Article body
Les séquences d’ADN répétitives adjacentes aux centromères sont assemblées sous une forme spéciale de chromatine, l’hétérochromatine péricentrique, qui est généralement considérée comme une région chromosomique inerte. Cette réputation lui vient en partie du fait qu’elle impose un silence transcriptionnel aux gènes qui y sont relocalisés en raison de translocations chromosomiques. L’état de ces gènes maintenus réprimés sous l’influence de l’hétérochromatine est transmis de façon stable à travers la mitose et la méiose. En accord avec ce caractère inerte et hostile à la transcription, l’hétérochromatine péricentrique demeure condensée et relativement inaccessible aux nucléases pendant tout le cycle cellulaire.
Les protéines HP1 (heterochromatin protein 1) sont une des composantes essentielles de l’hétérochromatine péricentrique. Ces protéines sont dépourvues de domaine de liaison à l’ADN. Leur association avec l’hétérochromatine dépend de leur domaine amino-terminal (chromo) qui se fixe de façon très stable (Kd = 2μM) à l’histone H3 méthylée sur le groupement ε-aminé de la lysine 9 (Lys 9) [1]. La présence d’un domaine de dimérisation dans sa partie carboxy-terminale (shadow chromo) permet à HP1 de fixer simultanément deux molécules d’H3 méthylées sur la Lys9 de manière intra- ou inter-nucléosomique (Figure 1A) ((→) m/s 2002, n°1, p.17). Leur structure dimérique confère donc aux protéines HP1 le potentiel de compacter la chromatine en joignant des nucléosomes qui ne sont pas nécessairement adjacents le long de la fibre de chromatine. La capacité de ponter deux fibres de chromatine distinctes pourrait aussi être à l’origine de la relocalisation de certains gènes au sein de l’hétérochromatine péricentrique dans les cellules où ces gènes sont sujets à une répression transcriptionnelle [2]. Cette propriété des protéines HP1 pourrait aussi servir à renforcer la cohésion pericentromérique pour résister aux forces de tension du fuseau mitotique et empêcher la dissociation prématurée des chromatides soeurs durant la mitose [3].
Figure 1
Association des protéines HP1 avec les histones de l’hétérochromatine péricentrique.
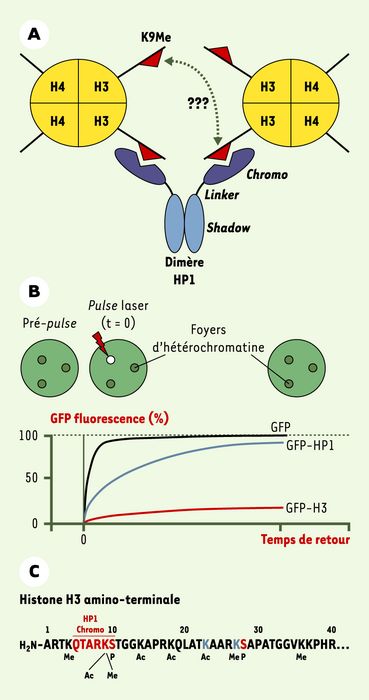
A. La protéine dimérique HP1 a le potentiel de se fixer, de manière intra- ou inter-nucléosomale, à deux molécules d’histone H3 méthylées sur la lysine 9 (K9).B. Technique utilisée pour mesurer la dynamique des protéines HP1 et des histones au niveau des foyers d’hétérochromatine. C. Séquence de la région amino-terminale de l’histone H3 (Ac = acétylation, Me = méthylation, P = phosphorylation). Certaines modifications, par exemple l’acétylation et la méthylation de la lysine 9, sont mutuellement exclusives et doivent nécessairement se retrouver dans des molécules d’H3 distinctes. La protéine HP1 se fixe sur une courte région d’H3 qui inclut la lysine K9 méthylée par Suv39h1/h2. Ce résidu est adjacent à la sérine 10 phosphorylée en mitose par les kinases Aurora.
Contre toute attente, deux études récentes révèlent que les protéines HP1 sont en fait associées de façon très dynamique avec l’hétérochromatine péricentrique [4, 5]. Ces études font appel à une technique de biologie cellulaire qui permet de mesurer la mobilité de protéines de fusion GFP (green fluorescent protein) in vivo. Cette technique, appelée «retour de fluorescence après photoblanchiment» (fluorescence recovery after photobleaching, FRAP), utilise un pulse laser pour éliminer la fluorescence provenant de molécules GFP-HP1 présentes dans une région restreinte de l’hétérochromatine (Figure 1B, cercle blanc). On mesure alors le temps de retour de la fluorescence qui est fonction du temps nécessaire pour dissocier les molécules GFP-HP1 dont le chromophore GFP a été détruit par le pulse laser et les remplacer par de nouvelles molécules GFP-HP1 provenant du nucléoplasme (Figure 1B). Ces études montrent que les protéines HP1, bien qu’elles soient moins mobiles que la GFP libre, s’associent à l’hétérochromatine de façon nettement plus dynamique que les histones H3 ou H4. Par exemple, alors que le temps de retour de fluorescence pour la majorité des molécules GFP-HP1 (entre 70 et 100 %) est d’environ 1-3 minutes [4, 5], une fraction importante de la fluorescence des molécules d’histone H3 ou H4-GFP (70%) est perdue de façon irréversible après photoblanchiment (temps de retour > 8heures) [6].
Il est donc clair que l’hétérochromatine doit désormais être considérée comme une structure dont l’une des composantes majeures, HP1, est en équilibre dynamique avec le nucléoplasme. Il est possible de réconcilier cette propriété avec la nature quasi statique de l’hétérochromatine en suggérant que, malgré la dissociation continuelle d’une fraction des molécules HP1, la structure globale est maintenue de façon stable. La modification de l’histone H3 sur la Lys9 au niveau de l’hétérochromatine est établie par les histone méthyltransférases (Suv39h1 et Suv39h2). Par conséquent, l’association des protéines HP1 avec l’hétérochromatine est abolie dans les cellules où ces deux enzymes sont absentes [5]. Contrairement à l’acétylation qui est sujette à un renouvellement très rapide grâce aux actions opposées des histone acétyltransférases et des désacétylases, la méthylation des histones est, quant à elle, une modification très stable [7]. Si la méthylation de la Lys9 d’H3 est effectivement irréversible, l’association permanente de cette histone avec l’hétérochromatine joue vraisemblablement un rôle important pour permettre aux protéines HP1 de retourner rapidement aux sites laissés vacants afin d’y rétablir la structure initiale.
L’association dynamique des protéines HP1 a le pouvoir d’altérer la condensation de la fibre de chromatine de façon transitoire et localisée. Cette décondensation pourrait faciliter l’action de plusieurs enzymes qui modulent la structure de l’hétérochromatine à différentes étapes du cycle cellulaire. Par exemple, les histones néo-synthétisées déposées durant la réplication de l’hétérochromatine sont initialement acétylées et dépourvues de méthylation sur la Lys9. Leur désacétylation et leur méthylation à chaque cycle cellulaire sont nécessaires pour maintenir l’intégrité de l’hétérochromatine péricentrique et prévenir les erreurs de ségrégation chromosomique [8]. La phosphorylation de l’histone H3 sur la sérine 10 (Ser10) par les kinases Aurora est elle aussi impliquée dans la ségrégation des chromosomes [9]. Cette phosphorylation est amorcée en phase G2 au niveau de l’hétérochromatine péricentrique, d’où elle se propage pour éventuellement couvrir l’ensemble du génome en prophase. La Ser10 de l’histone H3 est immédiatement adjacente à la Lys9 et la phosphorylation de la Ser10 par les kinases Aurora est partiellement bloquée in vitro par la présence de méthylation sur la Lys9 [10]. La fixation des protéines HP1 sur cette région d’H3 risque de renforcer cette inhibition (Figure 1C). Il devient donc important de déterminer si ces deux modifications post-traductionnelles de l’histone H3 (méthylation de la Lys9 et phosphorylation de la Ser10) se retrouvent dans les mêmes molécules d’H3 ou dans des molécules d’H3 distinctes. En conclusion, la ségrégation fidèle des chromosomes dépend de plusieurs modifications enzymatiques des histones (désacétylation, méthylation et phosphorylation) qui doivent avoir lieu dans un environnement particulièrement hostile, l’hétérochromatine péricentrique. Il est probable que ces modifications enzymatiques des histones sont facilitées par le comportement dynamique des protéines HP1.
Appendices
Références
- 1. Nielsen PR, Nietlispach D, Mott HR, et al. Structure of the HP1 chromodomain bound to histone H3 methylated at lysine 9. Nature 2002; 416: 103-7.
- 2. Azuara V, Brown KE, Williams RRE, et al. Heritable gene silencing in lymphocytes delays chromatin resolution without affecting the timing of DNA replication. Nat Cell Biol 2003; 5: 668-74.
- 3. Bernard P, Maure JF, Partridge JF, Genier S, Javerzat JP, Allshire RC. Requirement of heterochromatin for cohesion at centromeres. Science 2001; 294: 2539-42.
- 4. Festenstein R, Pagakis SN, Hiragami K, et al. Modulation of heterochromatin protein 1 dynamics in primary mammalian cells. Science 2003; 299: 719-21.
- 5. Cheutin T, McNairn AJ, Jenuwein T, Gilbert DM, Singh PB, Misteli T. Maintenance of stable heterochromatin domains by dynamic HP1 binding. Science 2003; 299: 721-5.
- 6. Kimura H, Cook PR. Kinetics of core histones in living human cells: little exchange of H3 and H4 and some rapid exchange of H2B. J Cell Biol 2001;153: 1341-53.
- 7. Bannister AJ, Schneider R, Kouzarides T. Histone methylation: dynamic or static? Cell 2002; 109: 801-6.
- 8. Taddei A, Maison C, Roche D, Almouzni G. Reversible disruption of pericentric heterochromatin and centromere function by inhibiting deacetylases. Nat Cell Biol 2001; 3: 114-20.
- 9. Hans F, Dimitrov S. Histone H3 phosphorylation and cell division. Oncogene 2001; 20: 3021-7.
- 10. Rea S, Eisenhaber F, O’Carroll D, et al. Regulation of chromatin structure by site-specific histone H3 methyltransferases. Nature 2000; 406: 593-9.
List of figures
Figure 1
Association des protéines HP1 avec les histones de l’hétérochromatine péricentrique.
A. La protéine dimérique HP1 a le potentiel de se fixer, de manière intra- ou inter-nucléosomale, à deux molécules d’histone H3 méthylées sur la lysine 9 (K9).B. Technique utilisée pour mesurer la dynamique des protéines HP1 et des histones au niveau des foyers d’hétérochromatine. C. Séquence de la région amino-terminale de l’histone H3 (Ac = acétylation, Me = méthylation, P = phosphorylation). Certaines modifications, par exemple l’acétylation et la méthylation de la lysine 9, sont mutuellement exclusives et doivent nécessairement se retrouver dans des molécules d’H3 distinctes. La protéine HP1 se fixe sur une courte région d’H3 qui inclut la lysine K9 méthylée par Suv39h1/h2. Ce résidu est adjacent à la sérine 10 phosphorylée en mitose par les kinases Aurora.