forêts primaires du bloc sud du PNS. Ces images, largement utilisées en Afrique pour cartographier la dynamique spatio-temporelle des paysages forestiers et urbains, ont répondu à l'objectif de l'étude (Diallo et al., 2011 ; ICCN, 2016). La classification des images satellitaires a été soutenue par la connaissance des observations sur le terrain, réduisant la confusion entre pixels proches thématiquement (Foody, 2010). Cependant, certaines confusions ont été observées entre les classes d’occupation du sol, sans affecter négativement les tendances générales de l’évolution de l'occupation du sol. L'évaluation de la composition des paysages autour des villages a utilisé une approche basée sur des zones tampons, efficace dans plusieurs études (Bamba et al., 2010 ; Havyarimana et al., 2017). Bien que l'utilisation de multiples indices de structure spatiale soit généralement requise, seuls l'aire et le nombre de taches ont été calculés, pertinents pour mettre en évidence la fragmentation des paysages (Bogaert et Mahamane, 2005). Il est à noter que plusieurs indices de structure spatiale sont corrélés, rendant le recours à plusieurs indices parfois non optimal en raison de la redondance créée (Bogaert et al., 2002).
Dynamique paysagère des forêts primaires du bloc sud du PNS en RD Congo
Entre 2002 et 2020, les forêts primaires du bloc sud du PNS ont régressé, perdant respectivement 0,5%, 3,7%, et 6,9% de leur superficie aux intervalles de 2002-2007, 2007-2014 et 2014-2020. Simultanément, les forêts secondaires ont augmenté, avec une expansion des champs-habitats. En combinant les forêts primaires et secondaires, le taux de déforestation annuel est de 0,006%, bien en deçà de la moyenne nationale en RDC (0,18 à 0,46%) (de Wasseige et al., 2012 ; Cabala et al., 2022). Comparativement, ce taux est inférieur à celui observé dans d'autres régions comme la réserve de biosphère de Yangambi (Kyale et al., 2019) et la forêt néphéliphile du mont Oku au Cameroun (Momo et al., 2012), attribuable à des différences d'échelle temporelle et d'intensité de pression anthropique.
Bien que le taux de déforestation global soit bas, les forêts primaires ont régressé rapidement, particulièrement de 2014 à 2020, avec une perte annuelle de 0,6%, dépassant la moyenne nationale (0,2-0,3%) et la moyenne sous-régionale (0,15% en 2008 dans le bassin du Congo). Cette accélération est attribuable à des différences méthodologiques et d'échelles temporelles (Useni et al., 2017). Les populations locales, en constante croissance, malgré la présence d'ONG proposant des alternatives économiques, cherchent à subsister sans se préoccuper de la durabilité des ressources dans un contexte économique précaire (Vermeulen et al., 2011).
La diminution de la couverture des forêts primaires observée dans cette étude confirme les tendances similaires signalées dans d'autres régions, notamment la forêt de Mayombe avec le parc marin de Mangrove dans le Kongo Central (Kisangala et al., 2019). Elle reflète également les tendances générales des forêts congolaises, telles que la régression de la forêt de Miombo dans l'Arc cuprifère katangais, la plaine de Lubumbashi, et la réserve de biosphère de Lufira (Bamba et al., 2010 ; Cabala et al., 2018 ; Useni et al., 2020 ; Cabala et al., 2022 ; Mpanda et al., 2022).
Dans le bloc sud du PNS, l'exploitation des ressources naturelles, telles que le bois, le gibier et les terres agricoles non appropriées, incite les riverains à s'installer de manière permanente, malgré l'opposition de la législation congolaise sur la gestion des parcs nationaux (Kalambay et al., 2016). Les Yaelima revendiquent des droits de résidence et de propriété, motivés par des besoins de subsistance tels que l'habitat, l'agriculture, la pêche et la chasse (Tingu et al., 2019). Cependant, cette présence humaine, associée à des activités telles que l'agriculture sur brûlis, la pêche illicite, la chasse de faune à des fins familiales et commerciales, ainsi que la recherche de bois pour la préparation alimentaire, menace les services écologiques, sociaux et économiques du parc (UICN – PACO, 2010 ; WCS, 2014). Ces activités de subsistance entraînent la dégradation des forêts primaires et la déforestation à proximité des zones d'activités intensives. Les ressources naturelles dans le bloc sud du Parc national de la Salonga sont en danger, mettant en péril des espèces rares, malgré les efforts de conservation déployés par des organisations nationales et internationales pour atténuer la pression sur les ressources naturelles en RDC (UICN – PACO, 2010). Des tendances similaires ont été observées au Cameroun, un autre pays du bassin du Congo, où la croissance démographique continue de menacer les écosystèmes forestiers, comme dans la forêt néphéliphile du mont Oku (Momo et al., 2012).
La situation socio-économique dans la zone d'étude montre une faible diversification économique, la paupérisation de la population, et la diminution de la main-d'œuvre locale après la fermeture d'entreprises clés, notamment ENTRIAC et SOCOPLAN. Les populations en périphérie, parfois dans le parc, voient ce dernier comme la seule source de survie, exerçant une pression significative, notamment à travers l'agriculture itinérante sur brûlis (Kalambay et al., 2016). Malgré la préservation de certains arbres utiles, la destruction totale d'autres et les incendies réguliers entravent la reconstitution des forêts primaires (Diallo et al. , 2011). Les feux, en particulier avec une quantité de combustibles élevée, réduisent la floraison, la fructification et le pourcentage de germination, entraînant la mortalité des plantules (Alvarado, 2012).
L'avènement du code forestier en 2002 a attiré des partenaires pour soutenir l'Institut congolais pour la conservation de la nature (ICCN) dans la gestion du PNS. Avec des pratiques de chasse de plus en plus restrictives, la majorité des chasseurs sont devenus agriculteurs, intensifiant la pression sur la forêt primaire, surtout après 2007. La mise en culture continue des sols forestiers a diminué le taux de matière organique, réduisant la fertilité du sol (Useni et al. , 2020). Les cultures vivrières principales des populations riveraines exigent une grande fertilité des sols, conduisant à considérer les forêts comme les zones les plus favorables à l'agriculture. Les champs responsables de la déforestation se situent le long des routes et autour des villages, facilitant l'activité des paysans.
L'indice de perturbation (U), multiplié par près de 12 en 18 ans, signale une perturbation croissante des écosystèmes naturels dans le paysage (Keith et al., 2009). La nette progression de la valeur de U confirme l'impact croissant des activités humaines sur la perturbation des forêts du PNS (Mama et al., 2013), soulignant que la zone protégée n'est pas encore entièrement délimitée matériellement. Un constat similaire au Burundi, où l'absence de délimitation matérielle de la réserve naturelle forestière de Bururi (RNFB) et le manque de contrôle ont entraîné l'installation des populations riveraines dans la réserve, provoquant la déforestation (Havyarimana et al., 2017). Les paysages du PNS subissent une dénaturation progressive et radicale sous l'effet des perturbations anthropiques, comme la déforestation et l'agriculture, perdant ainsi leur évolution naturelle.
Les populations vivant dans les villages du Parc national de la Salonga sont en constante croissance, ce qui accentue la pression sur les ressources forestières et conduit à leur dégradation. Cette situation est exacerbée par des problèmes récurrents de gouvernance des ressources naturelles dans le pays (Polepole, 2013). Bien que la gestion du parc implique la participation des communautés locales, les politiques de gestion s'avèrent globalement inefficaces, et un écart considérable persiste entre les objectifs déclarés et les résultats obtenus (Kambala, 2023).
Implication pour la gestion durable du parc
Le paysage, un ensemble d’écosystèmes en interaction, reflète la dynamique des conditions écologiques et des pratiques socio-économiques. Dans le PNS, une dégradation rapide, matérialisée par la conversion des forêts primaires en forêts secondaires ou en champs-habitat autour des villages, est observée. Bien que le taux de déforestation soit relativement faible dans le bloc sud du PNS, ces écosystèmes subissent depuis des décennies des perturbations majeures liées à l'agriculture, l'élevage, l'exploitation du charbon de bois, la recherche de bois de feu et les feux de végétation (Vermeulen et al., 2011 ; Tingu et al., 2019). Les forêts tropicales, riches en espèces et importantes pour la conservation du carbone fournissent des services écosystémiques cruciaux (Keith et al., 2009 ; Mangaza et al., 2022). Leur conversion en forêts secondaires diminue ces capacités, y compris pour atténuer les effets du changement climatique (Baccini et al., 2017). Face à la dépendance croissante des populations aux ressources forestières, des modèles économiques durables sont nécessaires pour diversifier les revenus au-delà de l'agriculture itinérante sur brûlis. La prise en compte des intérêts socio-économiques des communautés locales, couplée à une meilleure sécurisation des réserves forestières, peut favoriser la reconstitution du couvert forestier, comme observé au Burundi (Havyarimana et al., 2017). Des politiques axées sur l'aménagement des zones impactées sont cruciales pour préserver les forêts primaires en tant que poumons essentiels de la planète. Ces politiques devraient s'appuyer sur la restauration forestière, l'aménagement des forêts secondaires et la réhabilitation des terres fortement dégradées.
Conclusion
La présente recherche a mis en exergue la pertinence de l’approche cartographique couplée aux outils d’analyse de l’écologie du paysage à l’évaluation de la dynamique paysagère des forêts primaires du PNS en RDC. Les résultats révèlent une régression de la surface de forêts primaires à travers une dynamique de morcellement et suppression des taches, parallèlement à une augmentation du nombre de taches et de la surface totale des forêts secondaires et champs-habitats. Bien que la forêt primaire demeure dominante dans la matrice paysagère, elle subit des pertes annuelles de l'ordre de 0,61%, principalement en faveur des forêts secondaires et des champs-habitats, indiquant une tendance générale à la dégradation du paysage, particulièrement marquée autour des villages. Ces résultats soulignent la nécessité pour les gestionnaires du PNS d'intégrer, dans les programmes de conservation, des modèles économiques et de développement favorisant la diversification des sources de revenus, tout en planifiant de manière à préserver les différentes fonctions du paysage.
Appendices
Remerciements
Nos remerciements à l’Académie de recherche et d’enseignement supérieur (ARES) pour le financement des missions de terrain, à travers le projet de recherche pour le développement « Renforcement des capacités de gestion durable de la forêt claire de miombo par l’évaluation de l’impact environnemental de la production de charbon de bois et l’amélioration des pratiques vis-à-vis des ressources forestières (CHARLU, Charbon de Lubumbashi) et le programme d’appui institutionnel avec l’Université de Kinshasa (UNIKIN). Les gestionnaires du Parc national de la Salonga sont remerciés pour les facilités accordées sur les sites.
Notes
-
[1]
TM : Thematic Mapper ; EMT+ : Enhanced Thematic Mapper Plus ; OLI : Operational Land Imager.
-
[2]
Pour consulter le site United States Geological Survey (USGS), consulter le lien suivant [En ligne], URL : https://earthexplorer.usgs.gov/
Bibliographie
- Alvarado, S., 2012, Évaluation du rôle des feux de brousse sur la composition, la structure et la phénologie, et la résistance de la végétation des bois de tapia (Uapaca bojeri) du massif d’Inoty, Nouvelle Aire Protégée, en vue de sa gestion durable, Thèse de doctorat, Université d’Avignon et des Pays de Vaucluse, France, et l’Université d’Antananarivo, Madagascar, 223 p.
- Anderson, G. P., Felde G.W., Hoke M.L., Ratkowski A.J., Cooley T.W., Chetwynd J.H., Jr., Gardner J.A., Adler-Golden S.M., Matthew M.W., Berk A., Bernstein L.S., Acharya P.K., Miller D.P., et Lewis P.E., 2002, MODTRAN4-based atmospheric correction algorithm: FLAASH (fast line-of-sight atmospheric analysis of spectral hypercubes), dans Shen, S.S., Lewis, P.E. (dir.), Algorithms and Technologies for Multispectral, Hyperspectral, and Ultraspectral Imagery VIII (Proceedings of SPIE) , Society of Photo Optics, Orlando, USA, pp. 65–71.
- Baccini, A., Walker W., Carvalho L., Farina M., Sulla-Menashe D., et Houghton R. A., 2017, Tropical Forests Are a Net Carbon Source Based on Aboveground Measurements of Gain and Loss, Science , 358, 6360, pp. 230-234.
- Bamba, I., Barima Y.S.S. et Bogaert J., 2010, Influence de la densité de la population sur la structure spatiale d’un paysage forestier dans le bassin du Congo en R.D. Congo, Tropical Conservation Science, 3, 1, pp. 31-44.
- Barima, Y.S.S., Barbier N., Bamba I., Traore D., Lejoly J., Bogaert J., 2009, Dynamique paysagère en milieu de transition forêt-savane ivoirienne, Bois et Forêts des Tropiques, 299, 1, pp. 15-25.
- Bellassen, V., Gitz V., 2008, Reducing emissions from deforestation and degradation in Cameroon—assessing costs and benefits, Ecological Economics , 68 , 1-2, pp. 336-344.
- Bogaert, J., Ceulemans R., et Salvador-Van Eysenrode D., 2004, Decision tree algorithm for detection of spatial processes in landscape transformation, Environmental Management , 33, 1, pp. 62-73.
- Bogaert, J., Hecke P. V., et Ceulemans R., 2002, The Euler number as an index of spatial integrity of landscapes: evaluation and proposed improvement, Environmental Management , 29 , pp. 673-682.
- Bogaert, J., Mahamane A., 2005, Ecologie du paysage : cibler la configuration et l’échelle spatiale, Annales des Sciences Agronomiques du Bénin, 1, 7, pp. 39-68.
- Bogaert, J., Vranken I., et André M., 2014, Anthropogenic effects in landscapes: historical context and spatial pattern, dans Hong S.K., Bogaert J. et Min Q. ( dir. ), Biocultural landscapes. Diversity, functions and values , Springer, Dordrecht, The Netherlands, pp. 89-112.
- Brandon, K., 2014, Ecosystem services from tropical forests: review of current science, Center for Global Development Working Paper , 380, pp. 1-86.
- Cabala, K.S., Useni S.Y., Mwana Y.A., Munyemba K.F., et Bogaert J., 2022, Activités anthropiques et dynamique des écosystèmes forestiers dans les zones territoriales de l’Arc Cuprifère Katangais (RD Congo), Tropicultura, 40, 3-4, pp. 1-27.
- Cabala, K.S., Useni S.Y., Munyemba K.F., et Bogaert J., 2018, Activités anthropiques et dynamique spatiotemporelle de la forêt claire dans la plaine de Lubumbashi, dans Bogaert J., Colinet G., Mahy G. (dir.), Anthropisation des paysages katangais, Presses Universitaires de Liège, Liège, Belgique, pp. 253-266.
- Caloz, R., Collet C., 2001, Précis de télédétection : Traitements numériques d’images de télédétection, Presses de l’Université du Québec, Québec, 3, 386 p.
- Ceballos, G., Ehrlich P. R., Barnosky A. D., García A., Pringle R. M., et Palmer T. M., 2015, Accelerated modern human–induced species losses: Entering the sixth mass extinction, Science Advances , 1 , 5, [En ligne], URL : https://www.science.org/doi/10.1126/sciadv.1400253
- Congalton, R.G., Green K., 2009, Assessing the Accuracy of Remotely Sensed Data: Principles and Practices , seconde édition, Taylor & Francis Group, Boca Raton, 200 p.
- Conghe, S., Curtis E.W., Karen C.S., Mary Pax L., et Scott A.M., 2001, Classification and Change Detection Using Landsat TM Data: When and How to Correct Atmospheric Effects?, Remote Sensing of Environment , 75 , pp. 230–244.
- Crowley, M.A., Cardille J.A., 2020, Remote Sensing’s Recent and Future Contributions to Landscape Ecology, Current Landscape Ecology Reports , 5, pp. 45–57.
- de Haulleville, T., Rakotondrasoa O.L., Ratsimba R. H., Bastin J-F., Brostaux Y., Verheggen, F.J. Rajoelison G.L., Malaisse F., Poncelet M., Haubruge E., Beeckman H., et Bogaert, J., 2018, Fourteen years of anthropization dynamics in the Uapaca bojeri Baill. forest of Madagascar, Landscape and Ecological Engineering , 14 , pp. 135–146.
- de Wasseige, C., de Marcken P., Bayol N., Hiol H.F., Mayaux P., Desclée B., Nasi R., Billand A., Defourny P., et Ebáa, A.R., 2012, Les forêts du bassin du Congo : État des forêts 2010, Office des Publications de l’Union Européenne, Luxembourg, 276 p.
- de Wasseige, C., Flynn J., Louppe D., Hiol Hiol F., Mayaux P., 2014, Les forêts du bassin du Congo-Etat des forêts 2013, Weyrich, Neufchâteau, Belgique, 350 p.
- Diallo, H., Bamba I., Barima Y.S.S., Visser M., Ballo A., Mama A., Vranken I., Maiga M., et Bogaert J., 2011, Effets combinés des climats et des pressions anthropiques sur la dynamique évolutive de la dégradation d’une aire protégée du Mali (la Réserve de Fina, Boucle du Baoulé), Sécheresse, 22, pp. 97-107.
- Doumenge, C., Palla F., Scholte P., Hiol H.F., et Larzillière A., 2015, Aires protégées d’Afrique centrale – État 2015, OFAC, Kinshasa, République Démocratique du Congo et Yaoundé, Cameroun, 256 p.
- Farah, A., Algouti A., Algouti, A., Ifkirne M., et Rafik A., 2021, Remote sensing for spatio-temporal mapping of land surface temperature and surface energy fluxes in the Bouregreg-Chaouia Region of Morocco, Journal of Environmental and Agricultural Studies , 2, 1, pp. 23-35.
- Foody, G.M., 2010, Assessing the accuracy of land cover change with imperfect ground reference data, Remote Sensing of Environment , 114, 10, pp. 2271-2285.
- Gardner, T. A., Barlow J., Chazdon R., Ewers R. M., Harvey C. A., Peres C. A., Sodhi N. S., 2009, Prospects for tropical forest biodiversity in a human‐modified world, Ecological Letters , 12 , 6, pp. 561-582.
- Havyarimana, F., Masharabu T., Kouao J. K., Bamba I., Nduwarugira D., Bigendako M. J., Hakizimana P., Mama A., Bangirimana F., Banyankimbona G., Bogaert, J., et De cannière C., 2017, La dynamique spatiale de la forêt située dans la réserve naturelle forestière de Bururi au Burundi, Tropicultura, 35, 3, pp. 158-172.
- Hubert-Moy, L., Cotonnec A., Le Du L., Chardin A., et Perez P., 2001, A Comparison of Parametric Classification Procedures of Remotely Sensed Data Applied on Different Landscape Units, Remote Sensing of Environment , 75, pp. 174-187.
- Ibesoa, J.M., 2008, Proposal of a sustainable management of the Salonga National Park in the Democratic Republic of Congo , Thèse de doctorat, Mendel University of Agriculture and Forestry (MUAF), Brno, 127 p.
- ICCN (Institut Congolais pour la Conservation de la Nature), 2016, Plan d’Aménagement et de Gestion du Parc National de la Salonga 2016-2025, République Démocratique du Congo, 137 p.
- Issiaka N.T., Arouna O., et Imorou I.T., 2016, Cartographie De La Dynamique Spatio-Temporelle Des Parcours Naturels Des Troupeaux Transhumants Dans Les Communes De Banikoara Et De Karimama Au Benin (Afrique De L’ouest), European Scientific Journal, 12, 32, pp. 251-268.
- Kalambay, B.M., Rudant J.P., Lumbuenamo R., Beland M., et Riera, B., 2016, Dynamique spatiale du domaine de chasse et réserve de Bombo Lumene entre 2000 et 2015 par imagerie satellitaire optique, International Journal of Innovation and Applied Studies, 18, 2, pp. 559-568.
- Kambala Luadia Tshikengela, B. 2023, Analyse critique des politiques de gestion du Parc national de la Salonga en République Démocratique du Congo, Monde en développement, 50, 1, pp. 69-88.
- Kareiva, P., Watts S., McDonald R., et Boucher, T., 2007, Domesticated nature: shaping landscapes and ecosystems for human welfare, Science , 316 , 5833, pp. 1866-1869.
- Keith, H., Mackey B., et Lindenmayer D., 2009, Reevaluation of forest biomass carbon stocks and lessons from the world’s most carbon-dense forests, Proceedings of the National Academy of Sciences , 16 , 28, pp. 11635-11640.
- Khoji, M.H., N’Tambwe N.D., Malaisse F., Waselin S., Kouagou R.S., Cabala K.S., Munyemba K.F., Bastin J-F., Bogaert J., et Useni S., 2022, Quantification and simulation of landscape anthropization around the mining agglomerations of southeastern Katanga (DR Congo) between 1979 and 2090, Land , 11 , 850, pp. 1-18.
- Kisangala, M.M., Kamozi Z., et Kafuti S., 2019, Évaluation de l’état de la déforestation de la forêt du Mayombe par Télédétection de 1990 à 2015 (Province du Kongo Central-R.D. Congo), Congoscience, 7, 1, pp. 102-107.
- Kyale, K.J., Wardell D.A., Mikwa J.-F., Masimo K.J., Maindo M.N.A., Oszwald J., et Doumenge C., 2019, Dynamique de la déforestation dans la Réserve de biosphère de Yangambi (République démocratique du Congo) : variabilité spatiale et temporelle au cours des 30 dernières années, Bois et Forêts des Tropiques, 341, 3, pp. 15-28.
- Lagabrielle, E., Metzger P., Martignac C., Durieux L., et Grellet G., 2005, Guide pratique d’utilisation des informations produites dans le cadre du projet TEMOS à la Réunion, [En ligne], URL : https://www.researchgate.net/publication/237482654
- Lee, K.-H., Yum J.-M., 2019, A Review on Atmospheric Correction Technique Using Satellite Remote Sensing, Korean Journal of Remote Sensing , 35, 6, 1, pp. 1011–1030.
- Lillesand, T. M., Kiefer R. W., et Chipman, J. W., 2008, Remote Sensing and Image Interpretation , sixième edition, Wiley, New York, 720 p.
- López-Serrano, P. M., Corral-Rivas J. J., Díaz-Varela R. A., Álvarez-González J. G., et López-Sánchez C. A., 2016, Evaluation of radiometric and atmospheric correction algorithms for aboveground forest biomass estimation using Landsat 5 TM data, Remote sensing , 8, 5, 369, [En ligne], URL : https://www.mdpi.com/2072-4292/8/5/369
- Mama, A., Sinsin B., De Cannière C., et Bogaert J., 2013, Anthropisation et dynamique des paysages en zone soudanienne au nord du Bénin, Tropicultura, 31, 1, pp. 78-88.
- Mangaza, L., Makana J. R., Hubau W., Sonwa D. J., Batsi G., et Fayolle A., 2022, Impacts of land use change on biomass and diversity in the forest landscape of the Yangambi Biosphere Reserve in the Democratic Republic of Congo, Bois et Forêts des Tropiques , 353 , pp. 61-73.
- Misser, F., 2013, Les aires protégées en République Démocratique du Congo : menaces et défis, L’action de l’Union européenne, Parcs Réserves, 68, 3, pp. 1-51.
- Momo, S.M.C., Chabrerie O., Gallet-Maron E., Nkongmeneck B-A., Leumbe L.O.N., et Decocq G., 2012, Analyse de la dynamique de déforestation par télédétection couplée aux modèles d’équations structurales : exemple de la forêt néphéliphile du mont Oku (Cameroun), Acta Botanica Gallica, 159, 4, pp. 451-466.
- Mpanda, M.M., Khoji M.H., N’Tambwe N.D., Sambieni R.K., Malaisse F., Cabala K.S., Bogaert J., et Useni S.Y., 2022, Uncontrolled Exploitation of Pterocarpus tinctorius Welw. and Associated Landscape Dynamics in the Kasenga Territory: Case of the Rural Area of Kasomeno (DR Congo), Land , 11 , pp. 1-14.
- Otukei, J.R., Blaschke T., 2010, Land cover change assessment using decision trees, support vector machines and maximum likelihood classification algorithms, International Journal of Applied Earth Observation and Geoinformation, 12, 1, pp. S27-S31.
- Pélissier, C., de Marcken P., Mapilanga wa Tsaramu J. J., et Wilungula B. C., 2015, République Démocratique du Congo, dans Doumenge C., Palla F., Scholte P., Hiol Hiol F., Larzillière A. (dir.), Aires protégées d’Afrique centrale – État 2015, OFAC, Kinshasa, République démocratique du Congo, Yaoundé, Cameroun, pp. 111-147.
- Phan, T.N., Kush V., et Lehnert L.W., 2020, Land Cover Classification using Google Earth Engine and Random Forest Classifier—The Role of Image Composition, Remote Sensing , 12, 15, [En ligne], URL : https://www.mdpi.com/2072-4292/12/15/2411
- Polepole Buhendwa, P., 2013, Cadre légal et institutionnel de la gouvernance des ressources naturelles en RD Congo, L’Harmattan, 208 p.
- Pontius, R. G., Millones, M., 2011, Death to Kappa: birth of quantity disagreement and allocation disagreement for accuracy assessment, International Journal of Remote Sensing , 32, 15, pp. 4407–4429.
- Potapov, P.V., Turubanova S., Hansen M.C., Adusei B., Broich M., Altstatt A., Mane L., et Justice C.O., 2012, Quantifying Forest cover loss in Democratic Republic of Congo, 2000-2010 with Landsat ETM+ data, Remote Sensing of Environment , 122 , pp. 106-116.
- Reinartz, G.E., Isia I.B., Ngamankosi M., et Wema L.W., 2006, Effects of forest type and human presence on bonobo (Pan paniscus) density in the Salonga National Park, International Journal of Primates , 27 , pp. 603-634.
- Saito, H., Miura Y., 2019, Skew Detection and Ortho-rectification for System Corrected Landsat TM Images , Conference PhotonIcs & Electromagnetics Research Symposium, Spring, IEEE, pp. 2789-2794.
- Schmitt, C.B., Burgess N. D., Lauren C., Belokurov A., Besançon C., Boisrobert L., Campbell A., Fish L., Gliddon D., Humphries K., Kapos V., Loucks C., Lysenko I., Miles L., Mills C., Minnemeyer S., Pistorius T., Ravilious C., et Steininger Winkel G., 2009, Global analysis of the protection status of the world’s forests, Biological Conservation , 142 , 10, pp. 2122-2130.
- Soulama, S., Kadeba A., Nacoulama B.M.I., Traoré S., Bachamann Y., et Thiombiano A., 2015, Impact des activités anthropiques sur la dynamique de la végétation de la réserve partielle de faune de Pama et de ses périphéries (sud-est du Burkina Faso) dans un contexte de variabilité climatique, Journal of Applied Biosciences, 87, pp. 8047-8064.
- Steinskog, D. J., Tjøstheim D. B., Kvamstø N. G., 2007, A cautionary note on the use of the Kolmogorov–Smirnov test for normality, Monthly Weather Review , 135 , 3, pp. 1151-1157.
- Tingu, C., Mathunabo A., Bondjembo T., et Mabhungu P., 2019, Impact des activités anthropiques sur la viabilité des ressources naturelles du Parc National de la Salonga (PNS), en RD Congo : cas des activités des Yaelima dans le bloc sud, Journal d’Économie, de Management, d’Environnement et de Droit , 2, 2, pp. 86-101.
- UICN – PACO , 2010, Parcs et réserves de la République Démocratique du Congo : évaluation de l’efficacité de gestion des aires protégées , UICN/PACO Ouagadougou, Burkina Faso, 149 p.
- Useni, S.Y., Boisson S., Cabala K., Nkuku K.C., Malaisse F., Halleux J-M., Munyemba K.F., et Bogaert J., 2020b, Dynamique de l’occupation du sol autour des sites miniers le long du gradient urbain-rural de la ville de Lubumbashi, RD Congo, Biotechnologie, Agronomie, Société et Environnement, 44, 1, pp. 1-14.
- Useni, S.Y., Cabala K.S., Halleux J-M., Bogaert J., et Munyemba K.F., 2018, Caractérisation de la croissance spatiale urbaine de la ville de Lubumbashi (Haut-Katanga, R.D. Congo) entre 1989 et 2014, Tropicultura, 36, 1, pp. 99-108.
- Useni, S.Y., Khoji M.H., et Bogaert J., 2020, Miombo woodland, an ecosystem at risk of disappearance in the Lufira Biosphere Reserve (Upper Katanga, DR Congo)? A 39-years analysis based on Landsat images, Global Ecology and Conservation , 24 , pp. 1-15.
- Useni, S.Y., Malaisse F., Cabala K.S., Munyemba K.F., et Bogaert J., 2017, Le rayon de déforestation autour de la ville de Lubumbashi (Haut-Katanga, RD Congo) : Synthèse, Tropicultura, 35, 3, pp. 215-221.
- Van Krunkelsven, E., Draulans D., 2000, A survey of bonobos and other large mammals in the Salonga National Park, Democratic Republic of Congo, Oryx , 34 , 3, pp. 180-187.
- Vermeulen, C., Dubiez E., Proces P., Mukumary S.D., Yamba Yamba T., Mutambwe S., Peltier R., Marien J-N., et Doucet J-L., 2011, Enjeux fonciers, exploitation des ressources naturelles et forêts des communautés locales en périphérie de Kinshasa, RDC, Biotechnologie, Agronomie, Société et Environnement, 15, 4, pp. 535-544.
- Wabasa, S.M., 2017, Repenser le rôle des institutions de conservation de la nature à l’ère de la globalisation en Afrique : À propos du parc national de la Salonga (RDC), European Scientific Journal, 13, 11, pp. 92-111.
- WCS (Wildlife Conservation Society), 2014 , Recensement et enquête socioéconomique sur la communauté Yaelima/Parc National de la Salonga/Sud , 109 p.
- Wittemyer, G., Elsen P., Bean W.T., Burton A.C.O., et Brashares, J.S., 2008, Accelerated human population growth at protected area edges, Science , 321 , 5885, pp. 123-126.
List of figures
Figure 1
Localisation du Bloc Sud du PNS
Figure 2
Localisation des classes d'occupation du sol du bloc sud du parc national de la Salonga
Figure 3
Différentes zones tampons selon la distance autour des villages
Figure 4
Cartes de l’occupation du sol dans le bloc sud du PNS en 2002, 2007, 2014 et 2020
Figure 5
Proportion des forêts primaires et secondaires ainsi que le complexe champs-habitats
Figure
Évolution de la proportion de la forêt primaire (en haut) et de la forêt secondaire (au milieu) et du complexe champs-habitats à différentes distances par rapport aux villages villages (Iyamba, Ila, Nsese, Bosanja, Isambo Botongola, Bokumu Beke) entre 2000 et 2020 sur base de la classification des images Landsat de 2002, 2007, 2014 et 2020. La zone tampon comprise entre 0 et 2 km a une surface de 12,5 km2, celle comprise entre 2 et 4 km a une surface de 37,6 km2 et celle comprise entre 4 et 6 km une surface de 62,7 km2.
List of tables
Tableau 1
Caractéristiques des images Landsat TM, ETM + et OLI
λ est la longueur d'onde (Farah et al. , 2021).
Tableau 2
Description des classes d'occupation du sol
Tableau 3
Précision des classifications supervisées des images Landsat de 2002, 2007, 2014 et 2020 appuyée sur l'algorithme du maximum de vraisemblance
Les valeurs des colonnes réfèrent aux données classifiées et celles des lignes aux données de référence (terrain). Les paramètres de précisions sont en pourcentage (%).
Tableau 4
Matrices de transition (en pourcentage) des classes d’occupation du sol issue de la classification des images Landsat de 2002, 2007, 2014 et 2020
Images Landsat de 2002, 2007, 2014 et 2020 observées au sein du Parc National de la Salonga. 1%= 131,4 km2.
Tableau 5
Valeurs du nombre de taches (n) et de l’aire totale (at) en km2 par année et par classe d’occupation du sol dans le bloc Sud du Parc National de la Salonga en 2002, 2007, 2014 et 2020
Les processus de transformation spatiale (PTS) ont été identifiés sur la base de l’arbre de décision (12).