Article body
Le récepteur de chimiokines CCR5 est utilisé par le virus de l'immunodéficience humaine de type 1 (VIH-1) pour pénétrer dans les cellules du système immunitaire. Les porteurs de la mutation CCR5Δ32 sont naturellement résistants au VIH-1 [1] car la délétion de 32 paires de bases dans le gène de ce corécepteur empêche le virus de pénétrer dans les cellules cibles [2] (Figures 1 et 2).
Figure 1
La mutation Δ32 se traduit par la délétion de 32 paires de bases, ce qui introduit un décalage dans le cadre de lecture et conduit à l'arrêt prématuré de la synthèse du récepteur CCR5.

Figure 2
Trois des sept domaines transmembranaires sont absents dans la protéine CCR5Δ32.
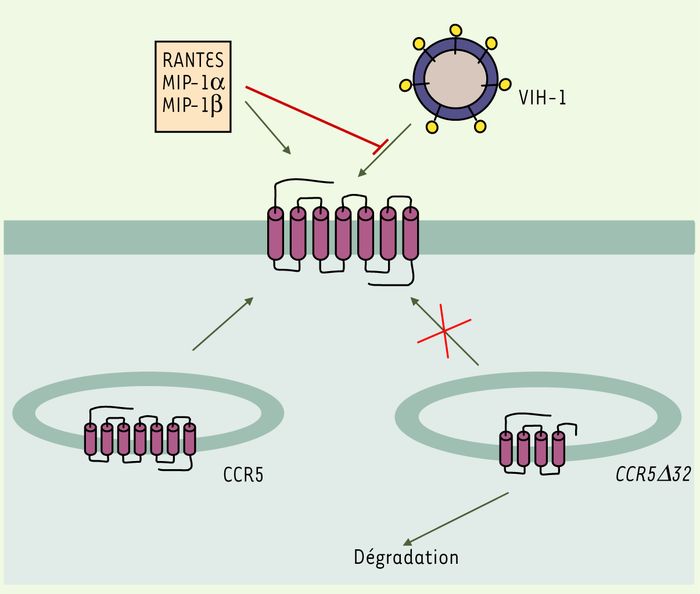
Il en résulte que la protéine est mal repliée et se dégrade, au lieu d'être transportée dans la membrane plasmique. MIP-1α, MIP-1β : macrophage inflammatory protein 1α, β ; RANTES : chimiokine.
La fréquence et la distribution du génotype CCR5Δ32 dans la population suggèrent une sélection récente. Les individus homozygotes CCR5Δ32/CCR5Δ32 ne se distinguent par aucun phénotype autre que leur apparente résistance au VIH-1. De façon surprenante, le génotype CCR5Δ32 n'est présent exclusivement que dans la population caucasienne : la fréquence moyenne est de 10 % (donc 1 % d'homozygotes) et va jusqu'à 16 % dans les populations de Finlande et de Russie. Ces observations permettent donc de supposer que l'émergence de la mutation CCR5Δ32 s'est produite en Europe assez récemment. Elle résulterait d'une sélection positive dont la cause fait l'objet d'une hypothèse audacieuse formulée par des chercheurs britanniques dans un article publié dans le Journal of Medical Genetics en 2005 [3].
Hypothèses sur la nature de la pression sélective favorisant CCR5Δ32
Selon les estimations, la peste noire tua 40 % de la population européenne au xive siècle, et continua de sévir par flambées récurrentes sporadiques pendant 400 ans. Une autre hypothèse, soutenue par un modèle mathématique publié en 2003 par Alison P. Galvani et Montgomery Slatkin, désigne la variole comme origine probable de la pression de sélection en faveur de CCR5Δ32 [4]. Causée par un virus de la famille des Poxviridae, la variole a provoqué des épidémies importantes en Europe au xviie et xviiie siècles. La distribution de la variole représenterait mieux, selon les auteurs, le gradient de distribution nord-est/sud-ouest de CCR5Δ32 en Europe que la peste, qui, elle, aurait sévi plus fortement en Europe centrale. De plus, le rôle facilitateur des récepteurs de chimiokines pour l'entrée virale de certains poxvirus dans des cellules cibles a certes été suggéré, mais uniquement dans un rapport isolé en 1999 [5]. En revanche, l'analyse de souris dont le gène ccr5 est inactivé a donné des résultats ambigus sur une éventuelle implication de la bactérie Yersinia pestis, l'agent de la peste. En effet, la délétion de ccr5 n'affecte pas les taux de survie après inoculation à dose létale [6, 7], même si l'absence du récepteur semble réduire le taux de phagocytose [6]. L'ensemble de ces données serait donc en faveur des vagues de variole à l'origine de la sélection positive de CCR5Δ32, plutôt que des épidémies de peste.
Mais cette hypothèse est actuellement remise en question par un travail de réévaluation des pressions sélectives historiques exercées par les différentes épidémies lors des siècles passés. Un groupe de chercheurs de l'Université de Liverpool (Royaume-Uni) avance que la durée des épidémies de variole et leur taux de mortalité, supposés par Alison P. Galvani et Montgomery Slatkin pour leur modèle de simulation mathématique, ne correspondraient pas aux données épidémiologiques documentées recueillies à partir d'archives [3]. Selon Christopher Duncan et ses collègues, la sélection initiale serait due aux épidémies documentées, rapportées sous le nom de « pestes ». Mais ces infections seraient pour la plupart des fièvres hémorragiques d'origine virale, et appelées « pestes hémorragiques ». Ce serait alors un virus (non identifié) déclencheur de la peste hémorragique, et non Yersinia pestis, qui aurait causé la plupart des « pestes » épidémiques européennes et qui serait à l'origine de la sélection de CCR5Δ32. Le virus aurait utilisé le récepteur des chimiokines CCR5 comme porte d'entrée pour atteindre ses cibles, tout comme le VIH, épargnant alors les porteurs de CCR5Δ32 et favorisant ainsi la fréquence du gène mutant. L'importance du virus de la variole se limiterait donc au maintien d'un taux élevé de la mutation dans la population caucasienne après l'extinction des pestes hémorragiques.
Pour stimulante qu'elle soit, l'hypothèse de C. Duncan et al. à la recherche des causes de la fréquence de CCR5Δ32 risque de rester controversée. Car supposer que certaines des épidémies de peste historiquement célèbres aient pu être provoquées par d'autres agents que Yersinia demeure sujet à débats [8], d'autant plus que des traces d'ADN de Yersinia ont été trouvées dans la pulpe dentaire d'ossements humains provenant de nécropoles où furent rassemblées des victimes de la peste en France et en Allemagne [9, 10]. En outre, les fièvres hémorragiques historiques, données en exemple par les auteurs, ne sont pas confinées à l'Europe mais ont frappé l'Égypte pharaonique, la Mésopotamie, les Empires byzantin et islamique. De surcroît, on ne connaît aucun exemple de virus, agent de fièvre hémorragique, ayant accès aux cellules cibles par l'intermédiaire de récepteurs des chimiokines. On peut, par ailleurs, s'interroger sur le rôle de CCR5 dans l'épidémie ayant imposé la sélection positive de CCR5Δ32 : se limiterait-il, comme pour le VIH, à l'entrée du pathogène dans des cellules cibles, ou ne pourrait-il pas également contribuer à la montée d'une réaction immunitaire exagérée ou inefficace en réponse à une infection, quel que soit l'agent infectieux, ce qui influencerait le taux de mortalité ?
Enfin, une nouvelle publication allemande semble doublement réfuter le rôle de Yersinia pestis, du virus de la variole et du virus évoqué par les Britanniques [11]. D'une part, en analysant des squelettes d'anciens cimetières allemands et italiens, il apparaît que la fréquence de CCR5Δ32 n'est pas moins importante parmi les victimes de la peste que parmi des victimes de famines. D'autre part, la mutation CCR5Δ32 était déjà prévalente dans des groupes humains beaucoup plus anciens, provenant de l'âge de bronze.
On le voit, la plus grande fréquence de CCR5Δ32 dans les populations européennes n'a pas encore trouvé son explication.
Appendices
Références
- 1. Dragic T, Litwin V, Allaway GP, et al. HIV-1 entry into CD4+ cells is mediated by the chemokine receptor CC-CKR-5. Nature 1996 ; 381 : 667-73.
- 2. Liu R, Paxton WA, Choe S, et al. Homozygous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell 1996 ; 86 : 367-77.
- 3. Duncan SR, Scott S, Duncan CJ. Reappraisal of the historical selective pressures for the CCR5Δ32 mutation. J Med Genet 2005 ; 42 : 205-8.
- 4. Galvani AP, Slatkin M. Evaluating plague and smallpox as historical selective pressures for the CCR5-Delta 32 HIV-resistance allele. Proc Natl Acad Sci USA 2003 ; 100 : 15276-9.
- 5. Lalani AS, Masters J, Zeng W, et al. Use of chemokine receptors by poxviruses. Science 1999 ; 286 : 1968-71.
- 6. Elvin SJ, Williamson ED, Scott JC, et al. Evolutionary genetics : ambiguous role of CCR5 in Y. pestis infection. Nature 2004 ; 430 : 417.
- 7. Mecsas J, Franklin G, Kuziel WA, et al. Evolutionary genetics : CCR5 mutation and plague protection. Nature 2004 ; 427 : 606.
- 8. Paterson R. Yersinia seeks pardon for black death. Lancet Infect Dis 2002 ; 2 : 323.
- 9. Raoult D, Aboudharam G, Crubezy E, et al. Molecular identification by suicide PCR of Yersinia pestis as the agent of medieval black death. Proc Natl Acad Sci USA 2000 ; 97 : 12800-3.
- 10. Wiechmann I, Grupe G. Detection of Yersinia pestis DNA in two early medieval skeletal finds from Aschheim (Upper Bavaria, 6th century AD). Am J Phys Anthropol 2005 ; 126 : 48-55.
- 11. Hummel S, Schmidt D, Kremeyer B, et al. Detection of the CCR5Δ32 HIV resistance gene in bronze age skeletons. Genes Immun 2005 online.
List of figures
Figure 1
La mutation Δ32 se traduit par la délétion de 32 paires de bases, ce qui introduit un décalage dans le cadre de lecture et conduit à l'arrêt prématuré de la synthèse du récepteur CCR5.
Figure 2
Trois des sept domaines transmembranaires sont absents dans la protéine CCR5Δ32.
Il en résulte que la protéine est mal repliée et se dégrade, au lieu d'être transportée dans la membrane plasmique. MIP-1α, MIP-1β : macrophage inflammatory protein 1α, β ; RANTES : chimiokine.