Abstracts
Abstract
A plant macrofossil record from the glacial Lake Hind basin is used to reconstruct early postglacial wetland plant succession and paleohydrology. Between >10.6 and 9.1 ka BP, there are four plant assemblage zones: (1) an early (>10.6 ka BP) zone dominated by Cyperaceae and aquatics; (2) a subsequent zone (~10.6-10.1 ka BP) with emergents (Menyanthes trifoliata, Potentilla palustris, Scirpus validus) and fewer aquatic plants; (3) an interval between ~10.1 and 9.8 ka BP dominated by Drepanocladus aduncus; and (4) a zone between ~9.8 and 9.1 ka BP with Menyanthes trifoliata and Equisetum. These data indicate a gradual decline in water depth between 10.6 and 10.1 ka BP due to deepening of one or more outlets of glacial Lake Hind. From ~10.6 to 9.1 ka BP, the importance of Menyanthes records pronounced, seasonal, flooding. Furthermore, lack of evidence for complete drawdown and terrestrialization in the basin – despite local and regional evidence for postglacial warming – indicates that this wetland was minimally impacted by climate change up to at least 9.1 ka BP. Persistence of very wet conditions locally is consistent with recent results from south-central Saskatchewan, and may be due to release of meltwater from stagnant ice. However, frequent low-energy flooding of the basin by the Souris River is more plausible. In general, the apparent insensitivity of aquatic habitats to abrupt climate change in some locales on the Canadian Prairies demonstrates the potential long-term mitigating effects of local hydrological factors.
Résumé
L’analyse de macrofossiles de plantes dans la cuvette du Lac glaciaire Hind a servi à déterminer la paléohydrologie et la succession de plantes d’un milieu humide du début du Postglaciaire. Entre >10,6 et 9,1 ka BP, quatre zones d’assemblages se succèdent : (1) une première zone (>10,6 ka BP) dominée par les Cyperacées et les plantes aquatiques ; (2) une zone subséquente (~10,6-10,1 ka BP) caractérisée par quelques plantes émergentes (Menyanthes trifoliata, Potentilla palustris, Scirpus validus) et certaines plantes aquatiques ; (3) une troisième zone (~10,1-9,8 ka BP) dominée par Drepanocladus aduncus ; et (4) une dernière zone (~9,8-9,1 ka BP) composée de Menyanthes trifoliata et d’Equisetum. Selon les données, la profondeur d’eau de la cuvette du Lac glaciaire Hind a diminué de façon progressive entre 10,6 et 10,1 ka BP à la suite du surcreusement d’un ou de plusieurs de ses exutoires. De 10,6 à 9,1 ka BP, l’importance de Menyanthes témoigne d’importants débordements saisonniers des eaux. De plus, l'absence d’indices d’un assèchement total de la cuvette (malgré le réchauffement postglaciaire local et régional) indique que cette zone humide a résisté au changement climatique, au moins jusqu’à 9,1 ka BP. La persistance de conditions localement très humides concorde avec de récents résultats provenant du centre-sud de la Saskatchewan et pourrait être attribuable à la fonte de glace stagnante. Cependant, l'hypothèse de débordements fréquents mais modérés de la rivière Souris dans la cuvette est plus plausible. En général, l'apparente résistance, dans certains secteurs des Prairies, des habitats aquatiques aux changements climatiques brutaux démontre bien le potentiel à long terme des effets atténuants des facteurs hydrologiques locaux.
Resumen
El registro macrofósil vegetal de la cuenca del Lago glaciar Hind fue usado para reconstruir la sucesión vegetal y la paleohidrología de la zona húmeda. En un periodo situado hace unos 10 600 a 9100 años se distinguen cuatro zonas vegetales : la primera zona (hace mas de 10 600 años) dominada por Cyperaceae y acuáticas ; la segunda datando de aproximadamente unos 10 600 a 10 100 años, representada por plantas emergentes (Menyanthes trifoliata, Potentilla palustris, Scirpus validus) y unas pocas plantas acuáticas ; una tercera situada en el intervalo cubierto entre unos 10 100 y 9800 años, dominada por Drepanocladus aduncus; y la última que abarca el periodo comprendido entre unos 9800 y 9100 años, representada por Menyanthes trifoliata y Equisetum. Estos datos indican una disminución gradual de la profundidad del lago debido a la excavación de uno o varios de los afluentes del lago glaciar Hind y que la sitúa hace aproximadamente 10 600 a 10 100 años. Alrededor del periodo comprendido entre unos 10 600 y 9100 años, los registros obtenidos de Menyanthes reflejan episodios importantes y estacionales de inundaciones. Mas aun, la carencia de evidencia de un descenso del nivel y de la sedimentación de la cuenca – a pesar de la evidencia local y regional de calentamiento postglaciar – indica que esta zona húmeda fue poco perturbada por el cambio climático hasta hace menos de 9100 años. La persistencia de condiciones muy húmedas de la localidad concuerda con los resultados recientes de la zona centro-sur de Saskatchewan, y puede deberse a la liberación del agua de fusión proveniente del hielo estancado. Sin embargo, es mas probable que se trate de las inundaciones frecuentes de baja energía que ocurrieron en la cuenca provocadas por el Souris River. En general, la aparente resistencia de los ambientes acuáticos frente a cambios abruptos del clima en algunas localidades de la Praderas canadienses demuestra el potencial a largo plazo del efecto atenuante de los factores hídricos locales.
Article body
Introduction
Multi-decadal to century-scale responses of wetlands to periods of abrupt climate change (Yu and Wright, 2001) are not well understood on the Canadian Prairies (Wiken, 1986). Beyond the range of modern observation, dynamic cause-and-effect connections between vegetation, hydrology, and climate change at this temporal scale can only be investigated through detailed analysis of the paleoecological record. Plant macrofossils are particularly useful in this respect because, unlike pollen, they are typically deposited in situ, and can be often identified to species (Birks, 1973). Thus, the macrobotanical record permits reliable and precise local-scale models of past hydrologic and biotic responses to climate change.
The early postglacial period in North America displays climatic oscillations caused by earth-surface and solar forcing mechanisms (summarized in Yu and Wright, 2001). Of particular interest for this paper is the well-documented period of abrupt warming and drying beginning ~10 ka BP (uncalibrated), but prior to the brief Pre-Boreal Oscillation (PBO) at 9.65 ka BP. There has been little research into the responses of wetland ecosystems to this warm episode on the northern Prairies. In part, this situation stems from a general lack of perennial lakes with continuous paleoecological records in this region, resulting in few detailed macrobotanical studies. In many lakes on the Prairies, episodes of prolonged aridity before the late Holocene caused these basins to dry up, and the compacted sediments formed during these events have rendered recovery of underlying early Holocene materials difficult (Barnosky et al., 1987; Beaudoin, 1993; Vance et al., 1993). Recently, however, it has been shown that non-traditional paleoecological sites such as (now dry) kettle-fill sites on the Canadian Prairies are sometimes excellent repositories of early Holocene plant remains (Yansa, 1998; Yansa and Basinger, 1999). Consequently, these sites offer an excellent opportunity to examine past ecosystem processes during the early postglacial period.
The study site (Flintstone Hill) is located in the glacial Lake Hind basin, southwestern Manitoba, Canada. Today, the Hind basin is underlain by a large groundwater system (the Oak Lake aquifer). Between ~10.6 to 9.1 ka BP, following the regression of glacial Lake Hind, aquatic and emergent plants colonized shallow wetlands in the basin (Boyd et al., 2003). Because water depth, amplitude of water level fluctuations, water chemistry, and other hydrologic parameters are important factors that determine the wetland plant assemblage present at a site (Warner, 1990), changes in the macrobotanical record permit detailed reconstruction of hydrologic regimes through time. With this approach, I reconstruct the relationship between hydrology, wetland plant succession, and early postglacial warming beginning ~10 ka BP. I compare these results with other early postglacial macrobotanical assemblages from the Canadian Prairies in order to differentiate regional-, from subregional- or local-scale, processes.
Study site
The Flintstone Hill (FSH) site is a cutbank of the Souris River, located in the south-central glacial Lake Hind basin (Figs. 1 and 2). Glacial Lake Hind was one of several interconnected proglacial lakes that formed across the northern Prairies during the period of final (Late Wisconsinan) deglaciation (e.g., Klassen, 1972; Clayton and Moran, 1982; Fenton et al., 1983; Kehew and Clayton, 1983; Klassen, 1983; Kehew and Lord, 1986; Kehew and Teller, 1994; Sun and Teller, 1997). Lake Hind was part of the northern Plains proglacial lake-spillway system. It received meltwater from western Manitoba, Saskatchewan, and North Dakota through at least 10 channels, and discharged eastwards into glacial Lake Agassiz through the Pembina spillway (Sun, 1996; Sun and Teller, 1997). Much of glacial Lake Hind was drained shortly before ~10.4 ka BP (Boyd et al., 2003), due to the catastrophic routing of meltwater from glacial Lake Regina, which deepened and widened the Pembina spillway (Sun, 1996; Sun and Teller, 1997).
The modern landscape of the Hind basin is dominated by ~18 discontinuous dune fields (“Oak Lake dunes” of David, 1977) mostly of late Holocene age (Boyd, 2000; Running et al., 2002; Wallace, 2002). Near Flintstone Hill, the dominant eolian landforms are large parabolic dunes with arms (<10 m high, 500-2 000 m long) that are oriented WNW-ENE. Interdunal swales support shallow wetlands fed by the Oak Lake aquifer. The combination of a high water table and considerable topographic variability sustain forest (Populus-Quercus-Fraxinus), grassland, and wetland plant communities.
Although river cutbanks are unconventional sites for paleoecological analysis, Flintstone Hill exposes a >2 km long lithostratigraphic sequence spanning the terminal Late Pleistocene to the present (Boyd, 2000, 2002; Running et al., 2002; Boyd et al., 2003). The basal unit (A1), the focus of this paper, contains a laminated organic deposit with diverse and well-preserved macrofossils constrained by 14C dates to the interval between >10.6 and 9.1 ka BP. This record has recently been used to address the nature of early Paleoindian land-use in the glacial Lake Hind basin (Boyd et al., 2003). For a detailed discussion of the entire FSH lithostratigraphic sequence, see Boyd (2000) and Running et al. (2002).
The basal, fine-textured sediments exposed at Flintstone Hill are at least 2.25 m thick and extend below the present river level. The lowest unit (A1) grades from a gleyed massive to planar-bedded, carbonate-rich, clay to silty clay upward into a 30 cm thick peaty (organic) deposit with alternating silty clay and organic laminae (Fig. 3). This organic deposit in the upper 30 cm of Unit A1 exhibits progressively finer texture (clay loam and silty clay loam) and thinner planar beds (~1-5 mm thick) upwards. Organic matter (largely plant detritus) also increases upward. Alternating organic and clastic beds are more frequent in the upper 15 cm, and small (<5 mm high), symmetrical ripple structures are preserved in places. Two AMS radiocarbon dates on seeds of the emergent Menyanthes trifoliata L. were obtained from the bottom and top portions of the organic deposit: 10 420 ± 70 BP (12 677-11 991 cal BP) (Beta-116994), and 9250 ± 90 BP (10 596-10 228 cal BP) (TO-7692), respectively (Boyd, 2000; Boyd et al., 2003). Occasional carbonate-rich clastic beds with ripples in this deposit (particularly in the upper 15 cm) indicate periods of low-energy flooding and above-surface water levels. Macrofossils of emergent plants are abundant, suggesting generally low water levels. This unit is capped by ~10 cm of marl (carbonate mud) which overlies the laminated organic deposit and contains few plant remains. The contact between this marl (bottom of Unit A2) and Unit A1 is sharp and the marl exhibits rip-up clasts (mm-1 cm long axes) composed of detritus from the underlying organic deposit. Unit A1 documents the regression of glacial Lake Hind to at least the Phase 9 level (Sun, 1996) before 10.4 ka BP (Fig. 2) (Boyd et al., 2003).
Figure 1
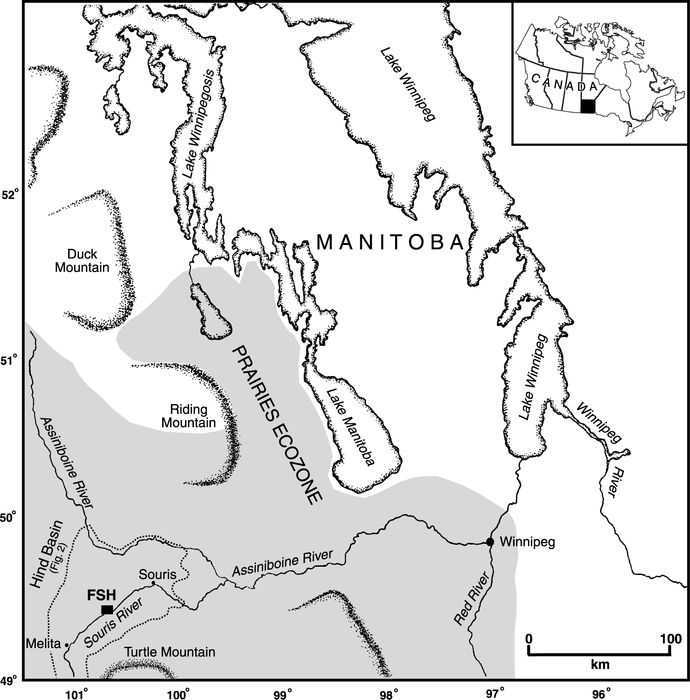
Map of southern Manitoba with the location of glacial Lake Hind, the Flintstone Hill (FSH) study site, southwestern Manitoba, and major physiographic features. Canadian Prairies Ecozone (Wiken, 1986) is shaded in grey.
Carte du sud du Manitoba et localisation du Lac glaciaire Hind, du site d’étude Flintstone Hill (FSH), dans le sud-ouest du Manitoba, et des principaux éléments physiographiques. L’écozone des Prairies (Wiken, 1986) est en gris.
Unit A2 is composed of interbedded fine sand, silt, clay and marl, with an overall higher carbonate content than Unit A1, and abundant gastropod and bivalve shells (Running et al., 2002); pollen and plant macrofossils are rare. This unit coarsens upward to a thinner, discontinuous, organic deposit up to 15 cm thick in a fine sand matrix. A conventional radiocarbon age on wood from this organic deposit places it at 6700 ± 70 BP (Beta-111142; 7667-7462 cal BP) (Boyd, 2000). Unit A2 represents the early Souris River prior to incision that established its present channel through the glacial Lake Hind basin (see Running et al., 2002). The observed distribution of marl, grain sizes and bedding that generally coarsen upward, and greater carbonate content compared to Unit A1, are consistent with low-energy accretion in a position away from the thalweg of the channel (Running et al., 2002). This interpretation is supported by the gastropod and bivalve shell assemblage recovered from Unit A2 deposits at Flintstone Hill (Running et al., 2002). Vertical accretion facies of Unit A2 with cumulic A-horizons are locally observed in other cutbank exposures along the Souris River (Running et al., 2002).
Materials and methods
In 2000, one litre samples were collected for macrobotanical processing. The laminated organic portion of Unit A1 and the underlying massive sediment were sampled completely and continuously, with each sample representing a depth of 2 cm. Samples were excavated directly from the cutbank face, and the depth of sampling was limited by the height of the water table and river. Subsamples of 50 ml were washed through 250 μm and 125 μm sieves, and the detritus was air-dried before examination under a dissecting microscope. A minimum of 150 ml of sediment was processed in this manner per sample, and macrofossil counts were calculated to a fixed volume (150 ml). Plant macrofossil identifications used comparative collections of the University of Manitoba Herbarium, in addition to keys in Berggren (1969), Montgomery (1977), Lévesque et al. (1988), and Martin and Barkley (2000). Macrofossil data were zoned by stratigraphically constrained incremental sum of squares cluster analysis (Euclidean distance dissimilarity coefficient) (Grimm, 1987).
Figure 2

Glacial Lake Hind with location of Flintstone Hill (FSH) study site. Phase 4 to Phase 9 lake shorelines after Sun (1996).
Lac glaciaire Hind et localisation du site d’étude Flintstone Hill (FSH). Phases 4 à 9 du littoral d'après Sun (1996).
Complete pollen analysis of Unit A1 at the FSH site is presented in Boyd (2000) and Boyd et al. (2003). In this study, only the pollen spectrum for Picea glauca (white spruce) is presented due to its importance as an early postglacial paleoclimatic indicator. The pollen of this species was distinguished from Picea mariana using the qualitative criteria developed by Hansen and Engstrom (1985).
Unless otherwise stated, all radiocarbon dates are uncalibrated, and standard deviations are 2σ. Calibrated dates were obtained using CALIB (Stuiver and Reimer, 1993). Within Unit A1, dates were also interpolated assuming a steady rate of sedimentation; this assumption is probably warranted within the organic deposit only, because it is lithologically uniform and does not exhibit unconformities.
Results
Zone I-a (34-29 cm) – Cyperaceae and aquatics (>10.6 ka BP)
Zone I-a lies in the gleyed silts and clays that underlie the organic deposit in Unit A1 (Figs. 3 and 4). This zone contains the highest diversity of macroremains from emergent and aquatic plant taxa; Cyperaceae is dominant (Fig. 5). In particular, the zone has high but declining values of Carex (55-10 seeds/150 ml), with a secondary component consisting of Menyanthes (0-30 seeds/150 ml), Myriophyllum (10-30 seeds/150 ml), Picea needles (10-25 seeds/150 ml), Hippuris vulgaris (0-15 seeds/150 ml), Scirpus (5-15 seeds/ 150 ml); low values of Eleocharis (0-10 seeds/150 ml), Potamogeton (0-10 seeds/150 ml), Zannichellia (0-2 seeds/ 150 ml), Potentilla palustris (0-2 seeds/150 ml), Equisetum (0-10 stem fragments/150 ml), Chara (10-0 oogonia/150 ml), mosses, and one Populus bud scale. Preservation in this zone was excellent because several Carex perigynia were intact and contained achenes. Assuming a steady rate of sedimentation, this zone ends ~10.6 ka BP.
Zone I-b (29-20 cm) – Emergents (~10.6-10.1 ka BP)
This zone is characterized by a decline in macroremains from Cyperaceae and aquatics, an increase in emergents such as Menyanthes trifoliata (50-200 seeds/150 ml), Potentilla palustris (0-20 seeds/150 ml), Scirpus cf. S. validus (10-20 seeds/150 ml) and Equisetum, and abundant remains of Catoscopium. Small quantities of the following were also observed: Picea, Eleocharis, Hippuris, Myriophyllum, Potamogeton, and others.
Zone I-c (20-14 cm) – Drepanocladus (10.1-9.8 ka BP)
This zone is dominated by the moss Drepanocladus (400-600 fragments/150 ml). Remains of aquatic plants, with the exception of Hippuris vulgaris (7 seeds/150 ml), are absent. Macroremains from emergent taxa such as Menyanthes, Equisetum, and Carex are present, although in relatively low numbers.
Zone II (14-0 cm) – Menyanthes-Equisetum (~9.8-9.1 ka BP)
Zone II is dominated by macroremains from Menyanthes trifoliata (50-420/150 ml) and Equisetum (50-450/10 ml). In the middle of this zone, M. trifoliata peaks while all other taxa decline (Fig. 4). In contrast to zones I-b and I-c, there were no mosses and only a few seeds of Hippuris and Myriophyllum. Zone II also displays an overall decline in Picea and, at the top of this zone, a decline in all taxa.
Figure 3
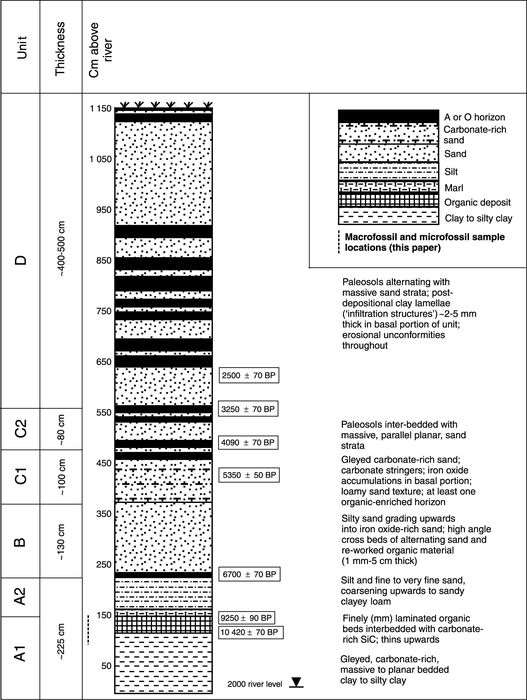
Composite schematic column of the Flintstone Hill cutbank, after Boyd et al. (2003). Paleoecological sample section in Unit A1 indicated by dashed line.
Stratigraphie de la coupe de la rive de Flintstone Hill (d'après Boyd et al., 2003). L'emplacement de l'échantillon paléoécologique prélevé dans la coupe de l’unité A1 est indiqué par un trait.
Interpretation
Zone I-a
Zone I-a (>10.6 ka BP) documents an herbaceous community dominated by Cyperaceae. Most taxa in Zone I-a indicate somewhat deeper water levels in contrast to subsequent zones (Jeglum, 1971). Deeper water levels in this zone are suggested by the abundance of aquatic taxa such as Myriophyllum, Hippuris, and Potamogeton. As well, Chara indicates that the water was carbonate-rich, warm, shallow and therefore not directly fed by meltwater (cf. Teller, 1989; Teller et al., 2000).
Carbonized Picea needles in Zone I- a indicate that spruce was part of the regional vegetation by at least this time. Furthermore, it was likely confined to the drier uplands surrounding the Hind basin, because virtually all spruce needles in the macrobotanical assemblage are carbonized, and were therefore carried into the basin by fire updrafts or water transport. Thus, although the local landscape is interpreted as treeless throughout the Pleistocene-Holocene transition, spruce was undoubtedly present on well-drained uplands within the study area (Boyd et al., 2003).
Figure 4
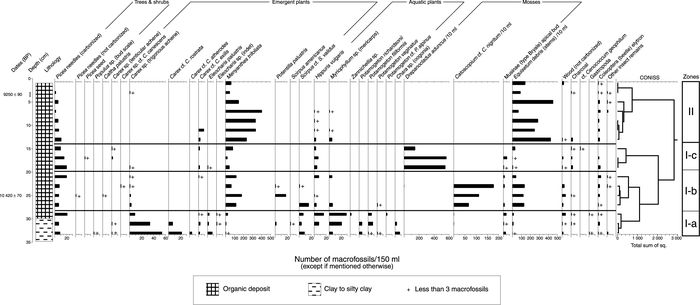
Plant macrofossil assemblage, Unit A1, Flintstone Hill site.
Assemblage de macrofossiles de plantes de l’unité A1, au site de Flintstone Hill.
Figure 5
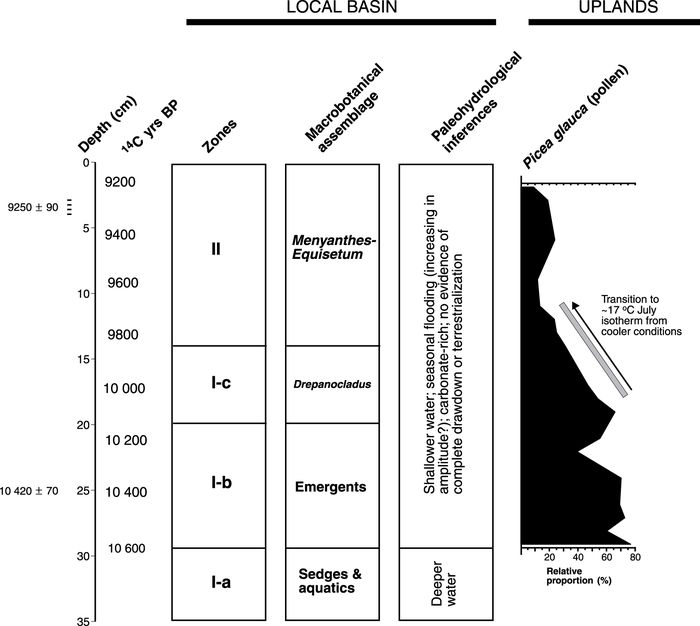
Summary of Unit A1 macrofossil zones with their paleohydrological implications. The Picea glauca (white spruce) pollen profile obtained from this deposit is in its exact stratigraphic position.
Sommaire des zones de macrofossiles de l’unité A1 et implications paléohydrologiques. Le profil pollinique de Picea glauca (épinette blanche) prélevé est dans sa position stratigraphique exacte.
Zone I-b
The decline in aquatics and increase in emergent plants in this zone indicate shallower water levels beginning between 10.6 and 10.1 ka BP. However, the absence of mesophytes, soil development, and deterioration of macrofossils suggest that water levels were always above surface in this zone. Although the lateral extension of a floating mat could also produce a greater representation of emergent plants, the absolute decline in aquatic plant remains supports the idea that an overall decline in water level occurred within the local basin. These lower water levels, furthermore, persisted until at least 9.1 ka BP (Zone II). Paleoecologic evidence of drainage is supported by the stratigraphic transition from massive silty clay to fine organic laminae in Unit A1 (Boyd et al., 2003) (Figs. 3 and 4).
Because the apparent decline in water depth in Zone I-b has no relationship to the onset of early postglacial warming (see Fig. 5), climate was probably not responsible for hydrologic change. Instead, this process may reflect a gradual decline in the base level of glacial Lake Hind. As argued in an earlier study (Boyd et al., 2003), much of the central glacial Lake Hind basin was drained shortly before ~10.4 ka BP due, at least in part, to the routing of meltwater from glacial Lake Regina (Sun, 1996; Sun and Teller, 1997). This event deepened and widened the Pembina spillway (Sun, 1996; Sun and Teller, 1997) and would have drained Lake Hind to a lower elevation.
Zone I-c
Zone I-c (~10.1-9.8 ka BP) largely consists of Drepanocladus aduncus (= D. polycarpus) remains. This sickle-branch moss thrives when submerged and is a good indicator of open, wet, alkaline and calcareous habitats (Crum, 1976; Crum and Anderson, 1981). Today, D. aduncus is widespread across the boreal forest in nutrient-rich, oxygen-poor water (Johnson et al., 1995: 328).
At the Andrews site in south-central Saskatchewan (Yansa, 1998; Yansa and Basinger, 1999), layers composed almost entirely of this moss species dated to ~10.2 ka BP, and disappeared from the local fossil record before 8.8 ka BP. The disappearance of D. aduncus from the paleoecological record on the Canadian Prairies may record the shift to warmer conditions in the early Holocene. At the FSH site, Drepanocladus remains were not found in sediment younger than ~9.8 ka BP (i.e., Zone II).
Zone II
Menyanthes trifoliata is a clonal aquatic macrophyte found in shallow bogs and river margins throughout the boreal ecosystem (Thompson et al., 1998). Because there are no obvious vegetative (e.g., leaf, stem, root) remains of this plant in the samples, the seeds probably floated in to the site; complete deterioration of these remains is unlikely because vegetative debris from other plants (e.g., Equisetum, Drepanocladus) is abundant. Absence of abrasion on the seed coats suggests that water transport was over a small distance and that it occurred under low-energy conditions. The dominance of Menyanthes suggests persistence, throughout Zone II (~9.8-9.1 ka BP), of large local stands.
The importance of Menyanthes trifoliata may reflect regular flooding in the basin. Haraguchi (1991) observes that pure stands of M. trifoliata are found on floating mats where annual water level fluctuations are greater than 8 cm. This species, furthermore, is more likely to dominate a wetland as the amplitude of the flooding-drawdown cycle increases (Haraguchi, 1991: 255-257). Under decreased fluctuations, the Menyanthes-dominated mat is invaded by other species (Haraguchi, 1991: 262). This relationship exists because M. trifoliata is very tolerant of flooding (Coult and Vallance, 1958; Haraguchi, 1991: 261) due, at least in part, to its ability to withstand intense reducing conditions (Armstrong and Boatman, 1967). It is presumably for this reason that Menyanthes is commonly found on river margins in the boreal forest (Thompson et al., 1998). Scirpus validus, and Eleocharis palustris populations, on the other hand, rapidly die out under repeated flooding (Harris and Marshall, 1963: 338). It is possible, therefore, that the dominance of Menyanthes – particularly in Zone II – records regular flooding of the local basin. This agrees with the stratigraphic evidence of low-energy flooding in Unit A1 (see above).
Because the mean water level did not increase before 9.1 ka BP, flooding was likely seasonal and was balanced by drawdown or other means of drainage. For this reason, flooding due to differential isostatic tilt (Johnston, 1946; Teller and Thorleifson, 1983; Yu and McAndrews, 1994; Yu et al., 1996) is ruled out during this time. Although the source of water is not directly apparent, the proximity of the site to the Souris River channel (Fig. 2) suggests that the local basin may have regularly received floodwater once this river system was established. Sun (1996: 175) proposes that the Souris River began incising in the south-central Hind basin following regression of the lake to the Phase 9 level. Stratigraphic evidence from FSH indicates that the regression of Lake Hind to this level occurred shortly before ~10.4 ka BP (Boyd et al., 2003), and that alluvium from the Souris River was deposited locally between at least 9.1 and 6.7 ka BP (i.e., Unit A2) (Boyd, 2000; Running et al., 2002). On this basis, it is likely that the paleoecologic and stratigraphic evidence of low-energy flooding by at least 9.8 ka BP may simply record seasonal fluctuations of the ancestral Souris River and periodic overflow and/or seepage into the local basin.
Response to early postglacial warming?
There is good evidence for increased atmospheric temperatures in Zone II (9.8-9.1 ka BP). In this zone, the few Picea macrofossils matches the regional decline in Picea pollen (e.g., Ritchie and Lichti-Federovich, 1968; Ritchie, 1969, 1976; Yansa, 1998; Yansa and Basinger, 1999; Grimm, 2001). Local deterioration of white spruce beginning ~10.1 ka BP is also recorded in the pollen profile from Unit A1 (Boyd et al., 2003) (Fig. 5). The decline in spruce across the southern Canadian Prairies before 10 ka BP represents a shift in the southern limit of white spruce due to atmospheric warming to at least the 17 or 18 °C July isotherm (Zoltai, 1975; Ritchie and Harrison, 1993). Under a warming climate, onset of aridity no longer made white spruce viable and eliminated it from the late-glacial landscape (Ritchie, 1976). During Zone II, the Laurentide Ice Sheet was in full retreat following a brief readvance in southern Manitoba ~10 ka BP (Teller et al., 1983; Smith and Fisher, 1993; Thorleifson, 1996).
On the northwestern Plains and Mackenzie Delta, pollen evidence for dry lake basins and drought-tolerant taxa is common between 10 to 8 ka BP (Ritchie et al., 1983; Barnosky, 1989; Schweger and Hickman, 1989). In contrast, Chenopodium salinum maxima in a macrofossil record from the Missouri Coteau of Saskatchewan (Yansa, 1998; Yansa and Basinger, 1999) suggest delayed onset of the Hypsithermal until sometime between 8.8 and 7.7 ka BP. However, mineralogical data from the same site (Aitken et al., 1999) do not support this conclusion, and instead indicate severe aridity beginning ~10 ka BP. Similarly, in southwestern Saskatchewan, mineral assemblages from both Clearwater (Last et al., 1998; Leavitt et al., 1999) and North Ingebrigt (Shang and Last, 1999) Lakes indicate saline to hypersaline conditions and low relative humidity between ~10 and 9 ka BP. As suggested by Aitken et al. (1999: 180), variation in the timing of maximum aridity from different proxies may reflect differing response times of hydrological, ecological, and geomorphic systems. Additionally, differences in the sensitivity of the proxies themselves may also account for these results. For example, Chenopodium salinum peaks in the fossil record on the Canadian Prairies are probably a better marker of maximum aridity (Aitken et al., 1999: 180) rather than the onset of aridity.
Given the local and regional evidence of abrupt postglacial warming and increased aridity ~10 ka BP, does the macrobotanical record from FSH provide any clear vegetative responses to this significant change in climate? In a previous study (Boyd et al., 2003), I hypothesized that higher-amplitude flooding-drawdown cycles reconstructed for Zone II (9.8-9.1 ka BP) were induced by climate change. However, if evapotranspiration rates increased in the Hind basin after ~10 ka BP, the absence of evidence for terrestrialization (e.g., soil development, presence of mesophytes, poor organic preservation) indicates that complete drawdown did not occur in this shallow wetland before 9.1 ka BP. This suggests that climate-induced drawdown was buffered for at least 1000 years after the first signs of local and regional climate change (Fig. 5). These results are consistent with plant macrofossil data from the Missouri Coteau of Saskatchewan (Yansa, 1998; Yansa and Basinger, 1999). In her explanation, Yansa (1998) argues that buried ice masses may have boosted moisture levels in the early Holocene through prolonged release of meltwater into aquifers. Certainly, the Turtle Mountain upland located on the southeastern side of the Hind basin held unusually thick accumulations of stagnant dirty ice following deglaciation (Deal, 1972; Sun, 1996: 157). The Riding and Duck Mountain uplands immediately to the north of glacial Lake Hind were also covered with large stagnant ice bodies (Klassen, 1979). This suggests that a regular meltwater supply may have existed in southwestern Manitoba long after deglaciation; this supply, furthermore, may have increased after 10 ka BP due to summer melting of buried ice. Based on the available evidence, however, it is not possible to directly evaluate the impact of regional changes in meltwater supply on the hydrology of the local basin.
Instead, a more direct explanation is that regular flooding of the Souris River may have been sufficient to mitigate the ecological effects of climate change in this shallow wetland before ~9.1 ka BP. In other words, regular input of floodwater may have helped maintain a permanently saturated environment at a time when many other basins on the northern Prairies were dry (Barnosky, 1989; Schweger and Hickman, 1989).
Conclusions
Based on changes in plant macrofossils through time in a late glacial–early Holocene (>10.6-9.1 ka BP) wetland deposit, I propose the following:
Aquatic macrophyte succession at the study site indicates a gradual decline in water depth between ~10.6 and 10.1 ka BP. This process may record a lowering of the base level of glacial Lake Hind due to drainage through its northeastern outlet(s), and was not directly due to climate change.
The importance of Menyanthes trifoliata in this sequence after ~10.6 ka BP, but particularly between ~9.8 and 9.1 ka BP, may reflect seasonal, low energy, flooding of the local basin. This interpretation is supported by stratigraphic evidence (e.g., clastic intebeds with ripples in the upper 15 cm of Unit A1; deposition of alluvium in Unit A2). Because a mean increase in water depth is not indicated, flooding due to differential isostatic tilt did not occur during this time frame. Instead, proximity of the FSH site to the Souris River channel suggests that the basin may have periodically received floodwater from this system.
Local decline of Picea glauca on uplands surrounding the Hind basin is recorded by both pollen and macrofossils, and indicates atmospheric warming up to at least the 17 or 18 °C July isotherm beginning ~10.1 ka BP. The timing of Picea decline at this site is consistent with other records on the Canadian Prairies.
Despite local and regional evidence for abrupt postglacial warming ~10 ka BP, there is no stratigraphic or paleoecologic evidence of complete drawdown in this shallow wetland before 9.1 ka BP. This suggests that drawdown was locally buffered by some means. A similar delay in the response of wetland vegetation to early Holocene climate change was also observed on the Missouri Coteau of Saskatchewan (Yansa, 1998; Yansa and Basinger, 1999). In this region, regular feeding of basins by stagnant ice may have produced this apparent delay (Yansa, 1998). Although uplands surrounding the Hind basin held thick accumulations of stagnant ice, a more direct explanation is that regular floodwater input from the ancestral Souris River was a sufficient buffer against climate-induced drawdown up to at least 9.1 ka BP. In general, this discussion highlights the potential importance of local hydrologic processes in producing complex, asynchronous, responses of wetland vegetation to abrupt climate change.
Appendices
Acknowledgments
This analysis was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC). I am grateful to this agency for funding, and to the Department of Geological Sciences at the University of Manitoba for the use of its facilities. Thanks are also due to A. Beaudoin, R. Baker, and J.H. McAndrews for their helpful critiques. Any errors in fact and/or interpretation are mine.
References
- Aitken, A.E., Last, W.M. and Burt, A.K., 1999. The lithostratigraphic record of late Pleistocene-Holocene environmental change at the Andrews site near Moose Jaw, Saskatchewan, p. 173-181. In D. Lemmen and R.E. Vance, eds., Holocene Climate and Environmental Change in the Palliser Triangle: A Geoscientific Context for Evaluating the Impacts of Climate Change on the Southern Canadian Prairies. Geological Survey of Canada, Ottawa, Bulletin 534, 295 p.
- Armstrong, D. and Boatman, D.J., 1967. Some field observations relating the growth of bog plants to conditions of soil aeration. Journal of Ecology, 55: 101-110.
- Barnosky, C.W., 1989. Postglacial vegetation and climate in the northwestern Great Plains of Montana. Quaternary Research, 31: 57-73.
- Barnosky, C.W., Grimm, E.C. and Wright, H.E., Jr., 1987. Towards a postglacial history of the northern Great Plains: A review of the paleoecologic problems. Annals of the Carnegie Museum, 56: 259-273.
- Beaudoin, A.B., 1993. A compendium and evaluation of postglacial pollen records in Alberta. Canadian Journal of Archaeology, 17: 92-112.
- Berggren, G., 1969. Atlas of Seeds and Small Fruits of Northwest-European Plant Species, Part 2: Cyperaceae. Swedish Natural Science Research Council, Stockholm, 68 p.
- Birks, H.H., 1973. Modern macrofossil assemblages in lake sediments in Minnesota, p. 173-189. In H.J.B Birks and R.G. West, eds., Quaternary Plant Ecology. Blackwell Scientific Publications, Oxford, 326 p.
- Boyd, M., 2000. Late Quaternary Geoarchaeology of the Lauder Sandhills, Southwestern Manitoba, Canada. Ph.D. thesis, University of Calgary, 300 p.
- _____ 2002. Identification of anthropogenic burning in the paleoecological record of the northern Prairies: A new approach. Annals of the Association of American Geographers, 92: 471-487.
- Boyd, M., Running IV, G.L. and Havholm, K., 2003. Paleoecology and geochronology of glacial Lake Hind during the Pleistocene-Holocene transition: A context for Folsom surface finds on the Canadian Prairies. Geoarchaeology: An International Journal, 18: 583-607.
- Clayton, L. and Moran, S.R., 1982. Chronology of Late Wisconsin glaciation in middle North America. Quaternary Science Reviews, 1: 55-82.
- Coult, D.A. and Vallance, K.B., 1958. Observations on the gaseous exchanges which take place between Menyanthes trifoliata L. and its environment, part II. Journal of Experimental Botany, 9: 384-402.
- Crum, H., 1976. Mosses of the Great Lakes Forest. Revised Edition. University Herbarium, University of Michigan, Ann Arbor, 471 p.
- Crum, H.A. and Anderson, L.E., 1981. Mosses of Eastern North America. Columbia University Press, New York, 2 vol., 1328 p.
- David, P.P., 1977. Sand Dune Occurrences of Canada: A Theme and Resource Inventory of Eolian Landforms in Canada. Department of Indian and Northern Affairs, National Parks Branch, Ottawa, Report 74-230, 183 p.
- Deal, D.E., 1972. Geology of Rolette County, North Dakota. North Dakota Geological Survey, Grand Forks, Bulletin 58, 89 p.
- Fenton, M.M., Moran, S.R., Teller, J.T. and Clayton, L., 1983. Quaternary stratigraphy and history in the southern part of the Lake Agassiz basin, p. 40-74. In J.T. Teller and L. Clayton, eds., Glacial Lake Agassiz. Geological Association of Canada, St. John’s, Special Paper 26, 451 p.
- Grimm, E.C., 1987. CONISS: A FORTRAN-77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Computers and Geosciences, 13: 13-35.
- _____ 2001. Trends and palaeoecological problems in the vegetation and climate history of the northern Great Plains, USA. Biology and Environment: Proceedings of the Royal Irish Academy of Sciences, 101B: 47-64.
- Hansen, B.C.S. and Engstrom, D.R., 1985. A comparison of numerical and qualitative methods of separating pollen of black and white spruce. Canadian Journal of Botany, 63: 2159-2163.
- Haraguchi, A., 1991. Effect of a flooding-drawdown cycle on vegetation in a system of floating peat mat and pond. Ecological Research, 6: 247-263.
- Harris, S.W. and Marshall, W.H., 1963. Ecology of water-level manipulations on a northern marsh. Ecology, 44: 331-343.
- Jeglum, J.K., 1971. Plant indicators of pH and water level in peatlands at Candle Lake, Saskatchewan. Canadian Journal of Botany, 49: 1661-1676.
- Johnson, D., Kershaw, L., MacKinnon, A. and Pojar, J. 1995. Plants of the Western Boreal Forest and Aspen Parkland. Lone Pine Publishing, Edmonton, 392 p.
- Johnston, W.A., 1946. Glacial Lake Agassiz With Special Reference to the Mode of Deformation of the Beaches. Geological Survey of Canada Memoir 128, 20 p.
- Kehew, A.E. and Clayton, L., 1983. Late Wisconsin floods and the development of the Souris-Pembina spillway system in Saskatchewan, North Dakota, and Manitoba, p. 187-209. In J.T. Teller and L. Clayton, eds., Glacial Lake Agassiz. Geological Association of Canada, St. John’s, Special Paper 26, 451 p.
- Kehew, A.E. and Lord, M.L., 1986. Origin and large-scale erosional features of glacial-lake spillways in the northern Great Plains. Bulletin of the Geological Society of America, 97: 162-177.
- Kehew, A.E. and Teller, J.T., 1994. History of the Late Glacial runoff along the southwestern margin of the Laurentide Ice Sheet. Quaternary Science Reviews, 13: 859-877.
- Klassen, R.W., 1972. Wisconsin events and the Assiniboine and Qu'Appelle Valleys of Manitoba and Saskatchewan. Canadian Journal of Earth Sciences, 9: 544-560.
- _____ 1979. Pleistocene Geology and Geomorphology of the Riding Mountain and Duck Mountain Areas, Manitoba-Saskatchewan. Geological Survey of Canada, Ottawa, Memoir 396, 52 p.
- _____ 1983. Assiniboine Delta and the Assiniboine-Qu'Appelle Valley system – implication concerning the history of Lake Agassiz in southwestern Manitoba, p. 211-220. In J.T. Teller and L. Clayton, eds., Glacial Lake Agassiz. Geological Association of Canada, St. John’s, Special Paper 26, 451 p.
- Last, W.M., Vance, R.E., Wilson, S. and Smol, J.P., 1998. A multi-proxy record of early Holocene hydrologic change on the northern Great Plains of southwestern Saskatchewan, Canada. The Holocene, 8: 503-520.
- Leavitt, P.R., Vinebrook, R.D., Hall, R.I., Wilson, S.E, Smol, J.P., Vance, R.E. and Last, W.M., 1999. Multiproxy record of prairie lake response to climate change and human activity, Clearwater Lake, Saskatchewan, p. 125-138. In D.S. Lemmen and R.E. Vance, eds., Holocene Climate and Environmental Change in the Palliser Triangle: A Geoscientific Context for Evaluating the Impacts of Climate Change on the Southern Canadian Prairies. Geological Survey of Canada, Ottawa, Bulletin 534, 295 p.
- Lévesque, P.E.M., Dinel, H. and Larouche, A., 1988. Guide to the Identification of Plant Macrofossils in Canadian Peatlands. Agriculture Canada, Research Branch, Ottawa, Publication 1817, 65 p.
- Martin, A.C. and Barkley, W.D., 2000. Seed Identification Manual. Reprint. Blackburn, Caldwell, 221 p.
- Montgomery, F.H., 1977. Seeds and Fruits of Plants of Eastern Canada and Northeastern United States. University of Toronto Press, 232 p.
- Ritchie, J.C., 1969. Absolute pollen frequencies and carbon-14 age of a section of Holocene lake sediment from the Riding Mountain area of Manitoba. Canadian Journal of Botany, 47: 1345-1349.
- _____ 1976. The Late-Quaternary vegetational history of the western interior of Canada. Canadian Journal of Botany, 54: 1793-1818.
- Ritchie, J.C., Cwynar, L.C. and Spear, R.W., 1983. Evidence from north-west Canada for an early Holocene Milankovitch thermal maximum. Nature, 305: 126-128.
- Ritchie, J.C. and Harrison, S.P., 1993. Vegetation, lake levels, and climate in western Canada during the Holocene, p. 401-414. In H.E. Wright Jr., J.E. Kutzbach, T. Webb III, W.F. Ruddiman, F.E. Street-Perrott and P.J. Bartlein, eds., Global Climates Since the Last Glacial Maximum. University of Minnesota Press, Minneapolis, 569 p.
- Ritchie, J.C. and Lichti-Federovich, S., 1968. Holocene pollen assemblages from the Tiger Hills, Manitoba. Canadian Journal of Earth Sciences, 5: 873-880.
- Running IV, G.L., Havholm, K., Boyd, M. and Wiseman, D., 2002. Holocene stratigraphy and geomorphology of Flintstone Hill, Lauder Sandhills, glacial Lake Hind basin, southwestern Manitoba, Canada. Géographie physique et Quaternaire, 56: 291-303.
- Schweger, C.E. and Hickman, M., 1989. Holocene paleohydrology of central Alberta: Testing the general-circulation-model climate simulations. Canadian Journal of Earth Sciences, 26: 1826-1833.
- Shang, Y. and Last, W.M., 1999. Mineralogy, lithostratigraphy, and inferred geochemical history of North Ingebrigt Lake, Saskatchewan, p. 99-110. In D.S. Lemmen and R.E. Vance, eds., Holocene Climate and Environmental Change in the Palliser Triangle: A Geoscientific Context for Evaluating the Impacts of Climate Change on the Southern Canadian Prairies. Geological Survey of Canada, Ottawa, Bulletin 534, 295 p.
- Smith, D.G. and Fisher, T.G., 1993. Glacial Lake Agassiz: The Northwestern outlet and paleoflood. Geology, 21: 9-12.
- Stuiver, M. and Reimer, P.J., 1993. Extended 14C data base and revised Calib 3.0 14C age calibration program. Radiocarbon, 35: 215-230.
- Sun, C., 1996. Sedimentology and Geomorphology of the Glacial Lake Hind Area, Southwestern Manitoba, Canada. Ph.D. thesis, University of Manitoba, Winnipeg, 189 p.
- Sun, C. and Teller, J.T., 1997. Reconstruction of glacial Lake Hind in southwestern Manitoba, Canada. Journal of Paleolimnology, 17: 9-21.
- Teller, J.T., 1989. Importance of the Rossendale site in establishing a deglacial chronology along the southwestern margin of the Laurentide Ice Sheet. Quaternary Research, 32: 12-23.
- Teller, J.T., Risberg, J., Matile, G. and Zoltai, S., 2000. Postglacial history and paleoecology of Wampum, Manitoba, a former lagoon in the Lake Agassiz basin. Bulletin of the Geological Society of America, 112: 943-958.
- Teller, J.T. and Thorleifson, L.H., 1983. The Lake Agassiz-Lake Superior connection, p. 261-290. In J.T. Teller and L. Clayton, eds., Glacial Lake Agassiz. Geological Association of Canada, St. John’s, Special Paper 26, 451 p.
- Teller, J.T., Thorleifson, L.H., Dredge, L.A., Hobbs, H.C. and Schreiner, B.T., 1983. Maximum extent and major features of Lake Agassiz, p. 43-45. In J.T. Teller and L. Clayton, eds., Glacial Lake Agassiz. Geological Association of Canada, St. John’s, Special Paper 26, 451 p.
- Thompson, F.L., Hermanutz, L.A. and Innes, D.J., 1998. The reproductive ecology of distylous Menyanthes trifoliata (Menyanthaceae). Canadian Journal of Botany, 76: 818-828.
- Thorleifson, L.H., 1996. Review of Lake Agassiz history, p. 55-84. In J.T. Teller, L.H. Thorleifson, G. Matile, and W.C. Brisbin, eds., Sedimentology, Geomorphology, and History of the Central Lake Agassiz Basin. Geological Association of Canada/Mineralogical Association of Canada, Joint Annual Meeting (Winnipeg, Manitoba), Fieldtrip Guidebook, 101 p.
- Vance, R.E., Clague, J.J. and Mathewes, R.W., 1993. Holocene paleohydrology of a hypersaline lake in southeastern Alberta. Journal of Paleolimnology, 8: 103-120.
- Wallace, W.G., 2002. Eolian sand dunes of the glacial Lake Hind basin, Manitoba, Canada. M.Sc. thesis, University of Wisconsin-Madison, 69 p.
- Warner, B.G., 1990. Plant macrofossils. In B.G. Warner, ed., Methods in Quaternary Ecology. Geoscience Canada, St. John’s, Reprint Series 5, 170 p.
- Wiken, E.B., 1986. Terrestrial Ecozones of Canada. Environment Canada, Lands Directorate, Ottawa, Ecological Land Classification Series 19, 26 p.
- Yansa, C.H., 1998. Holocene paleovegetation and paleohydrology of a prairie pothole in southern Saskatchewan, Canada. Journal of Paleolimnology, 19: 429-441.
- Yansa, C.H. and Basinger, J.F., 1999. A postglacial plant macrofossil record of vegetation and climate change in southern Saskatchewan, p. 139-154. In D. Lemmen and R.E. Vance, eds., Holocene Climate and Environmental Change in the Palliser Triangle: A Geoscientific Context for Evaluating the Impacts of Climate Change on the Southern Canadian Prairies. Geological Survey of Canada, Ottawa, Bulletin 534, 295 p.
- Yu, Z. and McAndrews, J.H., 1994. Holocene water levels at Rice Lake, Ontario, Canada: Sediment, pollen and plant-macrofossil evidence. The Holocene, 4: 141-152.
- Yu, Z., McAndrews, J.H. and Siddiqi, D., 1996. Influences of Holocene climate and water levels on vegetation dynamics of a lakeside wetland. Canadian Journal of Botany, 74: 1602-1615.
- Yu, Z. and Wright, H.E., Jr., 2001. Response of interior North America to abrupt climate oscillations in the North Atlantic region during the last deglaciation. Earth-Science Reviews, 52: 333-369.
- Zoltai, S.C., 1975. Southern Limit of Coniferous Trees on the Canadian Prairies. Environment Canada Forestry Service, Northern Forest Research Centre, Edmonton, Information Report NOR-X-128, 12 p.
 10.7202/009112ar
10.7202/009112arList of figures
Figure 1
Map of southern Manitoba with the location of glacial Lake Hind, the Flintstone Hill (FSH) study site, southwestern Manitoba, and major physiographic features. Canadian Prairies Ecozone (Wiken, 1986) is shaded in grey.
Carte du sud du Manitoba et localisation du Lac glaciaire Hind, du site d’étude Flintstone Hill (FSH), dans le sud-ouest du Manitoba, et des principaux éléments physiographiques. L’écozone des Prairies (Wiken, 1986) est en gris.
Figure 2
Glacial Lake Hind with location of Flintstone Hill (FSH) study site. Phase 4 to Phase 9 lake shorelines after Sun (1996).
Lac glaciaire Hind et localisation du site d’étude Flintstone Hill (FSH). Phases 4 à 9 du littoral d'après Sun (1996).
Figure 3
Composite schematic column of the Flintstone Hill cutbank, after Boyd et al. (2003). Paleoecological sample section in Unit A1 indicated by dashed line.
Stratigraphie de la coupe de la rive de Flintstone Hill (d'après Boyd et al., 2003). L'emplacement de l'échantillon paléoécologique prélevé dans la coupe de l’unité A1 est indiqué par un trait.
Figure 4
Plant macrofossil assemblage, Unit A1, Flintstone Hill site.
Assemblage de macrofossiles de plantes de l’unité A1, au site de Flintstone Hill.
Figure 5
Summary of Unit A1 macrofossil zones with their paleohydrological implications. The Picea glauca (white spruce) pollen profile obtained from this deposit is in its exact stratigraphic position.
Sommaire des zones de macrofossiles de l’unité A1 et implications paléohydrologiques. Le profil pollinique de Picea glauca (épinette blanche) prélevé est dans sa position stratigraphique exacte.