Abstracts
Résumé
La ghréline, peptide essentiellement produit par l’estomac endocrine, a été découverte en tant que ligand naturel du récepteur des sécrétagogues de l’hormone de croissance (GH). Très rapidement, elle s’est révélée être le premier facteur orexigène périphérique. À côté de cette fonction, la ghréline influence de nombreuses voies neuroendocriniennes, métaboliques et extraendocriniennes, notamment au sein du système cardiovasculaire. La liaison de la ghréline au récepteur des sécrétagogues de GH de type 1a (GHS-R 1a) nécessite la présence, sur le peptide, d’une acylation de la sérine située en position 3. Si cette modification est indispensable pour la stimulation de la GH et pour certains effets métaboliques, elle n’est pas requise pour les autres fonctions du peptide, dans lesquelles d’autres récepteurs sont probablement impliqués. Les souris invalidées pour le gène de la préproghréline ne sont ni anorexiques, ni naines. En revanche, les souris GHS-R–/– sont légèrement moins lourdes, et les actions GH-sécrétagogue et orexigène de la ghréline sont neutralisées chez ces animaux. Ainsi, l’histoire fascinante de la ghréline et ses implications physiopathologiques potentielles en endocrinologie et en médecine interne sont encore en devenir.
Summary
Ghrelin, a peptide predominantly produced by the stomach, has been discovered as a natural ligand of the growth hormone secretagogue receptor (GHS-R) type 1a. Shortly there after, it attracted enormous interest since it appeared as the first peripheral orexigenic factor. Besides, ghrelin exerts other neuroendocrine metabolic and non-endocrine actions (e.g. cardiovascular activities) that may rely on the widespread distribution of ghrelin and its receptor (GHS-R). The existence of several GHS-R subtypes and evidences that neuroendocrine and metabolic but not all other ghrelin actions are dependent on acylation on serine 3 add further complexity to the system whose major physiological role remains to be definitely elucidated. Ghrelin knockout(-/-) mice are neither anorectic nor dwarf though GHS-R-/- are slightly underweight and do not respond to ghrelin with increased GH secretion or appetite. Thus, the continuation of the fascinating ghrelin story as well as its potential pathophysiological implications in endocrinology and internal medicine remain open avenues for future investigations.
Article body
L’histoire de la ghréline (de la racine sanscrite ghr : croissance) commence vingt-trois ans avant sa découverte en tant que ligand naturel du récepteur des sécrétagogues de l’hormone de croissance (GH) à partir d’extraits d’estomac de rat, fin 1999 [1]. En effet, c’est en 1976 que Cyril Bowers, en recherchant des agonistes opiacés, observe qu’un dérivé de la met-enképhaline, le Tyr-DTrp-Gly-Gly-Phe-MetNH2, stimule la sécrétion de GH in vitro, alors qu’il ne présente plus d’activité opiacée [2] (Figure 1). La découverte de ce premier sécrétagogue de la GH (GHS) est suivie, en 1984, par celle du GH-releasing peptide-6 (GHRP-6, His-DTrp-Ala-Trp-Dphe-LysNH2), première molécule active in vitro et in vivo. Après la mise en évidence de la GHRH (growth hormone releasing hormone), neurohormone hypothalamique directement responsable de la sécrétion de GH hypophysaire, les GHS sont cependant considérés comme une curiosité pharmacologique sans pertinence physiologique.
Figure 1
Ghréline et autres sécrétagogues de la GH (GHS), et leur récepteur GHS-R 1a.
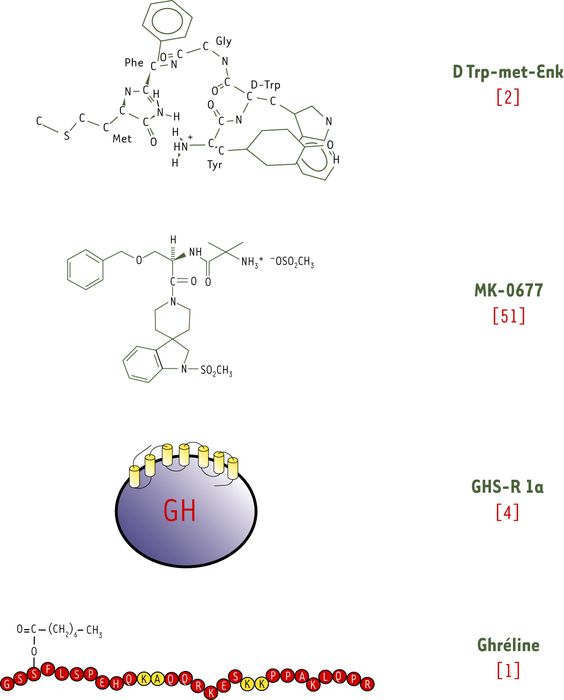
Ce n’est qu’au début des années 90, lorsque des effets bénéfiques d’un traitement par la GH recombinante sont observés chez les personnes âgées (diminution de la masse grasse, augmentation de la masse maigre, amélioration du bien-être…), que l’industrie pharmaceutique s’intéresse de plus près aux GHS. Merck emploie une stratégie de pharmacologie inverse en déterminant le système de transduction utilisé par le GHRP-6, qui se révèle rapidement être la mobilisation du calcium intracellulaire ([Ca2+]i) par l’intermédiaire d’une protéine Gq. En trois ans, des peptidomimétiques sont développés, faisant passer l’affinité des composés du micromolaire au nanomolaire, avec une biodisponibilité et une demi-vie permettant les études chez l’homme. Une spiropéridine, le MK 0677 (Figure 1), entraîne en administration orale chronique une « jouvence » de l’axe somatotrope chez la personne âgée [3]. Après radiomarquage, cette même molécule permet la mise en évidence de récepteurs au niveau de l’antéhypophyse et de l’hypothalamus. L’expression infime du récepteur nécessite alors un clonage par expression, qui aboutit en 1996 à la découverte d’un récepteur orphelin, couplé à une protéine Gq, baptisé GHS-R [4].
Ayant cloné le récepteur, la découverte du ligand endogène paraît à portée de main. Effectivement, on découvre que l’adénosine hypothalamique est un ligand endogène du récepteur [5] ; toutefois, elle ne stimule pas la sécrétion de GH. C’est un groupe japonais qui, à partir des homologies de séquence entre le GHS-R et le récepteur de la motiline, un peptide gastro-intestinal, entreprend une approche systématique d’extraction et de purification de peptides, de l’oesophage jusqu’au colon, afin d’évaluer la capacité des extraits acides bouillis à stimuler le [Ca2+]i de cellules surexprimant le GHS-R. Un extrait d’estomac correspondant à un peptide de 28 acides aminés est ainsi sélectionné, mais le peptide synthétique se révèle inactif. Cependant, le temps de rétention de l’extrait naturel en RP-HPLC est légèrement plus long que celui du peptide synthétique, et la spectrométrie de masse de la ghréline purifiée donne une masse moléculaire de 3 315, contre 3 189 au peptide synthétique de 28 acides aminés. Ce peptide montre d’ailleurs des homologies de séquences si fortes avec la motiline qu’il est indépendamment dénommé motilin-like related-peptide [6]. Les chimistes japonais pensent alors que la modification la plus probable pour expliquer une différence de masse de 144 est l’addition d’un acide gras, l’acide n-octanoïque, sur la sérine en position 3. Le peptide de 28 acides aminés, octanoylé sur la sérine en position 3, est synthétisé et se révèle parfaitement bioactif : la quête pour la ghréline est terminée, mais non les surprises…
Combien de ligands et de récepteurs ?
Chez l’homme, le gène de la préproghréline est localisé sur le chromosome 3, en 3p25-26. Quatre exons et trois introns ont été décrits en position 141 (2 000 bp intron), 258 (3 000 bp intron) et 367 (800 bp intron) d’un ADNc de 511 bp. Cet ADNc code pour une protéine de 117 acides aminés, ayant un peptide signal de 23 acides aminés et une forme proghréline de 94 acides aminés, dont les 28 premiers constituent, après un processus d’acylation sur la sérine en position 3, la ghréline (Figure 1). Les quatre ou cinq premiers acides aminés, essentiels pour activer la mobilisation du [Ca2+]i par le GHS-R 1a, sont conservés au cours de l’évolution [7]. Un variant d’épissage, aboutissant à la synthèse d’un peptide mature ghréline de 27 acides aminés dépourvu d’un résidu glutamine en position 14, possède les mêmes propriétés que le peptide de 28 acides aminés.
Le gène codant pour le récepteur GHS-R est également situé sur le chromosome 3, en 3q26.2. Il code pour un récepteur à sept domaines transmembranaires de 366 acides aminés, le GHS-R 1a (Figure 1). Un variant d’épissage a été décrit, codant pour une protéine de 289 acides aminés à cinq domaines transmembranaires, mais l’existence de cette protéine, nommée GHS-R 1b, n’a jamais été démontrée in vivo. Dans des lignées transfectées, le type 1b ne lie pas le MK 0677 et ne présente pas d’activité biologique de libération de GH ou de mobilisation du [Ca2+]i.
La ghréline et les GHS augmentent le [Ca2+]i et le métabolisme des inositols phosphates. Lorsqu’il est transfecté, le GHS-R 1a est constitutivement actif [8] : la mesure du métabolisme des IP3 montre en effet que, en l’absence d’agoniste, le récepteur présente déjà 50 % de son activité. Deux ligands endogènes putatifs ont été décrits (Tableau I) : l’adénosine est un agoniste partiel, moins puissant que les GHS sur la mobilisation du [Ca2+]i et inactif sur la libération de GH [5] ; la cortistatine, un peptide cérébral apparenté à la somatostatine, se lie quant à elle aux sites de liaison hypophysaires de la ghréline et atténue ses effets sur la GH. Cependant, il est probable que cet effet passe également par les récepteurs de la somatostatine [9].
Tableau I
Récepteurs et ligands.

L’adénosine ne stimule pas la sécrétion de GH, la cortistatine atténue les effets de la ghréline. EP 51389 et TH 2306.34 stimulent la sécrétion de GH, BIM 28131 augmente la prise de poids, TZP 101 stimule la vidange gastrique. BIM 28163 inhibe l’effet de la ghréline sur la GH, mais augmente la prise de poids (il agit donc également par l’intermédiaire d’un autre récepteur). L’hexaréline, via CD36, prévient la formation d’athérosclérose.
Le récepteur GHS-R 1a ne rend pas compte de tous les sites de liaison de la ghréline ou des GHS. Dans le plasma, la ghréline se lie à un complexe lipoprotéique comprenant la clusterine (apolipoprotéine J) et la paraoxonase [10]. Cette estérase clive les lipides oxydés en LDL, et une faible activité paraoxonase est liée à la maladie coronarienne. Cette enzyme aurait donc pu être impliquée dans les mécanismes de désacylation de la ghréline : il ne semble pas que ce soit le cas [11]. L’hexaréline, mais pas le MK 0677 ni la ghréline, se fixe avec une affinité raisonnable au CD36, un récepteur d’épuration multifonctionnel [12]. Enfin, le (ou les) récepteur(s) relayant les effets de la ghréline non acylée reste(ent) totalement inconnus…
Effets neuroendocriniens et métaboliques
La ghréline augmente non seulement la sécrétion de GH, mais également, et rapidement, celle de l’ACTH (hormone adrénocorticotrope), du cortisol et de la prolactine [13]. Un traitement aigu par la ghréline ou par des GHS synthétiques stimule également la prise alimentaire [14]. Si la ghréline est essentiellement synthétisée par les cellules endocrines de l’estomac, l’existence d’un système neuronal intrahypothalamique [15] reste encore débattue [16]. Il est admis que la ghréline agit majoritairement par l’intermédiaire de récepteurs intrahypothalamiques exprimés par les neurones à GHRH [17], pour stimuler la libération de GH et, de manière indépendante, sur les neurones à neuropeptide Y/AgRP (Agouti-related protein), pour augmenter la prise alimentaire [18]. Cependant, elle agit aussi directement au niveau hypophysaire sur la sécrétion de GH [19]. La sécrétion gastrique et le complexe dorsovagal sont également impliqués [20]. Le récepteur de la ghréline est synthétisé dans les neurones vagaux et transporté aux terminaisons afférentes ; si la ghréline est administrée à des rats vagotomisés, elle ne stimule plus l’appétit et son effet sur la GH est considérablement diminué.
La régulation de l’appétit et de la prise de poids est un phénomène très complexe. Sur la base des effets du peptide administré de façon exogène, la ghréline apparaissait comme un régulateur important de la prise alimentaire et de la sécrétion de GH à court terme et, par conséquent, du poids corporel. Étant donné ses effets sur la GH, hormone lipolytique, on aurait pu s’attendre à une diminution de la masse grasse ; or la ghréline, en traitement chronique, augmente l’adiposité [21], et ce indépendamment d’une augmentation de la prise alimentaire, cet effet étant observé chez des animaux disposant d’une quantité limitée de nourriture. Par ailleurs, un traitement chronique par la ghréline entraîne un accroissement du quotient respiratoire, ce qui suggère une augmentation de l’ingestion des carbohydrates ou une diminution de celle des lipides, avec augmentation subséquente de leur stockage. Ce paradoxe semble en partie résolu : l’effet adipogène de la ghréline pourrait impliquer les formes non octanoylées du peptide, avec action directe sur les adipocytes par l’intermédiaire d’un récepteur distinct du GHS-R 1a [22]. L’importance du rôle orexigène et adipogène de la ghréline a cependant été remise en cause par les résultats obtenus sur des souris invalidées pour le gène de la préproghréline qui, en première analyse, ne présentent aucun phénotype particulier en termes de croissance ou d’appétit [23] : les souris ghréline–/– s’ajoutent ainsi à la liste des animaux qui, invalidés pour différents gènes de facteurs orexigènes (NPY, AgRP), ne montrent aucun phénotype majeur, notamment en comparaison des souris déficientes pour les gènes de facteurs anorexigènes (MCH-melanin concentrating hormone, leptine, mélanocortine) et de leurs récepteurs. En revanche, les souris invalidées pour le gène du récepteur GHS-R ne répondent plus ni à la ghréline, ni au MK 0677 en termes de sécrétion de GH ou de prise alimentaire [24] ; de plus, leur poids corporel est légèrement inférieur dès 16 semaines, et la concentration en IGF-1 (insulin-like growth factor) plus basse (– 21 %) à 24 semaines [16]. Pris dans leur ensemble, ces résultats suggèrent que l’absence de ghréline est compensée durant l’embryogenèse. Aucune donnée concernant les autres rôles de la ghréline, sur la motricité gastro-intestinale ou la sphère cardiovasculaire, n’a été rapportée dans ces modèles (Tableau II).
Tableau II
Modèles animaux d’invalidation ou de surexpression de la ghréline et du récepteur GHS-R.

Effets gastro-intestinaux
Étant donné sa localisation dans les cellules A de l’estomac, il n’est pas surprenant que la ghréline agisse sur la sécrétion acide gastrique et la motricité. Ces effets sont abolis lors d’un traitement par l’atropine et par une vagotomie bilatérale, mais pas par des antagonistes H2 [25]. L’activité motrice sur le tractus gastrointestinal passe en partie par les neurones à NPY et le nerf vague, mais il existe un effet direct analogue à celui de la motiline [26]. En revanche, la ghréline, mais pas la motiline, est capable de contrecarrer l’ileus gastrique postopératoire [27]. Elle protège également de l’ulcère gastrique induit par l’éthanol [28]. L’infection par Helicobacter pylori n’affecte pas les concentrations circulantes de ghréline, mais son éradication s’accompagne d’une augmentation de l’expression du peptide, ce qui pourrait expliquer l’augmentation d’appétit et la reprise de poids observées dans ces conditions [29]. Au niveau du pancréas exocrine, la ghréline est un puissant inhibiteur de la sécrétion d’amylase, par un mécanisme indirect impliquant probablement des neurones intrapancréatiques [30]. Elle augmente les concentrations sanguines de peptides gastrointestinaux, comme la somatostatine ou le polypeptide pancréatique, sans modifier les niveaux de gastrine [31]. Enfin, elle inhibe la sécrétion d’insuline, par un mécanisme qui pourrait faire intervenir sa forme non acylée [32].
Effets cardiovasculaires
Le rôle cardioprotecteur de la GH et de l’IGF-1 étant bien établi, il paraissait logique d’évaluer les effets des sécrétagogues de la GH (GHS) sur la fonction cardiaque. Des effets bénéfiques des GHS sur la fonction cardiaque ont été observés chez des rats pourtant hypophysectomisés ou chez des patients déficients en GH [33, 34]. De plus, la ghréline et les GHS augmentent les performances cardiaques et exercent des effets protecteurs sur les atteintes ischémiques/reperfusion sur le coeur isolé [35]. Enfin, la ghréline et les GHS synthétiques agissent comme des facteurs de survie, en protégeant les cardiomyocytes et les cellules endothéliales contre l’apoptose induite par la doxorubicine. La ghréline est elle-même synthétisée et sécrétée par les cardiomyocytes [36].
Bien que les actions neuro-endocriniennes de la ghréline dépendent de son acylation sur la sérine 3, ses effets cardiovasculaires sont également observés avec la forme non acylée, ce qui suggère que ces actions ne dépendent pas du GHS-R 1a. Le CD36 est un récepteur d’épuration multifonctionnel de type B, impliqué dans l’athérogenèse et le métabolisme des lipoprotéines. Exprimé principalement par les cardiomyocytes et les cellules endothéliales de la microvasculature, le CD36 semble intervenir dans les effets cardiovasculaires sélectifs des GHS de nature peptidique [12].
Des effets de la ghréline ont également été décrits sur l’axe reproducteur (la ghréline inhibe l’activité proliférative des cellules de Leydig immatures), sur les systèmes immunitaires et ostéo-articulaires, ou encore sur le sommeil et l’anxiété [35]. Mais il est souvent difficile de déterminer s’il s’agit d’une action directe ou secondaire aux effets décrits plus haut (Figure 2).
Figure 2
Effets pléïotropes de la ghréline et des sécrétagogues de la GH.
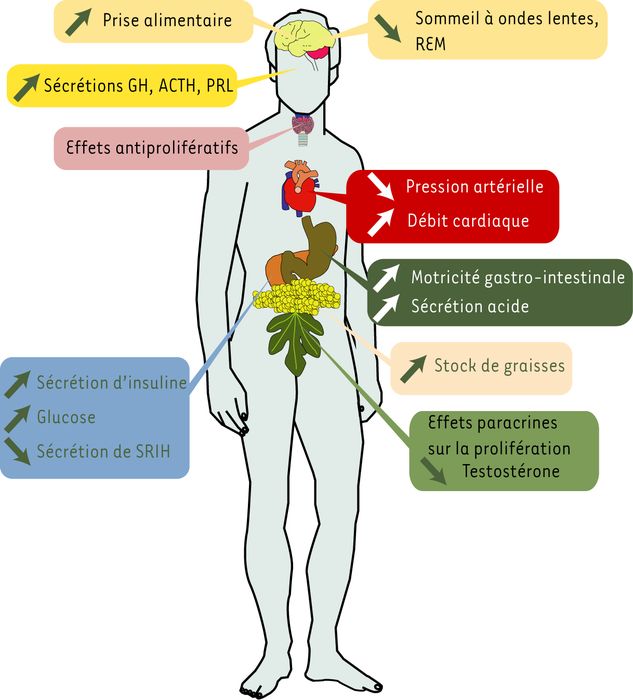
L’acylation de la ghréline est nécessaire aux effets centraux, aux sécrétions hypophysaires, à la prise alimentaire et à certains effets métaboliques et gastro-intestinaux. La forme non acylée est impliquée dans l’adipogenèse, les effets cardiovasculaires et les phénomènes de prolifération/différenciation.
Régulation physiologique de la ghréline et du GHS-R 1a
Un frein majeur pour évaluer précisément le rôle de la ghréline endogène est l’absence de méthode de dosage suffisamment précise. En effet, la plupart des dosages commerciaux reconnaissent indifféremment les formes acylée et non acylée de la ghréline circulante. De plus, les demi-vies de ces formes diffèrent, la forme acylée disparaissant beaucoup plus rapidement, ce qui pourrait expliquer qu’elle ne représente que 10 % à 20 % de la ghréline plasmatique totale.
De nombreux expoirs ont été fondés sur la capacité de la ghréline à mimer ou amplifier l’oscillateur biologique régulant la sécrétion de GH, qui s’effectue selon un rythme ultradien. Néanmoins, s’il existe bien une rythmicité de la sécrétion de ghréline « totale », elle apparaît plus en phase avec la prise de nourriture qu’avec la sécrétion de GH [37]. Une telle absence de relation entre les concentrations de ghréline et de GH circulantes a ainsi été observée chez un patient souffrant d’un ghrélinome pancréatique, ne présentant aucun signe d’acromégalie, et qui avait des concentrations de ghréline plasmatique 50 fois supérieures aux valeurs témoins, mais des concentrations normales de GH et d’IGF-1 [38] (Tableau III). Il est vrai que les récepteurs GHS-R 1a hypophysaires se désensibilisent rapidement [39]. De manière intéressante, l’effet adipogène de la ghréline en infusion continue semble moins sensible à la désensibilisation [21].
Tableau III
Non-corrélation entre valeurs circulantes de GH et de ghréline, en relation avec la prise alimentaire, dans différentes conditions physiopathologiques.
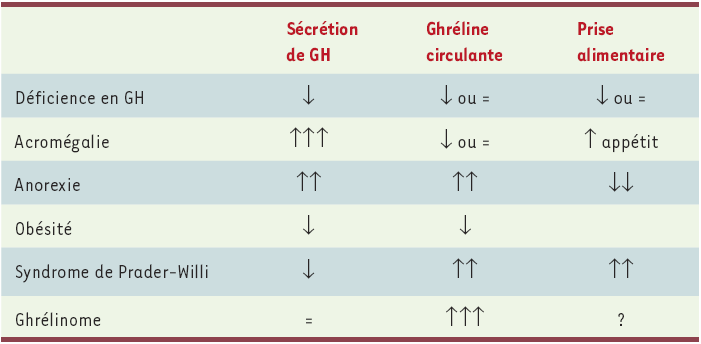
Dans le cas de l’obésité, des modifications non univoques de l’appétit et des préférences alimentaires sont observées. Un seul cas de ghrélinome a été décrit dans la littérature, et le comportement alimentaire de ce patient n’a pas été étudié.
Les pics de ghréline sont observés immédiatement avant les repas, et les nadirs juste après [40]. Une variation circadienne a également été décrite, avec des concentrations plus élevées en début de nuit, suivies d’une chute juste avant le réveil [40]. Le jeûne est un régulateur majeur de la concentration de ghréline circulante, mais l’effet s’atténue avec le temps et les concentrations reviennent aux valeurs de base au bout de quelques jours. Chez l’homme, en l’absence d’indice temporel ou de nourriture, l’augmentation de ghréline est bien corrélée à la sensation de faim, ce qui renforce l’hypothèse du rôle orexigène du peptide [40].
Il existe une corrélation négative entre l’indice de masse corporelle et les concentrations plasmatiques de ghréline à jeun : l’amaigrissement chez le sujet obèse s’accompagne d’une augmentation des niveaux de ghréline [41], tandis que l’inverse est observé avec la prise de poids en cas d’anorexie nerveuse [42]. Les patients insulinorésistants ou souffrant d’un diabète de type 2 présentent également de faibles concentrations de ghréline circulante [43], toujours en lien avec leur indice de masse corporelle [44].
Bien qu’un grand nombre de données sur la régulation de la ghréline et de l’expression du GHS-R par les hormones [35] aient été obtenues chez le rongeur, les différences interespèces compliquent leur interprétation.
Et la clinique dans tout çà ?
La première application des sécrétagogues de la GH (GHS) a été le diagnostic des déficiences en GH, en combinant GHRH et GHRP-6 [45]. Dans ce type d’affections, les GHS ne peuvent être utiles, d’un point de vue thérapeutique, que chez des patients ayant un axe hypothalamo-hypophysaire intact (brûlures, blessures, fractures, retard de croissance intra-utérin, personne âgée fragile…) (pour revue, voir [3]).
Un autre paradoxe de la ghréline a surgi de l’étude du syndrome de Prader-Willi, une des causes les plus fréquentes d’obésité d’origine génétique liée à une délétion en 15q11-15q13 et caractérisée par une hyperphagie massive, une déficience en GH et un hypogonadisme. Alors que les concentrations de ghréline sont abaissées dans tous les cas d’obésité humaine d’origine génétique (déficience en leptine, mutations du récepteur de la leptine ou du récepteur 4 de la mélanocortine, MC4), ou hypothalamique (craniopharyngiomes), elles sont particulièrement élevées dans le syndrome de Prader-Willi [46], du même ordre de grandeur qu’au cours de l’anorexie nerveuse : il a été proposé que ces concentrations très élevées de ghréline pouvaient contribuer à l’hyperphagie sévère des enfants atteints. Cependant, une perfusion de somatostatine chez quatre sujets adultes amènent les niveaux de ghréline aux valeurs normales sans diminuer l’appétit des patients [46]. Il est vrai qu’en même temps que la concentration de ghréline diminue, les concentrations du peptide YY3-36 (PYY3-36), une hormone anorexigène d’origine intestinale, augmentent… Il convient d’attendre la réalisation d’études utilisant un antagoniste du GHS-R 1a pour conclure quant au rôle de la ghréline dans l’hyperphagie liée à ce syndrome.
Dix-sept polymorphismes (SNP) ont été décrits pour le gène de la ghréline (pour revue, voir [35]). Le plus commun, Leu72Met, situé dans la séquence de la proghréline, est observé avec une fréquence de l’ordre de 10 % dans les populations caucasiennes. Il a été associé à un développement plus précoce de l’obésité dans certaines études, mais pas dans toutes, ainsi qu’à un indice de masse corporelle plus élevé et des concentrations d’insuline inférieures lors d’un test d’intolérance au glucose réalisé chez des enfants obèses de grande taille. Le polymorphisme Leu72Met n’intervient pas sur les concentrations circulantes du peptide mature dans sa forme bioactive ou non acylée. Le deuxième polymorphisme le plus fréquent (0,5 % à 2 % de la population selon les études), est Arg51Gln, situé au niveau du dernier acide aminé de la ghréline. Ce polymorphisme est susceptible de modifier la maturation de la proghréline et il a été associé (ou non) à des concentrations plus basses de ghréline circulante et à un risque accru de diabète de type 2.
En ce qui concerne le gène du GHS-R, 12 SNP ont été identifiés [35]. Deux d’entre eux ont été associés à des comportements boulimiques chez des enfants obèses. La mutation Ala204Glu, qui diminue considérablement la liaison de la ghréline au récepteur GHS-R, a été retrouvée chez des enfants présentant un retard de croissance idiopathique d’origine familiale, avec tendance à l’obésité [47].
Conclusions
En résumé, découverte comme sécrétagogue de la GH, la ghréline se révèle être une hormone orexigène et adipogène circulante, d’origine gastrique, principalement : elle est en effet synthétisée par de nombreux tissus, ce qui suggère un rôle endocrine/paracrine plus diversifié. La ghréline semble idéalement placée, avec ses effets centraux et périphériques convergeant vers une utilisation ou un stockage de l’énergie.
Malgré la réelle potentialité clinique de la ghréline ou de ses analogues synthétiques, agonistes ou antagonistes, un certain nombre de questions restent posées : combien existe-t-il de récepteurs pour la ghréline ? La ghréline est-elle « le » ou seulement un des ligands endogènes des GHS-R ? Existe-t-il un équilibre physiologique entre la forme acylée et la forme non acylée et, si oui, quel est son rôle dans les multiples actions de la ghréline ? Toutes ces questions nécessitent une réponse afin de déterminer plus précisément le potentiel clinique et thérapeutique du système ghréline.
Appendices
Références
- 1. Kojima M, Hosoda H, Date Y, et al. Ghrelin is a growth hormone-releasing peptide from stomach. Nature 1999, 402 : 656-60.
- 2. Bowers CY, Chang J, Momany FA, et al. Effects of enkephalins and enkephalins analogs on release of pituitary hormones in vitro. In : Mc Intyre I, Szelke M, eds. Molecular endocrinology. Amsterdam : Elsevier, 1977 : 287-92.
- 3. Thorner MO, Chapman IM, Gaylinn BD, et al. Growth hormone-releasing hormone and growth hormone-releasing peptide as therapeutic agents to enhance growth hormone secretion in disease and aging. Recent Prog Horm Res 1997 ; 52 : 215-44.
- 4. Howard AD, Feighner SD, Cully DF, et al. A receptor in pituitary and hypothalamus that functions in growth hormone release. Science 1996 ; 273 : 974-7.
- 5. Tullin S, Hansen BS, Ankersen M, et al. Adenosine is an agonist of the growth hormone secretagogue receptor. Endocrinology 2000 ; 141 : 3397-402
- 6. Tomasetto C, Karam SM, Ribieras S, et al. Identification and characterization of a novel gastric peptide hormone : the motilin-related peptide. Gastroenterology 2000 ; 119 : 395-405.
- 7. Kojima M, Hosoda H, Kaiya H, et al. Discovery of ghrelin, an endogenous ligand for the growth hormone secretagogue receptor. In : Ghigo E, ed. Ghrelin. Boston : Kluwers, 2004 : 15-25.
- 8. Holst B, Cygankiewicz A, Jensen TH, et al. High constitutive signaling of the ghrelin receptor : identification of a potent inverse agonist. Mol Endocrinol 2003 ; 17 : 2201-10.
- 9. Broglio F, Arvat E, Benso A, et al. Endocrine activities of cortistatin-14 and its interaction with GHRH and ghrelin in humans. J Clin Endocrinol Metab 2002 ; 87 : 3783-90.
- 10. Beaumont NJ, Skinner VO, Tan TM, et al. Ghrelin can bind to a species of high density lipoprotein associated with paraoxonase. J Biol Chem 2003 ; 278 : 8877-80.
- 11. De Vriese C, Gregoire F, Lema-Kisoka R, et al. Ghrelin degradation by serum and tissue homogenates : identification of the cleavage sites. Endocrinology 2004 ; 145 : 4997-5005.
- 12. Bodart V, Febbraio M, Demers A, et al. CD36 mediates the cardiovascular action of growth hormone-releasing peptides in the heart. Circ Res 2002 ; 90 : 844-9.
- 13. Frieboes RM, Antonijevic IA, Held K, et al. Hexarelin decreases slow-wave sleep and stimulates the secretion of GH, ACTH, cortisol and prolactin during sleep in healthy volunteers. Psychoneuroendocrinology 2004 ; 29 : 851-60.
- 14. Wren AM, Seal LJ, Cohen MA, et al. Ghrelin enhances appetite and increases food intake in humans. J Clin Endocrinol Metab 2001 ; 86 : 5992-95.
- 15. Cowley MA, Smith RG, Diano S, et al. The distribution and mechanism of action of ghrelin in the CNS demonstrates a novel hypothalamic circuit regulating energy homeostasis. Neuron 2003 ; 37 : 649-61.
- 16. Wortley KE, Anderson KD, Garcia K, et al. Genetic deletion of ghrelin does not decrease food intake but influences metabolic fuel preference. Proc Natl Acad Sci USA 2004 ; 101 : 8227-32.
- 17. Tannenbaum GS, Epelbaum J, Bowers CY. Interrelationship between the novel peptide ghrelin and somatostatin/growth hormone-releasing hormone in regulation of pulsatile growth hormone secretion. Endocrinology 2003 ; 144 : 967-74.
- 18. Nakazato M, Murakami N, Date Y, et al. A role for ghrelin in the central regulation of feeding. Nature 2001 ; 409 : 194-8.
- 19. Tolle V, Zizzari P, Tomasetto C, et al.In vivo and in vitro effects of ghrelin/motilin-related peptide on growth hormone secretion in the rat. Neuroendocrinology 2001 ; 73 : 54-61.
- 20. Date Y, Murakami N, Toshinai K, et al. The role of the gastric afferent vagal nerve in ghrelin-induced feeding and growth hormone secretion in rats. Gastroenterology 2002 ; 123 : 1120-8.
- 21. Tschop M, Smiley DL, Heiman ML, et al. Ghrelin induces adiposity in rodents. Nature 2000 ; 407 : 908-13.
- 22. Thompson NM, Gill DA, Davies R, et al. Ghrelin and des-octanoyl ghrelin promote adipogenesis directly in vivo by a mechanism independent of the type 1a growth hormone secretagogue receptor. Endocrinology 2004 ; 145 : 234-42.
- 23. Sun Y, Ahmed S, Smith RG. Deletion of ghrelin impairs neither growth nor appetite. Mol Cell Biol 2003 ; 23 : 7973-81.
- 24. Sun Y, Wang P, Zheng H, Smith RG. Ghrelin stimulation of growth hormone release and appetite is mediated through the growth hormone secretagogue receptor. Proc Natl Acad Sci USA 2004 ; 30 : 101 : 4679-84
- 25. Masuda Y, Tanaka T, Inomata N, et al. Ghrelin stimulates gastric acid secretion and motility in rats. Biochem Biophys Res Commun 2000 ; 276 : 905-8.
- 26. Fujino K, Inui A, Asakawa A, et al. Ghrelin induces fasted motor activity of the gastrointestinal tract in conscious fed rats. J Physiol 2003 ; 550 : 227-40.
- 27. Trudel L, Tomasetto C, Rio MC, et al. Ghrelin/motilin-related peptide is a potent prokinetic to reverse gastric postoperative ileus in rat. Am J Physiol Gastrointest Liver Physiol 2002 ; 282 : G948-52.
- 28. Sibilia V, Rindi G, Pagani F, et al. Ghrelin protects against ethanol-induced gastric ulcers in rats : studies on the mechanisms of action. Endocrinology 2003 ; 144 : 353-9.
- 29. Tatsuguchi A, Miyake K, Gudis K, et al. Effect of Helicobacter pylori infection on ghrelin expression in human gastric mucosa. Am J Gastroenterol 2004 ; 99 : 2121-7.
- 30. Zhang W, Chen M, Chen X, et al. Inhibition of pancreatic protein secretion by ghrelin in the rat. J Physiol 2001 ; 537 : 231-6.
- 31. Arosio M, Ronchi CL, Gebbia C, et al. Stimulatory effects of ghrelin on circulating somatostatin and pancreatic polypeptide levels. J Clin Endocrinol Metab 2003 ; 88 : 701-4.
- 32. Broglio F, Gottero C, Prodam F, et al. Non-acylated ghrelin counteracts the metabolic but not the neuroendocrine response to acylated ghrelin in humans. J Clin Endocrinol Metab 2004 ; 89 : 3062-5.
- 33. Torsello A, Bresciani E, Rossoni G, et al. Ghrelin plays a minor role in the physiological control of cardiac function in the rat. Endocrinology 2003 ; 144 : 1787-92.
- 34. Bisi G, Podio V, Valetto MR, et al. Cardiac effects of hexarelin in hypopituitary adults. Eur J Pharmacol 1999 ; 381 : 31-8.
- 35. Korbonits M, Goldstone AP, Gueorguiev M, et al. Ghrelin. A hormone with multiple functions. Front Neuroendocrinol 2004 ; 25 : 27-68.
- 36. Iglesias MJ, Pineiro R, Blanco M, et al. Growth hormone releasing peptide (ghrelin) is synthesized and secreted by cardiomyocytes. Cardiovasc Res 2004 ; 62 : 481-8.
- 37. Tolle V, Bassant MH, Zizzari P, et al. Ultradian rhythmicity of ghrelin secretion in relation with GH, feeding behavior, and sleep-wake patterns in rats. Endocrinology 2002 ; 143 : 1353-61.
- 38. Corbetta S, Peracchi M, Cappiello V, et al. Circulating ghrelin levels in patients with pancreatic and gastrointestinal neuroendocrine tumors : identification of one pancreatic ghrelinoma. J Clin Endocrinol Metab 2003 ; 88 : 3117-20.
- 39. Camina JP, Carreira MC, El Messari S, et al. Desensitization and endocytosis mechanisms of ghrelin-activated growth hormone secretagogue receptor 1a. Endocrinology 2004 ; 145 : 930-40.
- 40. Cummings DE, Frayo RS, Marmonier C, et al. Plasma ghrelin levels and hunger scores in humans initiating meals voluntarily without time- and food-related cues. Am J Physiol Endocrinol Metab 2004 ; 287 : E297-304.
- 41. Hansen TK, Dall R, Hosoda H, et al. Weight loss increases circulating levels of ghrelin in human obesity. Clin Endocrinol 2002 ; 56 : 203-6
- 42. Tolle V, Kadem M, Bluet-Pajot MT, et al. Balance in ghrelin and leptin plasma levels in anorexia nervosa patients and constitutionally thin women. J Clin Endocrinol Metab 2003 ; 88 : 109-16.
- 43. Poykko SM, Kellokoski E, Horkko S, et al. Low plasma ghrelin is associated with insulin resistance, hypertension, and the prevalence of type 2 diabetes. Diabetes 2003 ; 52 : 2546-53.
- 44. Shiiya T, Nakazato M, Mizuta M, et al. Plasma ghrelin levels in lean and obese humans and the effect of glucose on ghrelin secretion. J Clin Endocrinol Metab 2002 ; 87 : 240-4.
- 45. Pombo M, Leal-Cerro A, Barreiro J, et al. Growth hormone releasing hexapeptide-6 (GHRP-6) test in the diagnosis of GH-deficiency. J Pediatr Endocrinol Metab 1996 ; 9 : 333-8.
- 46. Tan TM, Vanderpump M, Khoo B, et al. Somatostatin infusion lowers plasma ghrelin without reducing appetite in adults with Prader-Willi syndrome. J Clin Endocrinol Metab 2004 ; 89 : 4162-5.
- 47. Pantel J, Cabrol S, Nivot S, et al.Familial short stature associated with a mutation in the growth hormone secretagogue receptor (GHSR), 86th Annual meeting of the Endocrine Society. New Orleans : Endocrine Society, 2004 : O10-6 (abstract).
- 48. Ariyasu H, Takaya K, Iwakura H, et al. Transgenic mice overexpressing des-acyl ghrelin show small phenotype. Endocrinology 2005 ; 146 : 355-64.
- 49. Shuto Y, Shibasaki T, Otagiri A, et al. Hypothalamic growth hormone secretagogue receptor regulates growth hormone secretion, feeding, and adiposity. J Clin Invest 2002 ; 109 : 1429-36.
- 50. Lall S, Balthasar N, Carmignac D, et al. Physiological studies of transgenic mice overexpressing growth hormone (GH) secretagogue receptor 1A in GH-releasing hormone neurons. Endocrinology 2004 ; 145 : 1602-11.
- 51. Smith RG, Cheng K, Schoen WR, et al. A nonpeptidyl growth hormone secretagogue. Science 1993 ; 260 : 1640-3.
List of figures
Figure 1
Ghréline et autres sécrétagogues de la GH (GHS), et leur récepteur GHS-R 1a.
Figure 2
Effets pléïotropes de la ghréline et des sécrétagogues de la GH.
L’acylation de la ghréline est nécessaire aux effets centraux, aux sécrétions hypophysaires, à la prise alimentaire et à certains effets métaboliques et gastro-intestinaux. La forme non acylée est impliquée dans l’adipogenèse, les effets cardiovasculaires et les phénomènes de prolifération/différenciation.
List of tables
Tableau I
Récepteurs et ligands.
L’adénosine ne stimule pas la sécrétion de GH, la cortistatine atténue les effets de la ghréline. EP 51389 et TH 2306.34 stimulent la sécrétion de GH, BIM 28131 augmente la prise de poids, TZP 101 stimule la vidange gastrique. BIM 28163 inhibe l’effet de la ghréline sur la GH, mais augmente la prise de poids (il agit donc également par l’intermédiaire d’un autre récepteur). L’hexaréline, via CD36, prévient la formation d’athérosclérose.
Tableau II
Modèles animaux d’invalidation ou de surexpression de la ghréline et du récepteur GHS-R.
Tableau III
Non-corrélation entre valeurs circulantes de GH et de ghréline, en relation avec la prise alimentaire, dans différentes conditions physiopathologiques.
Dans le cas de l’obésité, des modifications non univoques de l’appétit et des préférences alimentaires sont observées. Un seul cas de ghrélinome a été décrit dans la littérature, et le comportement alimentaire de ce patient n’a pas été étudié.