Article body
Les rythmes circadiens du comportement et de la physiologie des mammifères sont synchronisés par des signaux de lumière reçus par la rétine et communiqués aux noyaux supra-chiasmatiques. Cette horloge centrale synchronise à son tour des horloges esclaves dans la plupart des cellules de notre corps. Sans lumière, l’horloge circadienne dirige des processus comme le sommeil selon sa période intrinsèque, qui varie entre des individus [1]. Dans des conditions normales de jour et de nuit, l’horloge est réajustée deux fois chaque jour, matin et soir, afin de rester synchronisée avec la photopériode de l’environnement (qui est exactement de 24 heures). En revanche, si la longueur de la période intrinsèque de l’horloge diffère considérablement de celle de la photopériode, la phase active d’un individu est en avance ou retarde pendant la journée. La période intrinsèque étant déterminée par des facteurs génétiques, certains syndromes de décalage chronique de sommeil peuvent avoir des causes héréditaires liées aux polymorphismes des gènes de l’horloge [2]. Plus généralement, on estime que des individus ayant une période circadienne plus longue sont ceux dont l’activité se prolonge plus tardivement, et que les individus matinaux ont une période circadienne plus courte [3].
Malgré la diversité des phases du comportement chez l’homme, la caractérisation des gènes de l’horloge et leur influence sur le comportement restent difficiles à établir en raison de la quasi-impossibilité d’analyser de nombreux sujets pendant suffisamment longtemps (10-30 jours) dans des conditions de laboratoire constantes. Mais, par chance, il existe aussi une horloge autonome cellulaire dans les fibroblastes de la peau (et dans la plupart des autres cellules du corps). Ces horloges fonctionnent selon un mécanisme similaire ou identique à celle du SNC. Par conséquent, chez la souris, les différences génétiques ne s’expriment pas seulement dans le SNC, mais aussi dans les horloges périphériques [4].
Dans une étude récente [5], à partir de biopsies de peau prélevées chez 20 sujets, nous avons mis au point une méthode d’analyse des rythmes circadiens à partir des fibroblastes issus de ces biopsies. Nous avons utilisé un lentivirus contenant un promoteur circadien - celui du gène Bmal1 de l’horloge - qui dirige l’expression de la protéine luciférase, responsable de l’émission de lumière chez la luciole. Quand les fibroblastes provenant de ces biopsies sont infectés par ce lentivirus, ils émettent alors des photons selon un rythme circadien dépendant de la période intrinsèque de leurs horloges (Figure 1). Le nombre de photons, qui peut être mesuré par des tubes photomultiplicateurs à haute sensibilité, suit l’expression circadienne du gène Bmal1.
Figure 1

A. 50 000 fibroblastes sont cultivés à partir d’une seule biopsie de peau de 2 mm et sont infectés avec une sonde lentivirale. Les rythmes circadiens de ces cellules ont été synchronisées par la dexaméthasone [10], et le rythme de bioluminescence a ensuite été mesuré dans un luminomètre pendant plusieurs jours. B. Protocole identique appliqué à 50 000 kératinocytes cultivés à partir d’un poil.
Il a d’abord été vérifié que la période des fibroblastes est propre à l’individu et ne dépend, ni du nombre de particules virales utilisées dans l’infection, ni de l’endroit anatomique de la biopsie cutanée, ni du moment où la biopsie a été prélevée. La précision de notre méthode a été estimée à moins d’une demi-heure près pour les cellules d’un même individu. Ensuite, les rythmes circadiens des fibroblastes dans deux à quatre biopsies de chacun des 20 individus ont été mesurés (Figure 2A). La période moyenne que nous avons trouvée, 24,5 heures, est très similaire à celle constatée par d’autres groupes à partir d’observations du comportement de sujets, soit 24,2-24,5 heures. Néanmoins, la variabilité entre individus – avec une déviation standard de 1,5 heures et une variance maximale de 4 heures – est bien plus importante que celle enregistrée à partir du comportement, qui montrait une déviation standard de seulement 20 minutes [6, 7].
Figure 2
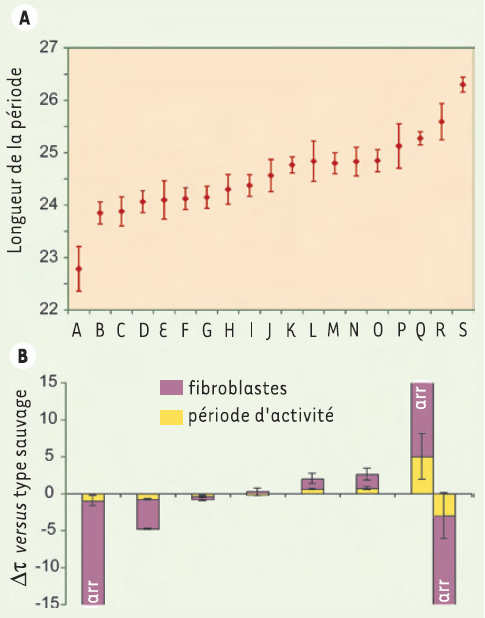
A. Biopsies prélevées à partir de peau provenant de l’abdomen, des fesses, ou du prépuce de 19 individus. Les fibroblastes ont ensuite été cultivés et les rythmes circadiens mesurés comme sur la Figure 1. Les individus sont désignés par les lettres A-S. B. Périodes des rythmes circadiens d’activité et de bioluminescence de fibroblastes chez diverses souches de souris, exprimées en différence par rapport au jour solaire de 24 heures. La période d’activité est représentée en jaune et, au-dessus, la période de bioluminescence des fibroblastes en violet. Les génotypes des souches, de gauche à droite, sont Per2brdm/brdm, Per1brdm/brdm, type sauvage, Cry2+/-, Cry2-/- ; Per1brdm/brdm,Cry2-/-, Per2brdm/brdm ; Cry2-/-.Per2brdm/brdm ; Cry2-/- ; les souris possédaient des périodes instables, de plus ou moins 24 heures. Leurs périodes sont ainsi montrées deux fois à droite. Les fibroblastes arythmiques sont désignés par les lettres « arr » (les références se rapportant aux modifications génétiques de ces souris se trouvent dans notre publication originale [5]).
Pour tenter de comprendre l’origine de cette variabilité, nous avons examiné notre système chez des souris génétiquement modifiées dont les périodes diffèrent considérablement de celles des souris de type sauvage. Nous avons pu mesurer la période circadienne intrinsèque de l’activité locomotrice de ces souris de façon extrêmement précise à l’aide d’une cage équipée d’une roue tournante, en déterminant le nombre de révolutions de la roue par unité de temps pendant plusieurs jours en obscurité totale. Parallèlement, nous avons déterminé la période des fibroblastes primaires de ces mêmes souris en utilisant notre système de lentivirus. De cette façon, nous avons pu corréler la longueur de la période circadienne du comportement avec celle des fibroblastes des mêmes souris. Nos résultats ont démontré que si une souris avait une période de d’activité comportementale de moins de 24 heures, ses fibroblastes avaient également une période de moins de 24 heures. De même, si une souris a une période plus longue que 24 heures, ses fibroblastes ont une période supérieure à 24 heures. Curieusement, la différence de période mesurée pour les fibroblastes de souris mutantes et de souris de type sauvage est toujours plus grande que la différence manifestée dans le comportement. Par exemple, les souris Per1-/- avec une période d’activité de 23 heures ont des fibroblastes produisant une période d’expression de seulement 20 heures en culture (Figure 2B) [8]. Si cette conclusion est applicable à l’homme, elle peut expliquer que la variabilité des périodes mesurées dans les fibroblastes est plus grande que celle des périodes déterminées pour le comportement également dans l’espèce humaine.
Bien que la période des horloges du SNC (qui détermine la période du comportement) n’est pas identique à celle des horloges des fibroblastes chez la souris, il existe une corrélation qualitative nette entre les deux. Tout porte à croire qu’une telle corrélation existe aussi chez l’homme. En conséquence, notre méthode pour déterminer la longueur de période circadienne chez l’homme pourrait fournir un protocole expérimental pour identifier les gènes liés aux différences circadiennes du comportement. D’autres chercheurs ont déjà utilisé la période du comportement (mesurée par roue tournante) avec grand succès pour l’identification des gènes qui influencent l’horloge par association génétique chez la souris [9]. La simplicité de notre méthode devrait maintenant rendre possible des études similaires chez l’homme. Jusqu’à ce jour, nous avons utilisé surtout des biopsies de peau pour nos études, mais dans des cas isolés, il a été possible de cultiver des cellules provenant d’un seul bulbe pileux à partir d’un poil arraché, et de déterminer le cycle circadien du sujet par la même technologie lentivirale. Une fois perfectionnée, une telle méthode, peu invasive, devrait permettre la mesure des rythmes circadiens chez de nombreux sujets et ainsi d’être en mesure d’effectuer une étude génétique exhaustive des différents « chronotypes » humains.
Appendices
Références
- 1. Schibler U, Sassone-Corsi P. A web of circadian pacemakers. Cell 2002 ; 111 : 919-22.
- 2. Toh KL, Jones CR, He Y, et al. An hPer2 phosphorylation site mutation in familial advanced sleep-phase syndrome. Science 2001 ; 291 : 1040-3.
- 3. Duffy JF, Rimmer DW, Czeisler CA. Association of intrinsic circadian period with morningness-eveningness, usual wake time, and circadian phase. Behav Neurosci 2001 ; 115 : 895-9.
- 4. Yagita K, Tamanini F, van der Horst GT, et al. Molecular mechanisms of the biological clock in cultured fibroblasts. Science 2001 ; 292 : 278-81.
- 5. Brown SA, Fleury-Olela F, Nagoshi E, et al. The period length of fibroblast circadian gene expression varies widely among human individuals. PloS Biol2005 ; 3 : e338.
- 6. Czeisler CA, Duffy JF, Shanahan TL, et al. Stability, precision, and near-24-hour period of the human circadian pacemaker. Science 1999 ; 284 : 2177-81.
- 7. Carskadon MA, Labyak SE, Acebo C, et al. Intrinsic circadian period of adolescent humans measured in conditions of forced desynchrony. Neurosci Lett 1999 ; 260 : 129-32.
- 8. Pando MP, Morse D, Cermakian N, et al. Phenotypic rescue of a peripheral clock genetic defect via SCN hierarchical dominance. Cell 2002 ; 110 : 107-17.
- 9. Shimomura K, Low-Zeddies SS, King DP. Genome-wide epistatic interaction analysis reveals complex genetic determinants of circadian behavior in mice. Genome Res 2001 ; 11 : 959-80.
- 10. Balsalobre A, Brown SA, Marcacci L, et al. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science 2000 ; 289 : 2344-7.
List of figures
Figure 1
A. 50 000 fibroblastes sont cultivés à partir d’une seule biopsie de peau de 2 mm et sont infectés avec une sonde lentivirale. Les rythmes circadiens de ces cellules ont été synchronisées par la dexaméthasone [10], et le rythme de bioluminescence a ensuite été mesuré dans un luminomètre pendant plusieurs jours. B. Protocole identique appliqué à 50 000 kératinocytes cultivés à partir d’un poil.
Figure 2
A. Biopsies prélevées à partir de peau provenant de l’abdomen, des fesses, ou du prépuce de 19 individus. Les fibroblastes ont ensuite été cultivés et les rythmes circadiens mesurés comme sur la Figure 1. Les individus sont désignés par les lettres A-S. B. Périodes des rythmes circadiens d’activité et de bioluminescence de fibroblastes chez diverses souches de souris, exprimées en différence par rapport au jour solaire de 24 heures. La période d’activité est représentée en jaune et, au-dessus, la période de bioluminescence des fibroblastes en violet. Les génotypes des souches, de gauche à droite, sont Per2brdm/brdm, Per1brdm/brdm, type sauvage, Cry2+/-, Cry2-/- ; Per1brdm/brdm,Cry2-/-, Per2brdm/brdm ; Cry2-/-.Per2brdm/brdm ; Cry2-/- ; les souris possédaient des périodes instables, de plus ou moins 24 heures. Leurs périodes sont ainsi montrées deux fois à droite. Les fibroblastes arythmiques sont désignés par les lettres « arr » (les références se rapportant aux modifications génétiques de ces souris se trouvent dans notre publication originale [5]).