Abstracts
Résumé
L’analyse pollinique et macrofossile des sédiments de deux lacs a livré l'histoire postglaciaire de la végétation au centre (lac du Triangle [48° 42’ 36’’ N, 65° 24’ 50’’ O]) et à l’est (Petit lac Bouchard [48° 51’ 14’’ N, 64° 35’ 52’’ O]) de la Gaspésie. Une toundra éparse à bouleau glanduleux s'est installée dès 11 200 BP (années étalonnées). L’afforestation s’effectue par l’entremise de l’épinette noire accompagnée ou non de peuplier faux-tremble vers 10 700 BP. La multiplication des populations d’épinette noire s’est ralentie ou a cessé vers 10 000 BP au profit de l'aulne crispé et du tremble, pendant une période où les feux étaient fréquents. La forêt fermée a mis un millénaire pour se constituer au centre de la Gaspésie mais dans l’est, il lui a fallu plus de 2000 ans. Cette longue période d’afforestation est attribuée à un climat maritime froid. Durant la phase forestière, les taxons régionalement thermophiles comme le pin blanc et le bouleau jaune ne se sont établis que dans la partie orientale, le centre de la Gaspésie restant dominé par des essences boréales : le sapin baumier, l'épinette noire et, dans une moindre mesure, le bouleau blanc. Le pin gris, aujourd’hui absent de la Gaspésie, était présent dans la région du lac du Triangle dès l’afforestation et jusque vers 3500 BP. Depuis 3500 ans, l'épinette noire et le sapin baumier se multiplient aux dépens des bouleaux dans les deux régions, traduisant ainsi l'instauration d'un climat plus humide et plus frais. Toutefois, le contraste avec la période antérieure est plus fort à l'est qu'au centre de la Gaspésie. Une première reconstitution des variations paléohydriques fondée sur les changements du niveau d’eau des lacs est proposée.
Abstract
Pollen and macrofossil analyses were used to reconstruct the postglacial vegetation history in the central (Lac du Triangle [48° 42’ 36’’ N, 65° 24’ 50’’ W]) and the eastern (Petit Lac Bouchard [48° 51’ 14’’ N, 64° 35’ 52’’ W]) part of Gaspésie. A sparse dwarf birch tundra existed as early as 11 200 BP (calendar years). Afforestation occurred around 10 700 BP involving black spruce sometimes accompanied or not by aspen. Growth of black spruce populations slowed down or was even reversed around 10 000 BP, and green alder and aspen experienced increased abundance during a period of elevated fire incidence. It took 1000 years for closed forests to be installed in central Gaspésie but in the eastern region, the process lasted 2000 years. Such a delayed afforestation could be due to a cold maritime climate. During the forest phase, regional thermophilous taxa like white pine and yellow birch occupied the eastern area whereas central Gaspésie was dominated by boreal tree species: balsam fir, black spruce and, to a lesser extent, white birch. Jack pine, presently absent from Gaspésie, was present around Lac du Triangle between afforestation to about 3500 BP. In response to a cooler and wetter climate during the last 3500 years, black spruce and balsam fir became more abundant, while white birch decreased in the two regions, more so around the eastern site. A first paleohydrological reconstruction is proposed, based on lake level changes.
Resumen
El análisis polínico y macrofósil de los sedimentos de dos lagos permitió establecer la historia postglaciar de la vegetación en la parte central y al este de la Gaspésie : Lac du Triangle (48o 42’ 36” N, 65o 24’ 50” O) y Petit lac Bouchard (48o 51’ 14” N, 64o 35’ 52” O) respectivamente. Hace unos 11 200 años se instaló una tundra recubierta de abedul glanduloso (Betula glandulosa). Más tarde, hace unos 10 700 años, la forestación se efectuó gracias al establecimiento de pícea negra (Picea mariana) y del álamo temblón americano (Populus tremuloides). Posteriormente, hace unos 10 000 años el aumento de la población de la pícea negra disminuyó o se detuvo dando paso a la colonización de alisos verdes (Alnus crispa) o álamos. Durante ese mismo periodo se registraron varios incendios. En la parte central de la Gaspésie el bosque tardó un millón de años en establecerse pero hacia el este solo requirió unos 2000 años. Este periodo tan largo de repoblación forestal es atribuido a un clima marítimo frío. Durante la fase forestal, los taxa de tipo termófilo tales como el pino blanco o el abedul amarillo no lograron establecerse mas que en la parte oriental, mientras que en la parte central de la Gaspésie dominaron las especies boreales del tipo del abeto balsámico, la pícea negra y en menor grado el abedul blanco. El pino gris, ausente actualmente en Gaspésie se encontraba presente en la región del Lac du Triangle desde el inicio de la forestación y hasta el año 3500 anterior al presente. Hace unos 3500 la pícea negra y el abeto balsámico se multiplicaron resplazando asi el abedul en las dos regiones, lo cual trajo como consecuencia el establecimiento de un clima más húmedo y más fresco. Sin embargo, el contraste con el periodo anterior es más fuerte en la parte este que en el centro de la Gaspésie. El manuscrito propone así mismo una primera reconstrucción de las variaciones paleohídricas basada en los cambios del nivel de agua de los lagos.
Article body
Introduction
La phytogéographie postglaciaire des marges septentrionales et méridionales de la péninsule gaspésienne est maintenant établie dans ses grandes lignes (Livingstone, 1968 ; Labelle et Richard, 1984 ; Jetté et Richard, 1992 ; Marcoux et Richard, 1995). Hormis l’étage sub-alpin au mont Albert (Richard et Labelle, 1989), l’intérieur des terres et l’est de la Gaspésie restent à ce titre « terres inconnues » (fig. 1).
Les études montrent des différences dans l’âge, dans la nature et dans la durée des diverses étapes de la colonisation végétale. La recrudescence, durant l’afforestation, d’un arbuste buissonnant, l’aulne crispé (Alnus viridis ssp. crispa [Ait.] Pursh.), tant dans les régions côtières qu’en milieu alpin, traduirait une halte ou même un recul dans la conquête des terres par les arbres. Le phénomène touche aussi le Bas-Saint-Laurent (Richard et al., 1992 ; Richard et Larouche, 1994) et la région de Charlevoix (Richard et Poulin, 1976 ; Labelle et Richard, 1981), mais son expression au coeur et à l’est de la Gaspésie n’est pas connue.
La déglaciation de la Gaspésie (Richard et al., 1997), étendue entre 13 000 et 9000 BP (années étalonnées), explique sans doute une part de la diversité initiale enregistrée, tant dans la durée que dans la nature même des formations végétales. Toutefois, les contrastes physiographiques que présente la péninsule par ses plateaux étagés, ses montagnes, ses côtes, devraient avoir induit des différences régionales durant tout le postglaciaire. C’est ce que nous voulons évaluer. Nous émettons donc l’hypothèse que le centre de la péninsule a toujours hébergé une végétation d’affinité plus boréale qu’en périphérie. Afin de la vérifier, nous avons effectué l’analyse pollinique et l’analyse macrofossile des sédiments du lac du Triangle et du Petit lac Bouchard. En incorporant des indications sur les changements du niveau lacustre et sur l’incidence générale des feux, nos résultats précisent l’histoire de la végétation et du climat postglaciaires à l’échelle gaspésienne.
Description des sites
Les caractéristiques des deux lacs étudiés et de leurs sédiments sont présentées au tableau I. Le lac du Triangle se trouve au centre de la péninsule gaspésienne, à la tête de la rivière Saint-Jean dans le canton de Gastonguay, à environ 30 km au sud de Murdochville (fig. 2a). Le Petit lac Bouchard est situé à l’est de la péninsule, à la marge du plateau gaspésien et à proximité du bas plateau côtier. Il se trouve au nord de la rivière Dartmouth, à environ 3,5 km du village de Cortéréal (fig. 2b). Il s’agit de deux petits lacs de tête (≤2 ha), de faible profondeur (≤2,5 m d’eau), qui occupent de petits bassins versants (<0,25 km2) dans des paysages de plateaux de collines ou montueux (fig. 2). Les deux cuvettes sont peu encaissées dans des dépôts meubles (sédiments alluviaux ou till) composés en partie de roches carbonatées provenant de formations géologiques calcareuses (ministère de l’Énergie et des Ressources du Québec, 1987 ; Veillette et Cloutier, 1993). Les conditions climatiques dans la région du lac du Triangle sont plus froides et plus humides et le nombre de jours sans gel moins élevé que dans la région de Gaspé, à l’est de la péninsule (Proulx et al., 1987 ; Environnement Canada, 2003) (tabl. I).
Figure 1

Sites des reconstitutions paléophytogéographiques réalisées en Gaspésie et localisation des sites de l’étude : (1) lac Turcotte et (2) lac à Léonard (Labelle et Richard, 1984), (3) lac du Diable (Richard et Labelle, 1989), (4) lac Harriman, (5) lac Caribou, (6) lac à Robin et (7) lac à Raymond (Jetté et Richard, 1992), (8) lac à Euloge et lac J’Arrive (9) (Marcoux et Richard, 1995), (10) lac du Triangle et (11) Petit lac Bouchard (cette étude).
Map of Gaspésie showing location of studied sites for paleophytogeographical reconstruction of this paper and other studies: (1) Lac Turcotte and (2) Lac à Léonard (Labelle et Richard, 1984), (3) Lac du Diable (Richard et Labelle, 1989), (4) Lac Harriman, (5) Lac Caribou, (6) Lac à Robin et (7) Lac à Raymond (Jetté et Richard, 1992), (8) Lac à Euloge et Lac J’Arrive (9) (Marcoux et Richard, 1995), (10) Lac du Triangle et (11) Petit Lac Bouchard (this paper).
Tableau I
Données sur les lacs étudiés et leurs sédiments

Au lac du Triangle, les stations mésiques sont dominées par le sapin baumier (Abies balsamea) et l’épinette noire (Picea mariana). Sur les monts environnants évoluent des peuplements denses de peupliers faux-trembles (Populus tremuloides) et de bouleaux blancs (Betula papyrifera) issus des coupes forestières et d’un feu de forêt survenu vers 1960. Aux abords ouest et sud-ouest du lac, une arbustaie à myrique baumier (Myrica gale) et à aulne rugueux (Alnus incana) accompagnée d’éricacées, de mélèzes (Larix laricina) et d’épinettes noires (Picea mariana) occupe une tourbière riveraine tapissée de sphaignes (Sphagnum capillifolium). Plus loin, une pessière noire dense ceinture le lac.
Aux environs du Petit lac Bouchard, les forêts matures sont rares et de grands espaces ont subi des coupes forestières récentes. Là où la forêt est présente, le sapin baumier, le pin blanc (Pinus strobus), l’épinette blanche (Picea glauca), le bouleau blanc, le bouleau jaune (Betula alleghaniensis), le peuplier faux-tremble et le peuplier baumier (Populus balsamifera) constituent des peuplements mixtes dans les stations les mieux drainées. Ils sont accompagnés de l’érable rouge (Acer rubrum), du chêne rouge (Quercus rubra var. borealis), de l’orme (Ulmus americana), du frêne de Pennsylvanie (Fraxinus pennsylvanica) et du thuya (Thuja occidentalis). Les stations les plus humides, en périphérie du lac, sont dominées par l’épinette noire et le mélèze, sauf sur les rives ouest et nord-ouest où le myrique baumier, l’aulne rugueux et les Éricacées accompagnés d’épinettes noires, de mélèzes et de bouleaux blancs constituent une arbustaie arborée dense surmontant un sol organique couvert de sphaignes (Sphagnum capillifolium et Sphagnum squarrosum).
Méthodes
Terrain
Les sédiments du lac du Triangle (TRIANGLE) et la partie basale de ceux du Petit lac Bouchard (BOUCHARDB) ont été prélevés en mars 1994 depuis le couvert de glace. Le reste du profil du Petit lac Bouchard (BOUCH96) a été obtenu en juin 1996. Les carottes proviennent du centre des bassins et furent extraites à l’aide d’une sonde à piston de type Livingstone (1955) modifiée. Au Petit lac Bouchard, un carottier de type Kajak-Brinkhurst (KB) modifié (Glew, 1988, 1989) fut utilisé pour récupérer intacte l’interface eau-sédiment (BOUCHS). La corrélation des carottes BOUCHARDB, BOUCH96 et BOUCHS par les profondeurs, la perte au feu et le contenu pollinique a permis de livrer un profil sédimentaire complet du Petit lac Bouchard (BOUCHARD).
Figure 2
a

Le lac du Triangle, à la tête de la rivière Saint-Jean.
Lac du Triangle, at the head of Rivière Saint-Jean.
b

Le Petit lac Bouchard, à la tête de la rivière Dartmouth.
Petit lac Bouchard, at the head of Rivière Dartmouth.
Topographie des sites d’étude
Maps that show the topography of studied sites
Laboratoire
Les sédiments furent conservés à une température de 5 °C. Les carottes furent décrites (Troëls-Smith, 1955 ; Aaby et Berglund, 1986) et des échantillons de 1 cm3, prélevés à tous les centimètres, furent soumis à la perte au feu (Dean, 1974) afin de déterminer leur contenu en matière organique (combustion à 600 °C) et en carbonates de calcium (CaCO3 ; combustion à 1 000 °C) (fig. 3a et b).
Les analyses sporopolliniques ont porté sur des volumes de 1 cm3. Les sédiments ont été soumis au traitement chimique standard d’extraction du pollen (Faegri et al., 1989). La concentration pollinique (n grains/cm3), fut établie par la méthode de Benninghoff (1962). L’identification et le dénombrement des grains de pollen, des spores et autres microfossiles (Pediastrum sp.) furent réalisés au microscope photonique à un grossissement de 400X (1000X pour les cas problématiques). La somme pollinique minimale est de 500 grains, sauf pour les niveaux inorganiques à la base des profils, là où la concentration sporopollinique est faible. Cette somme comprend le pollen des taxons arboréens, arbustifs et herbacés terricoles. La représentation des plantes aquatiques, des ptéridophytes et des algues fut exprimée par rapport à cette somme, dans la catégorie hors-total. L’identification s’est appuyée sur les clés de Richard (1970), de McAndrews et al. (1973), de Faegri et al. (1989) et de Moore et al. (1991), et sur la collection de référence du Laboratoire de paléophytogéographie et de palynologie de l’Université de Montréal. Les taxons arboréens Picea, Pinus, Abies et Populus ont été identifiés à l’espèce grâce à la clé de Richard (1970). Pour préciser l’identification du genre Betula, le diamètre équatorial de 100 grains de pollen a été mesuré à chaque niveau analysé. Le profil sédimentaire du lac du Triangle a été analysé à tous les 10 cm, alors que les sédiments du Petit lac Bouchard ont été analysés à tous les 2,5, 5 et 10 cm. Conjointement au pollen, la surface des charbons microscopiques a été mesurée afin d’évaluer l’incidence des feux durant la période d’accumulation sédimentaire. Les résultats sont exprimés en influx de charbons (mm2/cm2/année) (fig. 4).
Les macrorestes ont été extraits des sédiments par tamisage sous un doux jet d’eau et séparés dans quatre tamis (2 mm, 1 mm, 500 μm et 250 μm ; selon Larouche, 1979). Certains sédiments ont d’abord été chauffés quelques minutes dans une solution défloculante d’hydroxyde de potassium (KOH 10 %). L’identification a été réalisée à l’aide d’une loupe binoculaire à des grossissements de 10X et de 40X. Une collection de référence de pièces botaniques et les travaux de Lévesque et al. (1988) et de Young et Young (1992) furent consultés pour la détermination taxonomique. Au lac du Triangle, les analyses ont été effectuées sur des tranches de 1 cm à tous les 20 cm et resserrées aux portions supérieure (de 0 à 10 cm) et inférieure (de 350 à 387 cm) du profil ; pour le Petit lac Bouchard, elles ont porté sur des tranches de 3 cm à tous les 10 cm. Le volume analysé a varié de 10 à 100 cm3 ; les résultats furent standardisés pour un volume de 100 cm3.
Figure 3

Nature des sédiments (selon Aaby et Berglund, 1986), courbes de la perte au feu à 600 et 1 000 °C et des résidus et zones polliniques du lac du Triangle (a) et du Petit lac Bouchard (b).
Sediment stratigraphy (according to Aaby and Berglund, 1986), loss on ignition curves at 600 and 1 000 °C with regional pollen zone of Lac du Triangle (a) and Petit Lac Bouchard (b).
Des échantillons de macrorestes terricoles trouvés dans les sédiments du lac du Triangle et une partie de ceux du Petit lac Bouchard ont été soumis à la datation au 14C par spectrométrie de masse à l’aide d’un accélérateur de particules (SMA) (Litherland et Beukens, 1995). Des sédiments en vrac de la portion supérieure de BOUCHARD ont été soumis à la datation conventionnelle sur de la matière organique totale en raison de leur pauvreté en macrorestes terricoles. Les âges obtenus ont été étalonnés en années réelles, sidérales, à l’aide du logiciel CALIB 3.0 (Stuiver et Reimer, 1993).
Résultats
Nature des sédiments
Les sédiments du lac du Triangle couvrent 387 cm et présentent trois unités distinctes (fig. 3a). La première, entre 387 à 373 cm, correspond à l’argile silto-sableuse grise à la base du profil (PAF 600 °C : <15 %). La seconde, entre 373 et 125 cm, est composée de gyttja marneuse présentant de fines laminations (PAF 600 °C : 20 à 40 % ; PAF 1 000 °C : 10 à 30 %) et des lits de marne (373-366 cm et 200-212 cm ; PAF 1 000 °C : 30 à 35 %) qui alternent avec des couches de gyttja riche en matière organique (330-331 cm ; 310-315 cm ; 293-300 cm ; 250-270 cm et 226-240 cm ; PAF 600 °C : 30 à 55 %). Le fin litage de cette unité traduit la rareté, voire l’absence de perturbations biologique ou physique des sédiments de cette portion du profil. La troisième unité, allant de 125 à 0 cm, est une gyttja nettement plus riche en matière organique (PAF 600 °C : 65 et 85 % ; fig. 3a) et dépourvue de carbonates (PAF 1 000 °C moindre que 10 ou même 5 % ; Dean, 1974).
Figure 4
a

b

Taux d’accumulation des charbons microscopiques (mm2/cm2/an) et zones d’assemblage pollinique (ZAP) : (a) lac du Triangle et (b) Petit lac Bouchard. Chronologie en années étalonnées BP (années sidérales).
Microscopic charcoal accumulation rates (mm2/cm2/an) and regional pollen zones assemblages: (a) Lac du Triangle and (b) Petit Lac Bouchard. Chronology is in calibrated years BP (sideral years).
Le profil sédimentaire du Petit lac Bouchard, d’une longueur totale de 335 cm, présente deux unités : 1) des dépôts inorganiques : argile grise, silt, sable fin et sable grossier (330 à 324 cm) (PAF 600 °C < 5 %), et 2) de la gyttja indifférenciée (324 à 0 cm) (PAF 600 °C de 30 à 45 %) (fig. 3b). La perte au feu à 1 000 °C, se situant autour de 5 %, traduit l’absence de carbonates dans le profil.
Les influx de charbons de bois microscopiques dans les sédiments des deux lacs sont illustrés à la figure 4. Ils révèlent essentiellement une diminution des feux durant l’Holocène moyen (entre 7000 et 3000 BP environ ; Carcaillet et Richard, 2000 ; Asnong, 2001).
Données chronologiques
Un modèle de l’âge des sédiments selon la profondeur fut établi pour chacun des sites (fig. 5a et 5b) à partir des datations au 14C (tabl. II), un âge de 0 BP étant attribué à la surface des sédiments.
Deux des huit dates obtenues pour le lac du Triangle (2070 BP à 45,5-47 cm et 2095 BP à 40-53 cm), bien qu’elles se corroborent l’une et l’autre, ont été rejetées car jugées trop vieilles pour la profondeur dont elles proviennent (fig. 5a ; tabl. II). Ces âges supposent des variations du taux d’accumulation sédimentaire (TAS) qui ne sont pas appuyées par les concentrations polliniques ou le type de sédiment. Ils sont attribués au redépôt de vieux macrorestes terricoles à la suite d’une inondation des berges causée par l’élévation du niveau d’eau (voir plus loin). La date de 5580 BP (195-200 cm) est corroborée par la chute des pourcentages de Tsuga canadensis au niveau 190 cm (fig. 5a), chute présumée synchrone dans le nord-est de l’Amérique du Nord vers 5400 ± 300 étal. BP (4750 ± 50 BP ; Davis, 1981 ; Fuller, 1998 ; Bennett et Fuller, 2002). En conséquence, la date de 5720 BP fut rejetée parce que trop jeune. Malgré la nature variée des sédiments, le TAS, calculé par interpolation linéaire des dates retenues est relativement régulier et faible pour l’ensemble de l’Holocène (moyenne de 0,035 cm/an).
Figure 5
a

b

Courbes de l’âge en fonction de la profondeur des sédiments du lac du Triangle (a) et du Petit lac Bouchard (b). Les dates sont en années étalonnées BP. Les carrés noirs correspondent aux dates au radiocarbone par spectrométrie de masse à l’accélérateur de particules (SMA) de TRIANGLE ou de BOUCH96, les carrés blancs, aux dates SMA de BOUCHARDB, les losanges noirs et les triangles blancs, aux dates au radiocarbone conventionnelles de BOUCHS et BOUH96 respectivement. Les cercles représentent les dates par corrélation sporopollinique.
Age/depth curves of sediment of Lac du Triangle (a) and Petit Lac Bouchard (b). Black squares correspond to dates from accelerator mass spectrometry (AMS) for TRIANGLE or BOUCH96; white squares to AMS dates for BOUCAHRDB; black lozenges and white triangles, to standard radiocarbon dates for BOUCHS and BOUCH96. Circle are dates from pollen correlation.
Tableau II
Données chronologiques

Profondeurs ajustées par corrélation des carottes ; voir texte.
Le δ13C des dates TO- est normalisé à -25 ‰.
Abréviations des pièces macrofossiles : bra : bractée ; ecf : écaille de cône femelle ; ecm : écaille de cône mâle ; ffeu : fragment de feuille ; fru : fruit ; gra : graine ; mes : mésoblaste.
Dans le cas du Petit lac Bouchard (fig. 5b ; tabl. II), les dates étalonnées de 810 BP (7,5 cm), de 1860 BP (52,5 cm) et de 3275 BP (107,5 cm) de la carotte BOUCH96 furent rejetées car elles sont trop vieilles. La comparaison avec les dates obtenues sur les macrorestes terricoles révèle une contamination par du vieux carbone issu des carbonates répandus dans les roches régionales. Les autres dates de la carotte BOUCHS et celles de BOUCH96 ont été conservées puisque qu’elles sont en accord avec la profondeur des sédiments (fig. 5b). Bien que les dates de 8860 BP (290 cm) et 10 240 BP (325,5 cm) de BOUCHARDB supposent un TAS très lent à la base du profil, elles furent conservées puisqu’elles sont corroborées par les données du lac Davis (48° 58' 00" N ; 64° 38' 22" O ; altitude : 340 m ; Richard et al., 1997). Ce dernier présente en effet un âge de 10 200 BP à l’interface argile/gyttja et les courbes polliniques révèlent une toundra arbustive semblable à celle enregistrée au Petit lac Bouchard. Le déclin manifeste de la courbe pollinique de Tsuga canadensis à 175 cm correspondrait à 5400 BP. Le TAS fut calculé en appliquant une régression polynomiale d’ordre quatre ; cela minimise les changements abrupts. Un polynôme inférieur ou supérieur générait des âges déviant fortement des données. Le TAS ainsi obtenu présente des valeurs allant de moins de 0,02 cm/an à 0,095 cm/an, pour un taux moyen de 0,034 cm/an.
Données paléobotaniques
Les résultats des analyses sporopolliniques et macrofossiles ont été compilés à l’aide du logiciel Tilia/TiliaGraph (Grimm, 1991) et sont présentés sous forme de diagrammes de pourcentages (fig. 6) ou d’influx sporopolliniques (taux net d’accumulation : n grains/cm2/an, fig. 7), de mesures du diamètre équatorial des grains de pollen de Betula selon Marcoux et Richard (1993 ; fig. 8) et de concentration macrofossile (fig. 9). Trente-cinq taxons sporopolliniques faiblement représentés, soit 8 arbres, 6 arbustes, 11 herbes et 10 hors-total (aquatiques et ptéridophytes) ont été exclus des diagrammes polliniques.
Les zones d’assemblages polliniques (ZAP) ont été déterminées visuellement d’après les variations de la concentration pollinique totale et celles du pourcentage des taxons dominants ou caractéristiques, selon la procédure de Richard (1977), et de Labelle et Richard (1984) pour la Gaspésie. Les zones 1 (a et b) traduisent la phase non arboréenne de la colonisation végétale, les zones 2 (a et b), l’afforestation, et les zones 3 (a, b et c), la phase forestière (tabl. III et IV et fig. 7, 8 et 9).
Reconstitutions de la végétation
La végétation reflétée par les spectres polliniques des diagrammes en pourcentages (fig. 6a et 6b) est d’abord reconstituée par comparaison avec ceux produits par la végétation actuelle du Québec et des régions limitrophes (Richard, 1968, 1976, 1979, 1981 ; Webb et al., 1978 ; Comtois et Payette, 1984 ; Gajewski, 1991). Les données d’influx sporopollinique, indépendantes entre elles, sont utilisées pour évaluer les changements temporels dans l’abondance des populations végétales de chaque taxon. Elles dépendent aussi de la chronologie ; malgré le caractère uniforme des sédiments, les fluctuations de la courbe de la concentration pollinique du Petit lac Bouchard (fig. 6b) traduisent sans doute des changements du taux d’accumulation sédimentaire ignorés par notre modèle chronologique.
Sous réserve de ce qui précède, les valeurs d’influx de chaque taxon pollinique (ou sporal) sont comparables d’une couche sédimentaire à une autre, à condition que les processus d’accumulation soient restés les mêmes dans un même lac. Or l’accumulation sédimentaire entraîne une réduction progressive de l’épaisseur de la colonne d’eau et provoque l’étalement des apports polliniques sur une surface de taille croissante avec le temps, ce qui influence les influx. Leur interprétation doit donc tenir compte des phénomènes qui affectent la focalisation sédimentaire (Davis et al., 1984). L’intensité de cette focalisation peut notamment être modifiée par des changements du niveau d’eau des lacs.
La comparaison des influx d’un taxon donné d’un lac à un autre est plus hasardeuse. En effet, Davis et al. (1973) ont montré que les influx polliniques peuvent varier considérablement entre des lacs rapprochés, même bordés par une végétation similaire, en raison de la diversité des processus taphonomiques liés essentiellement à la morphométrie des bassins lacustres. Des bassins de forme et de taille semblables peuvent livrer des influx très différents.
C’est le cas de nos deux lacs (fig. 7a et 7b). L’influx pollinique total moyen est de 11 000 grains/cm2/an au Petit lac Bouchard (B), et de 7 000 grains/cm2/an au lac du Triangle (T), soit un rapport Bouchard/Triangle (B/T) de 1,6. L’influx moyen varie toutefois d’une zone pollinique à l’autre. Le rapport B/T tourne autour de 1,0 depuis près de 7000 ans (zones 3b et 3c), mais il se situait auparavant à 1,7 (zone 3a) ou à 2,2 (zone 2b). En zone 3a, le rapport B/T des influx de Betula est de 2,7 ; en zone 2b, celui d’Alnus type crispa est de 6,7. Ces différences peuvent donc être en grande partie attribuées à l’abondance même des populations de Betula spp. et d’Alnus type crispa autour du Petit lac Bouchard, mais une partie résulte d’une taphonomie différente en raison des changements de morphométrie, comme nous l’avons vu plus haut. L’examen des rapports d’influx pollinique de la pruche (Tsuga canadensis) permet de trancher.
En effet, l’allure des courbes polliniques de la pruche dans l’un et l’autre profil sédimentaire, tant en pourcentages qu’en influx, reflète très bien les changements de l’abondance de la pruche loin au sud et au sud-ouest et ce, durant tout l’Holocène. Comme la pruche est, de nos jours, absente de la Gaspésie (voir Scoggan [1950] et Rousseau [1974], contrairement à ce qui est illustré sur la carte de Fowells [1965], reprise par Farrar [1996] et par Thompson et al. [1999a, b]), les retombées éoliennes de son pollen, d’origine lointaine, doivent être semblables dans les deux lacs. On peut donc utiliser le rapport entre les influx de la pruche des deux sites afin d’évaluer, pour diverses périodes du passé, les éventuels biais ayant influencé l’incorporation sédimentaire du pollen au Petit lac Bouchard par rapport au lac du Triangle. Ainsi, pour la zone pollinique 3c, la plus récente, le rapport B/T est de 0,8 à 1,1 mais pour le début de la zone 3b, les influx polliniques de la pruche sont ~1,5 fois plus grands au Petit lac Bouchard qu’au lac du Triangle. Les deux lacs n’ont donc pas capté les apports polliniques de pruche de manière uniforme dans le temps, sans doute en raison de changements du niveau d’eau (voir plus loin). Les valeurs d’influx des autres taxons peuvent donc être corrigées d’un site à l’autre en tenant compte du rapport du pollen de Tsuga dans les couches où il est bien représenté, ce qui permet d’évaluer les éventuels changements survenus dans la taille de leurs populations.
Histoire de la végétation locale et riveraine
La végétation locale est celle qu’on retrouve dans les eaux du lac et la végétation riveraine, extra-locale sensu Janssen (1966), est celle qui occupe les berges environnantes. Leur histoire est reflétée par les courbes sporopolliniques et macrofossiles des plantes aquatiques, des ptéridophytes, des mousses, des herbes mais aussi par celles de certains arbres et arbustes souvent riverains ou tourbicoles tels Picea mariana, Larix laricina, Myrica gale et Alnus incana. Pour ces derniers, la difficulté consiste à identifier la source, extra-locale (riveraine) ou régionale (terricole mésophile) de leur pollen, car ce ne sont souvent pas des plantes riveraines strictes. À l’inverse, des arbres, arbustes ou buissons mésophiles peuvent croître tout près des rives d’un lac sans pour autant être inféodées au milieu riverain. Pour ces raisons, nous donnerons la priorité aux données macrofossiles dans la reconstitution de la dynamique du milieu lacustre et du milieu riverain, bien que la représentation macrofossile des plantes soit souvent plus aléatoire et moins étendue que la représentation pollinique (Birks, 1980).
Les reconstitutions s’appuient également sur les données sédimentologiques, puisque la densité de la végétation riveraine joue un rôle important sur l’érosion des terrains périphériques que traduit le type de sédiment accumulé dans les bassins lacustres.
Phase initiale à berges dénudées
Tant au lac du Triangle qu’au Petit lac Bouchard, les premiers sédiments accumulés, argilo-silteux, finement sableux par endroits témoignent de berges alors largement dénudées. L’érosion pelliculaire y a agi sans contrainte, entraînant les particules fines dans les plans d’eau. En l’absence de datation directe (tabl. II, fig. 5), nous évaluons la durée de cette phase à quelques siècles tout au plus, mais la fin, apparemment synchrone dans les deux lacs, est autour de 10 750 BP.
Au lac du Triangle, seules des graines de Ranunculus sceleratus sont identifiées à la base des sédiments (fig. 9a). Celles de Potamogeton sp. suivront, accompagnées de tiges de mousses et de restes de Salix herbacea, de Dryas integrifolia et de Betula glandulosa dans la partie supérieure de l’argile silteuse. Un fragment d’aiguille de Picea mariana fut trouvé à la toute base du profil, témoignant soit de la présence de l’épinette noire dès la déglaciation, soit d’un apport lointain par l’entremise des oiseaux (Watts, 1978 ; Birks, 1980). Au Petit lac Bouchard, l’abondance des macrorestes du bouleau glanduleux (fig. 9b) reflète soit une plus grande abondance de cet arbuste sur les berges, soit une meilleure dispersion des graines dans le lac. Des restes de Cypéracées (Carex sp.) et des oospores de Chara ou de Nitella y furent identifiés. La représentation macrofossile est particulièrement faible dans les sédiments inorganiques des deux lacs ; les plantes devaient donc être assez rares ou éloignées des berges, ce qui impliquerait un niveau d’eau élevé à cette époque. Toutefois, la présence de sable, en l’absence d’affluents, et compte tenu du faible relief autour des lacs, ne permet pas de supposer un niveau plus élevé que le plein bord moderne.
Dans ces sédiments inorganiques, les assemblages sporopolliniques (fig. 6 et 7 : zones 1, 1a, 1b) sont caractérisés par une représentation importante des herbes (Cypéracées, Poacées, Artemisia) et des spores de Ptéridophytes (fougères et lycopodes). Oxyria digyna est présent au lac du Triangle et l’arbuste basiphile Shepherdia canadensis, accompagné de saules, croissait près des deux lacs. Les taxons polliniques herbacés sont plus nombreux que les macrorestes, mais leur représentation mutuelle (%) est favorisée par l’absence d’un couvert forestier régional dont l’apport pollinique viendra, plus tard, masquer celui des plantes herbacées.
Le pollen de Betula atteint des pourcentages de 50 à 70 (fig. 6). D’après la petite taille des grains (fig. 8), il s’agit de bouleaux arbustifs. Les influx sont toutefois très faibles (fig. 7). Les macrorestes de Betula glandulosa (fig. 9a), n’abondent que durant le maximum de pourcentage de Betula, témoignant de la présence de petites populations sans doute riveraines. Ces données permettent déjà l’identification d’une toundra arbustive à bouleau glanduleux occupant les versants de manière éparse, puisqu’elle ne peut s’exprimer palynologiquement quand des arbres, grands producteurs de pollen sont présents. Au lac du Triangle et au Petit lac Bouchard, Salix herbacea, Dryas integrifolia (fig. 9) et Shepherdia canadensis (fig. 6) devaient être proches des rives.
Phase à berges fixées par la végétation
Depuis 10 750 ans environ, le Petit lac Bouchard et le lac du Triangle accumulent de la matière organique (fig. 3). Dans le premier cas, il s’agit jusqu’à nos jours d’une gyttja comprenant ~40 % de matière organique et très peu ou pas de carbonate de calcium ; dans le deuxième cas, une telle gyttja encore plus riche en matière organique (~70 %) ne s’est accumulée que depuis 3000 ans. Au lac du Triangle, la période entre 10 750 et 3000 BP est caractérisée par des sédiments carbonatés : marne, gyttja et gyttja marneuse plus ou moins finement litées (fig. 3), riches en gastéropodes, pélécypodes et ostracodes (fig. 9a). La précipitation des carbonates est un processus complexe (Dean et Schwalb, 2002) qui, lorsqu’elle survient dans les lacs de la Gaspésie, touche surtout les zones littorales, dans les conditions hydrogéologiques et climatiques actuelles. Puisque la carotte étudiée provient de la zone centrale du bassin, toujours dans la plus grande profondeur d’eau, les variations sédimentaires enregistrées entre 10 750 et 3000 ans au lac du Triangle furent sans doute entraînées par des niveaux d’eau souvent bas, mais fluctuants. Ainsi s’explique l’abondance des Turbellaria et de Cristatella mucedo, et probablement aussi celle des restes de mousses et de fragments ligneux bien qu’ils ne soient pas exclusifs aux sédiments marneux (fig. 9a). L’accumulation organique est, dans les deux cas, attribuée à un accroissement de la production planctonique et à la densification de la végétation environnante qui, en créant un humus à peu près continu autour des lacs, limita l’érosion et, partant, les apports inorganiques.
Figure 6
a

Lac du Triangle
Lac du Triangle
b
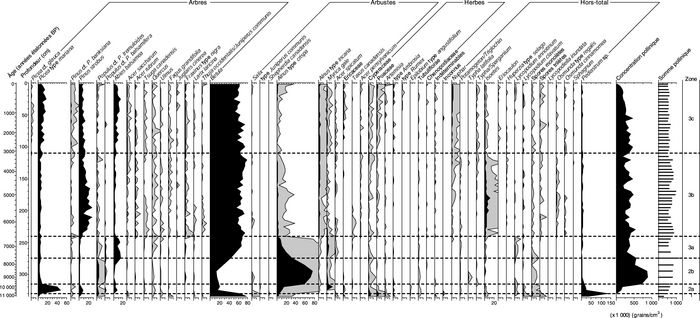
Petit Lac Bouchard
Petit Lac Bouchard
Diagrammes des pourcentages polliniques : (a) lac du Triangle (48° 42’ 36’’ N, 65° 24’ 50’’ O), (b) Petit lac Bouchard (48° 51’ 14’’ N, 64° 35’ 52’’ O).
Pollen and spore percentages diagrams: (a) Lac du Triangle (48° 42’ 36’’ N, 65° 24’ 50’’ W), (b) Petit Lac Bouchard (48° 51’ 14’’ N, 64° 35’ 52’’ W).
Figure 7
a

Lac du Triangle
Lac du Triangle
b

Petit Lac Bouchard
Petit Lac Bouchard
Diagrammes des influx polliniques : (a) lac du Triangle, (b) Petit lac Bouchard. Les courbes d’influx polliniques sont brutes, non corrigées.
Pollen accumulation rates diagrams: (a) Lac du Triangle, (b) Petit Lac Bouchard. Pollen accumulation rates curves were made with raw data (non-corrected data).
Figure 8
a
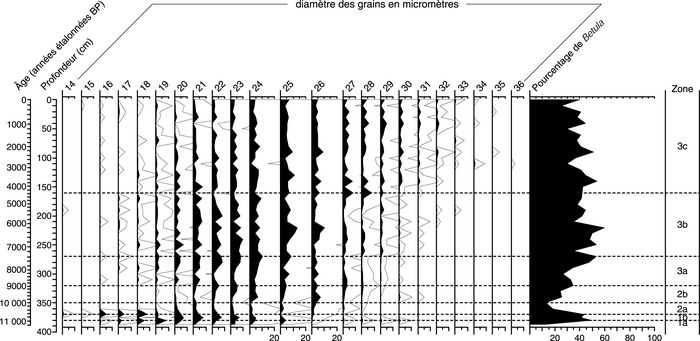
Lac du Triangle
Lac du Triangle
b
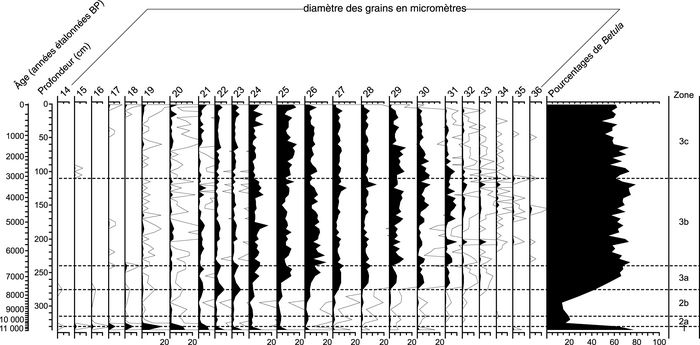
Petit Lac Bouchard
Petit Lac Bouchard
Courbes des pourcentages de Betula selon le diamètre équatorial des grains de pollen du lac du Triangle (a) et du Petit lac Bouchard (b).
Betula pollen grain size percentages diagrams of Lac du Triangle (a) and Petit Lac Bouchard (b).
Figure 9
a

Lac du Triangle
Lac du Triangle
b

Petit Lac Bouchard
Petit Lac Bouchard
Diagrammes des concentrations macrofossiles : a) lac du Triangle, b) Petit lac Bouchard (ath : anthère, bra : bractée, brf : bractée femelle, cac : coussinet d’aiguille de conifère, cap : capsule, col : colonie, com : cône mâle, ecf : écaille de cône femelle, ecm : écaille de cône mâle, ecr : écorce, eph : ephippia, feu : feuille, ffeu : fragment de feuille, fgra : fragment de graine, fru : fruit, gem : gemmule, gra : graine, meg : mégaspore, mes : mésoblaste, oosp : oospore, rac : racine, ram : rameau, rhi : rhizome, scl : sclérote, sta : statoblaste, tif : tige feuillée, tig : tige).
Macrofossil-concentration diagrams: a) Lac du Triangle, b) Petit Lac Bouchard (ath: anther, bra: bract, brf: female bract, cac: coniferous sterigmata, cap: capsule, col: colony, com: male cone, ecf: female cone scale, ecm: male cone scale, ecr: bark, eph: ephippia, feu: leaf, ffeu: leaf fragment, fgra: seed fragment, fru: fruit, gem: gemmule, gra: seed, meg: megaspore, mes: mesoblast, oosp: oospore, rac: root, ram: branchlet, rhi: rhizome, scl: sclerotium, sta: statoblast, tif: leafy stem, tig: stem).
Tableau III
TRIANGLE : Zonation pollinique régionale

CP : concentration pollinique (grains/cm3) ; PA/T : rapport du pollen des arbres (incluant le genre Betula) au total des grains.
D’après les données macrofossiles (fig. 9), les Characées (Chara/Nitella), déjà présentes au Petit lac Bouchard, abondèrent dans les eaux des deux lacs durant toute la phase d’accumulation organique. Najas flexilis et les potamots (Potamogeton type pusillus) se sont multipliés dans le Petit lac Bouchard, surtout entre 7200 et 2500 BP, tandis qu’ils furent représentés au lac du Triangle avant et après ces dates. Les restes d’autres plantes aquatiques sont rares, sauf ceux de Nuphar variegatum, qui sont présents au Petit lac Bouchard depuis environ 4000 ans. Quant à la représentation pollinique des plantes aquatiques, elle est particulièrement faible et sporadique au lac du Triangle (fig. 9a). Elle est beaucoup plus soutenue au Petit lac Bouchard où est enregistré le développement successif des populations de Potamogeton/Triglochin d’abord, de Nuphar et de Typha/Sparganium ensuite, puis celui des Eriocaulon sp. Fait remarquable pour une carotte en position centrale, les microspores d’Isoetes y affichent une forte représentation entre 6600 et 3000 ans avant l’actuel (fig. 6b), attribuée à un abaissement du niveau d’eau.
La végétation herbacée et arbustive riveraine est très peu représentée et de manière très discontinue par les macrorestes (fig. 6), tant chez les Cypéracées (Carex, Cyperus, Scirpus) que chez les Éricacées (Chamaedaphne calyculata, Andromeda glaucophylla) ; de même pour Myrica gale et les aulnes (A. incana, A. viridis crispa) pourtant abondants aujourd’hui sur les berges. Pour ces taxons et pour les Poacées, les données polliniques traduisent sans doute mieux que les macrorestes les changements dans l’abondance des populations riveraines durant la phase de sédimentation organique des deux lacs (fig. 6 et 7). On note ainsi l’ancienneté des aulnaies à aulne rugueux (A. incana) et le développement du myrique baumier (Myrica gale) durant les derniers millénaires, mais ce dernier a aussi été abondant entre 10 000 et 9500 ans et entre 7500 et 6500 ans avant nos jours au Petit lac Bouchard. Cette représentation pollinique trimodale pourrait refléter d’importantes fluctuations des myriçaies autour de ce lac, peut-être en liaison avec des changements du niveau d’eau élargissant ou rétrécissant les berges. Une représentation pollinique accrue de Myrica gale traduirait ainsi des niveaux lacustres élevés. Ces arbustaies riveraines hébergeaient des plantes herbacées diverses, mais leur identification à l’espèce est rarement possible, sauf pour les Lycopodiacées (fig. 6).
Tableau IV
BOUCHARD : Zonation pollinique régionale

CP : concentration pollinique (grains/cm3) ; PA/T : rapport du pollen des arbres (incluant le genre Betula) au total des grains.
Deux traits des diagrammes sporopolliniques sont remarquables. Le premier, au lac du Triangle, touche la période entre 6500 et 4000 BP durant laquelle le pollen des Cypéracées et les spores de Sphagnum affichent des valeurs élevées (fig. 6a et 7a). Cette expression palynologique, étayée par les macrorestes de sphaignes (fig. 9a), serait le reflet du développement centripète d’une tourbière riveraine en réponse à un abaissement du niveau du lac. Le second, au Petit lac Bouchard, concerne les spores d’Osmunda regalis et, dans une moindre mesure, celles d’Osmunda cinnamomea, deux fougères hygrophiles qui, entre 6600 et 3000 ans avant l’actuel montrent un accroissement sensible de leur représentation (fig. 6b). Encore ici, un abaissement du niveau du lac a sans doute entraîné un rapprochement des populations de fougères, favorisant leur enregistrement sporal au centre du lac. Ces déductions fondées sur les plantes riveraines corroborent donc celle tirée des spores d’Isoetes, une plante aquatique.
Durant cette phase organique, l’abondance réelle (influx ; fig. 7) des Pediastrum sp. diminue irrégulièrement au Petit lac Bouchard, traduisant ainsi une turbidité croissante et un appauvrissement graduel des eaux en éléments nutritifs (Smol, 1990 ; Nielsen et Sorensen, 1992). Au lac du Triangle, cette tendance générale n’est pas enregistrée mais, durant l’épisode de tourbière riveraine, la représentation des Pediastrum chute fortement (fig. 7a), tel qu’attendu dans des eaux alors sans doute plus acides et plus riches en matière organique dissoute.
Chez les arbres enfin, les macrorestes (fig. 9) montrent que l’épinette noire (Picea mariana) fut présente très tôt près des berges durant la phase d’accumulation organique, tant au lac du Triangle qu’au Petit lac Bouchard. Il est d’ailleurs remarquable que ses macrorestes soient abondants en même temps que ses pourcentages polliniques maximaux (fig. 6, zone 2a), sans que ses influx maximaux ne soient pas encore atteints (fig. 7). Le sapin baumier (Abies balsamea) était alors présent au Petit lac Bouchard aussi, mais il semble n’avoir jamais colonisé les abords mêmes du lac du Triangle (fig. 9). Le mélèze (Larix laricina), sans doute présent dès 10 000 à 9000 BP dans les deux sites n’a vu ses populations riveraines abonder que vers 6600 BP au Petit lac Bouchard. Il a probablement constitué une ceinture plus ou moins discontinue autour des lacs durant tout l’Holocène. Les variations dans l’abondance des macrorestes du mélèze traduisent des variations dans la taille des populations ou alors dans leur distance par rapport au centre des lacs en liaison avec les changements du niveau de l’eau, ce que corrobore assez étroitement le pollen (fig. 6 et 7).
On ne peut rien tirer de la représentation macrofossile de Betula papyrifera au lac du Triangle (fig. 9a). Au Petit lac Bouchard, le bouleau blanc était proche des rives dès 9400 BP et reste abondamment représenté jusque vers 4000 ans avant nos jours (fig. 9b). Dès 4800 BP, ses restes sont remplacés par ceux de Betula alleghaniensis jusqu’à l’actuel. Il s’agit seulement des populations riveraines car, comme le montre la figure 8b, les grains de pollen attribuables au bouleau jaune abondent depuis au moins 7500 BP. Des populations de pin blanc (Pinus strobus) occupent aussi les abords du Petit lac Bouchard depuis 7600 ans, mais apparemment en moins grand nombre depuis 3200 ans comme le montre la rareté relative des écailles de cônes mâles (fig. 9b). Il se peut que la représentation de ces pièces macrofossiles ait été favorisée alors que le niveau d’eau était bas.
Les 3700 dernières années sont caractérisées par une recrudescence inégale mais sensible de la représentation pollinique ou macrofossile des Poacées, des Éricacées, (Chamaedaphne calyculata), du myrique baumier, de l’aulne rugueux et des spores de sphaignes, traduisant ainsi un entourbement graduel des berges en réponse à une remontée progressive du niveau d’eau des deux lacs. Une part de la représentation pollinique de Picea mariana reflète sans doute une abondance accrue en bordure du lac, mais probablement aussi sur les versants autour des deux lacs.
Histoire de la végétation régionale
La végétation régionale est définie comme étant celle qui occupe les terres fermes au-delà des zones riveraines autour de chacun des lacs. Par conséquent, la reconstitution de cette végétation s’appuie essentiellement sur les valeurs des divers taxons arborescents, arbustifs et herbacés terricoles affichées dans les spectres (ou assemblages) polliniques. Toutefois, on l’a vu, certains taxons, surtout des arbres et des arbustes, peuvent indifféremment occuper les rives ou les terres fermes. Les reconstitutions de la végétation ligneuse régionale s’appuieront donc aussi sur celles de la végétation riveraine et des abords immédiats des lacs pour ce qui est des taxons terricoles représentés par les macrorestes.
Les assemblages polliniques du lac du Triangle et du Petit lac Bouchard (tabl. III et IV) ont permis d’identifier les trois grandes phases généralement reconnues en ce qui a trait au développement postglaciaire de la végétation régionale : 1) une phase initiale non arboréenne, 2) une phase d’afforestation et 3) une phase forestière. Les reconstitutions de la végétation de chaque site seront traitées en parallèle, au sein de chaque zone pollinique homologue, pour faire ressortir les ressemblances et les différences dans la dynamique du couvert végétal entre la côte orientale et l’intérieur des terres de la Gaspésie.
Phase non arboréenne (zone 1)
À la zone pollinique 1, antérieure à 10 640 BP au lac du Triangle et à 10 750 BP au Petit lac Bouchard, correspond une phase initiale de végétation dépourvue d’arbres, la toundra. Seule une pièce d’épinette noire a été trouvée dans les sédiments du lac du Triangle (fig. 9a) et la concentration pollinique y est très faible (fig. 6), comme les influx qui en dérivent, compte tenu du taux d’accumulation sédimentaire (fig. 7). Le pollen arboréen est entièrement attribué à des apports lointains à partir de régions boisées situées loin au sud et au sud-ouest. Cette phase n’est représentée que par deux ou trois spectres polliniques à la base des carottes et semble ne couvrir qu’une courte période, difficilement évaluable, de l’ordre de quelques siècles ou décennies.
Au lac du Triangle, les assemblages polliniques identifient une toundra herbacée quasi-désertique (sous-zone 1a) suivie d’une toundra arbustive éparse à bouleau glanduleux (sous-zone 1b), correspondant respectivement aux sous-zones 1b et 1c identifiées par Savoie (1978) et Labelle et Richard (1981). Seule la toundra arbustive est présente au Petit lac Bouchard. Comme on l’a vu pour la reconstitution de la végétation riveraine, c’est l’absence d’arbres dans les paysages environnants qui a permis l’enregistrement de pourcentages élevés du pollen de Betula glandulosa, malgré des influx polliniques très faibles.
Durant la phase non arboréenne, le couvert végétal devait donc être extrêmement réduit sur les versants des lacs étudiés, et constitué des mêmes plantes que celles occupant les rives : des Ericaceae, Cyperaceae, Poaceae, Asteraceae (Tubulifloreae), Artemisia, Ambrosia, Caryophyllaceae, Thalictrum sp., Epilobium sp., Shepherdia canadensis et des taxons à affinité arctique-alpine tels Oxyria digyna, Salix herbacea, Dryas integrifolia. Acer spicatum représente sans doute un apport pollinique lointain, comme pour les arbres. D’autres plantes ont pu être présentes, mais les données sont muettes à ce propos.
Phase d’afforestation (zone 2)
Cette phase se caractérise par l’établissement des arbres vers 10 700 BP, tant au lac du Triangle qu’au Petit lac Bouchard, entraînant progressivement la fermeture du couvert arborescent (fig. 6 ; tabl. III et IV). L’afforestation comprend deux étapes : 1) la multiplication des premières populations arborescentes (sous-zone 2a) et 2) une période de dominance du pollen d’Alnus type crispa (sous-zone 2b). La durée de l’afforestation est estimée à 1650 ans au lac du Triangle et à 3100 ans au Petit lac Bouchard. Une telle différence ne peut être attribuable aux seules imprécisions dans les chronologies. Par ailleurs, l’afforestation est caractérisée par une forte incidence des incendies naturels, les influx maximaux de charbons de bois microscopiques (fig. 4) étant associés à la phase à aulne crispé.
Les spectres polliniques de pourcentages (fig. 6a et 6b) montrent une succession très nette de Picea mariana (sous-zone 2a) et d’Alnus crispa (sous-zone 2b) dans la représentation pollinique. Les assemblages correspondent d’abord à ceux actuellement produits par la végétation du domaine de la Pessière noire (sous-zone 2a), puis à ceux produits dans la partie nordique du domaine de la Pessière à cladines ou même celui de la Toundra forestière du nord du Québec (sous-zone 2b) (Richard, 1981 ; Gajewski, 1991). Le tremble (Populus tremuloides) et le peuplier baumier (P. balsamifera) ont accompagné l’épinette noire autour des deux lacs, avec sans doute le genévrier commun (Juniperus communis). Au Petit lac Bouchard, le tremble est resté bien représenté durant la phase à aulne crispé.
La présence de macrorestes d’Abies balsamea en sous-zone 2a au Petit lac Bouchard étaye la courbe pollinique du sapin baumier qui enregistre son implantation dès le début de l’afforestation dans cette région. C’est là un caractère original qui rapproche ce site de ceux de la Basse-Côte-Nord (Mott, 1976 ; Lamb, 1980 ; King, 1986 ; Richard, 1993), les populations de sapin baumier s’établissant généralement après celles de l’épinette noire dans la séquence générale de l’afforestation ailleurs au Québec (Richard, 1977, 1994). Bien que la représentation pollinique de Betula (cf. B. papyrifera, d’après la taille des grains de pollen ; fig. 8) soit modeste dans les deux sites, les données macrofossiles du Petit lac Bouchard traduisent son abondance près des rives durant la deuxième partie de l’afforestation (sous-zone 2b). Larix laricina, le mélèze, y était aussi présent.
Les courbes des influx polliniques des taxons précités (fig. 6) éclairent les modalités de l’afforestation. Le déploiement initial des épinettes noires est confirmé durant les sept premiers siècles de la sous-zone 2a, mais le déclin des populations vers 10 000 BP fut beaucoup moindre qu’il le semble à partir des seuls pourcentages polliniques, qui passent de 40 à 5-10 % selon le lac (fig. 5). Les données d’influx du Petit lac Bouchard confirment le rôle important du genre Populus (sans doute le tremble) durant la dominance de l’aulne crispé (sous-zone 2b). Elles montrent que le sapin baumier (Abies balsamea) y a alors aussi connu un fléchissement de sa population, contrairement à ce qui a été observé à la même époque au lac du Triangle, où ses influx traduisent une croissance apparemment exponentielle. Ces données traduisent donc une différence majeure dans l’afforestation autour des deux sites, une régression des conifères étant manifeste au Petit lac Bouchard mais n’ayant touché que l’épinette noire au lac du Triangle, au coeur de la Gaspésie.
D’après les influx polliniques d’Alnus crispa, les paysages autour du lac du Triangle et du Petit lac Bouchard devaient être, à partir de 9700 BP, couverts d’arboraies ouvertes arbustives, voire d’arbustaies denses arborées, selon la dominance respective de l’épinette noire et de l’aulne crispé, essentiellement. De tels paysages végétaux régionaux, admettant aussi le tremble, le peuplier (?) et le bouleau blanc, n’existent pas actuellement au Québec-Labrador.
Les apports polliniques de Tsuga canadensis enregistrés au Petit lac Bouchard et au lac du Triangle sont alors semblables (B/T = ~1). Les influx de Picea mariana, de Pinus strobus, d’Abies balsamea et de Quercus durant l’afforestation peuvent donc être comparés dans les sédiments de l’un et l’autre lac, et le contraste entre les influx de Populus tremuloides et d’Alnus crispa s’en trouve ainsi confirmé. Les populations d’aulne crispé devaient véritablement abonder autour du Petit lac Bouchard durant la sous-zone 2b.
La standardisation entre les lacs (rapport B/T, voir plus haut) met en relief l’importance relative des influx du pin gris (Pinus banksiana) au lac du Triangle, durant l’afforestation. Ces données indiquent qu’il a participé à la flore dendrologique du centre de la péninsule de 10 000 à 4000 ans avant nos jours (fig. 7a). L’établissement du pin gris durant la phase d’afforestation a sans doute été facilité par le grand nombre de feux (fig. 4), auxquels l’essence est particulièrement bien adaptée. Il est probable, en outre, que les feux aient entretenu le caractère ouvert de la sylve, freinant l’établissement de la forêt durant l’afforestation.
Phase forestière (zone 3)
Cette phase correspond à la mise en place de forêts fermées, de structure semblable à celles des paysages actuels. Les assemblages polliniques (fig. 6), dominés essentiellement par le pollen des bouleaux arborescents (tabl. III et IV), montrent d’abord une forte représentation du sapin baumier (3a), puis du pin blanc (3b), puis une recrudescence de la représentation de l’épinette noire et du sapin (3c). Durant cette phase forestière, les influx polliniques corroborent généralement la représentation exprimée en pourcentages. Les variations des courbes polliniques des divers taxons y sont toutefois différemment exprimées. Les influx des taxons peuvent être comparés d’un lac à l’autre, notamment en zones 3c, mais le rapport B/T de la pruche (1,5) permet de déceler un important enrichissement taphonomique au Petit lac Bouchard à fin de la zone 3a et au début de la zone 3b. À la lumière des reconstitutions des végétations lacustre et riveraine, cet enrichissement correspondrait à l’abaissement du niveau d’eau des lacs, favorisant surtout les grains de pollen à ballonnets qui se concentrent dans les zones littorales (Davis et Brubaker, 1973).
Betula-Abies
Les assemblages polliniques traduisent la constitution de sapinières régionales entre 9000 et 7500 BP autour du lac du Triangle, mais il faudra attendre 7700 BP pour qu’elles s’étalent autour du Petit lac Bouchard, dominant les paysages végétaux jusque vers 6700 BP. Le sapin est polliniquement sous-représenté (Richard, 1976 ; Delcourt et al., 1984), mais les pourcentages et les influx qu’il atteint en sous-zone 3a sont suffisamment élevés (fig. 6 et 7) pour en déduire sa dominance. Sa présence à proximité du lac du Triangle vers 7500 BP est établie par les macrorestes (fig. 9a). Durant cette phase de sapinières, l’aulne crispé décline à mesure que les forêts se densifient. Il a dû jouer un rôle dans la fixation de l’azote atmosphérique, enrichissant les sols (Huss-Danell et al., 1999). Comme les influx de charbons microscopiques diminuent en même temps (fig. 4), il est probable que la diminution de la fréquence des feux ait favorisé la fermeture de la sylve et le développement du sapin baumier.
Le sapin était accompagné de l’épinette noire. Cette essence a progressé au sein des sapinières situées autour du lac du Triangle (fig. 6a et 7a) et a maintenu des populations non négligeables, sans doute riveraines, autour du Petit lac Bouchard (fig. 6b et 7b). L’abondance du peuplier faux-tremble a décliné, mais il devait rester important dans la sylve, compte tenu de sa sous-représentation pollinique notoire (Mott, 1978). Au lac du Triangle, le bouleau blanc (Betula papyrifera, d’après le diamètre des grains ; fig. 8a), ne devait pas être très abondant au sein des sapinières, car c’est un genre fortement surreprésenté par le pollen (Richard, 1968). Il en est allé autrement au Petit lac Bouchard où les pourcentages de Betula sont les doubles de ceux du lac du Triangle en sous-zone 3a. Les assemblages polliniques correspondent plus étroitement à ceux du domaine de la sapinière à bouleau blanc, mais il n’est pas exclu que le bouleau jaune (Betula alleghaniensis) se soit installé dans les environs dès 7500 BP (fig. 7b). Au lac du Triangle, c’est au domaine de la Sapinière à épinette noire que le couvert végétal d’alors appartenait, mais l’épinette noire devait y être moins abondante que plus tard (en sous-zone 3c) et le pin gris, en nombre modeste mais présent.
Betula-Pinus
Entre 7400 et 3500 BP au lac du Triangle, et entre 6600 à 3100 BP au Petit lac Bouchard, les assemblages polliniques sont caractérisés par une augmentation notable du pin blanc (Pinus strobus) dont les pourcentages atteignent ou dépassent 15 (fig. 6). L’aire de peuplement du pin blanc se limite aujourd’hui au pourtour de la Gaspésie, où il ne paraît avoir été ni abondant ni répandu, même avant l’exploitation forestière et les défrichements du XIXe siècle (Lortie, 1979). Le pin blanc fut plus abondant durant l’Holocène moyen dans la région du Petit lac Bouchard, mais il n’a pas dû coloniser l’intérieur de la péninsule. Au lac du Triangle, sa représentation en pourcentage est attribuable à des apports polliniques lointains et non à des populations régionales ; les influx polliniques y sont en effet moindres qu’au Petit lac Bouchard (fig. 7), même lorsque ces derniers sont standardisés d’un lac à l’autre par le truchement des influx de la pruche. Ses pourcentages élevés entre 6000 et 4000 BP correspondent à la phase d’entourbement pendant que le niveau lacustre était bas et sont ainsi attribuables à un changement taphonomique (Davis, 1968, 1973 ; Davis et Brubaker, 1973 ; Davis et al., 1984). Le minimum enregistré dans les pourcentages et les influx de Betula durant à peu près la même période a sans doute la même origine car il est étroitement centré sur le maximum des spores de sphaignes (fig. 7a), indicateur, en l’occurrence, d’un bas niveau lacustre.
D’après les pourcentages et les influx (fig. 6 et 7), la représentation pollinique du sapin baumier décroît dans les deux sites, mais plus fortement au Petit lac Bouchard. Autour du lac du Triangle, l’épinette noire a de façon générale multiplié sa population, malgré des fluctuations apparemment importantes d’après les influx (fig. 7a) ; autour du Petit lac Bouchard, elle l’a simplement maintenue (fig. 7b). Des variations dans les populations riveraines, en relation avec des changements du niveau lacustre et du taux d’accumulation sédimentaire, sont sans doute responsables des variations d’influx enregistrées. Les phénomènes taphonomiques sont probablement responsables des variations dans les influx polliniques du genre Betula aussi, mais les assemblages pris dans leur ensemble (fig. 6), si l’on tient compte du diamètre des grains de Betula (fig. 8), permettent de déduire que la végétation autour du lac du Triangle appartenait alors au domaine de la Sapinière à épinette noire et celle des environs du Petit lac Bouchard, au domaine de la Sapinière à bouleau jaune, voire à celui de la Bétulaie jaune à sapin (Saucier et Grondin, 1996). Les paysages de l’est de la Gaspésie relevaient donc alors de la Forêt mixte, formation végétale intermédiaire entre la Forêt boréale et la Forêt tempérée.
La flore arboréenne était sans doute déjà aussi diversifiée qu’actuellement dans la région de Gaspé, avec des frênes noirs et des ormes d’Amérique sur les dépôts alluviaux fins, des érables rouges dans les stations perturbées, et de petites populations de chêne rouge et d’érable à sucre sur les versants (Majcen, 1981). Ces essences semblent toutefois n’avoir jamais abondé dans le bassin versant du Petit lac Bouchard. Il est probable qu’elles se soient implantées dans les sapinières à bouleau jaune régionales dès le début de l’Holocène moyen et pour ce qui est de l’érable à sucre, qu’il y ait connu un développement réduit mais continu.
Le cas du pin gris est intéressant. L’essence est actuellement absente de la flore gaspésienne (Scoggan, 1950 ; Rousseau, 1974), mais d’après les influx polliniques (fig. 7a), elle a dû coloniser la région du lac du Triangle durant l’afforestation et y a persisté durant l’Holocène moyen, pendant que les feux étaient très peu fréquents dans les deux régions (fig. 4). Curieusement, le Petit lac Bouchard n’a enregistré des influx plus élevés de pollen de pin gris que plus tard, durant l’époque de la sous-zone 3c. Nous croyons que l’essence n’a jamais colonisé les marges gaspésiennes et que la recrudescence des apports polliniques éoliens lointains traduit des changements dans la direction des vents dominants de printemps dans la région orientale de la péninsule. C’est là une hypothèse ad hoc qu’il conviendra de vérifier.
Parmi les buissons et les arbustes, Taxus canadensis est bien représenté au début de la période. Il fait actuellement partie du cortège floristique de la sapinière à bouleau jaune et de la sapinière à bouleau blanc, pouvant être très abondant en fin de succession (Saucier et Grondin, 1996). C’est un buisson très apprécié des cerfs de Virginie, mais il supporte mal les feux de forêt (Heinselman, 1981). Comme il est sous-représenté par son pollen de nos jours, les valeurs qu’il a atteintes au début de l’époque de la sous-zone 3b traduisent sans doute une forte abondance dans les forêts, probablement en raison du faible nombre des feux durant une longue période (fig. 4). La présence de pollen du chèvrefeuille (Lonicera), du sumac vinaigrier (Rhus typhina), de la viorne trilobée (Viburnum trilobum) et de l’érable de Pennsylvanie (Acer pensylvanicum) (fig. 6b) atteste d’un enrichissement dans la flore arbustive au Petit lac Bouchard.
Betula-Picea-Abies
Depuis 3500 BP au lac du Triangle, et depuis 3100 BP au Petit lac Bouchard, les forêts régionales ont connu un accroissement marquant de l’abondance du sapin baumier, mais aussi de l’épinette noire, surtout au coeur de la péninsule. Au lac du Triangle, la végétation régionale s’est progressivement constituée en domaine de la Pessière noire à sapin baumier, et le pin gris est disparu. L’épinette blanche (Picea glauca) s’est par contre développée dans la région côtière (fig. 6b). Durant ce temps, le nombre des feux de forêts s’est accru, un phénomène général dans l’ensemble du Québec (Carcaillet et Richard, 2000).
Le bouleau blanc, déjà rare autour du lac du Triangle, a vu sa population décliner. Nous n’expliquons pas l’accroissement de la représentation des grains de Betula à gros diamètre (>29 μm) au lac du Triangle (fig. 8a) durant cette période, si ce n’est par un effet taphonomique lié au redépôt de grains préservés dans l’humus des sols environnants en raison du rehaussement du niveau lacustre depuis environ 4000 ans. L’augmentation de la proportion des grains corrodés et de fragments de grains (Cushing, 1967) appuie cette déduction (Asnong, 2001). Le milieu de conservation, différent, aurait alors probablement entraîné un gonflement des grains de pollen (Comtois, 1981). Si tel fut le cas, cela a dû toucher aussi les grains de Betula enregistrés au Petit lac Bouchard durant la même période, ce qui aurait pour effet d’accentuer le contraste entre les sous-zones 3b et 3a en ce qui a trait à la réduction de la représentation des grains de pollen de Betula de forte taille (fig. 8b). Ainsi, autour du Petit lac Bouchard, les données touchant Betula montrent une importante régression dans l’abondance du bouleau jaune, bien que des populations aient persisté proche des rives, d’après l’examen des macrorestes (fig. 9b).
Selon les données du Petit lac Bouchard, les populations de pin blanc ont décliné depuis environ 4000 ans dans la grande région de Gaspé. Celles de l’érable à sucre (Acer saccharum) aussi, depuis 3000 ans. Dans les deux régions, le peuplier faux-tremble fut moins abondant qu’auparavant durant l’Holocène supérieur, sa représentation pollinique diminuant encore plus tôt (5500 BP) au lac du Triangle.
En définitive, dans les deux régions, la fin de l’Holocène est caractérisée par un important enrésinement des paysages végétaux et, pour la région orientale, par une diminution de l’abondance de la plupart des essences relativement thermophiles, y compris le bouleau jaune.
Discussion
Changements du niveau des lacs
Une partie des transformations enregistrées dans la végétation lacustre et riveraine traduit des changements des niveaux d’eau des lacs qu’il convient de commenter, car ils reflètent sans doute des modifications du bilan hydrique à l’échelle régionale. À partir des données autécologiques des plantes aquatiques et de la microfaune lacustre représentées dans les sédiments (Asnong, 2001), les indications suivantes peuvent être déduites (fig. 10).
Au lac du Triangle et au Petit lac Bouchard, le niveau d’eau était élevé avant 10 750 BP. Selon les assemblages, la colonne d’eau faisait alors au moins 2 m au centre de chacun des lacs et pouvait atteindre 6 m au lac du Triangle et 5,3 m au Petit lac Bouchard (Hannon et Gaillard, 1997 ; Dieffenbacher-Krall et Halteman, 2000). Par la suite, les assemblages macrofossiles témoignent d’un niveau d’eau à la baisse dans les deux lacs. Le minimum est atteint au lac du Triangle entre 6500 et 4000 BP ; il y est particulièrement souligné par l’abondance de Sphagnum et, dans une moindre mesure, celle des Cypéracées, traduisant ainsi le développement centripète d’une tourbière riveraine. Contrairement à la région de Mariana Lake, en Alberta (Hutton et al., 1994), l’augmentation de la représentation des sphaignes ne peut être ici attribuée à la multiplication des tourbières à l’échelle régionale ; le contexte physiographique l’interdit. Le niveau de la colonne d’eau devait alors être égal ou inférieur à 1,6 m, puisqu’il n’y a pas dépôt de marne au centre du lac actuellement. Au Petit lac Bouchard, c’est entre 6600 et 3000 BP que le niveau d’eau fut le plus bas. L’abondance des Isoetes et de Potamogeton pusillus indique que la profondeur d’eau au centre du lac devait se situer entre 25 cm et 1,5 m (Hannon et Gaillard,1997 ; Dieffenbacher-Krall et Halteman, 2000). Il est possible que l’abaissement du niveau d’eau se soit amorcé avant 6600 BP, comme en témoigne Potamogeton pusillus vers 7000 BP. Le lac du Triangle et le Petit lac Bouchard connaissent une remontée progressive du niveau de l’eau depuis 4000 BP et depuis 3000 BP, respectivement.
Figure 10

Résumé des changements du niveau d’eau du lac du Triangle (a) et du Petit lac Bouchard (b) avec les profondeurs estimées et les bas et hauts niveaux d’eau durant l’Holocène : (c) le lac Albion, dans le sud-ouest du Québec (Lavoie et Richard, 2000), (d) le lac des Pluviers, dans le Québec nordique (Payette et Filion, 1993), (e) le lac Decoy (Yu, 1995) et (f) le lac Crawford (Yu et al., 1997), au sud de l’Ontario. Les parties noires des barres indiquent des hauts niveaux d’eau, les parties blanches, des bas niveaux et les grises, des niveaux intermédiaires.
Lake-level changes synthesis of Lac du Triangle (a) and of Petit Lac Bouchard (b) with estimated depths and Holocene lake-levels for (c) Lac Albion, southwestern Québec (Lavoie et Richard, 2000), (d) Lac des Pluviers, northern Québec (Payette et Filion, 1993), (e) Decoy Lake (Yu, 1995) and (f) Crawford Lake (Yu et al., 1997), southern Ontario. Black bars are for high lake-levels, white bars, for low lake-levels, and grey bars, for intermediate lake-levels.
Seule une combinaison d’indices sédimentaires et biologiques provenant d’une série de profils disposés le long d’un transect latéral pour chacun des deux lacs permettrait de préciser l’amplitude et la fréquence des changements du niveau d’eau (Digerfeldt, 1986 ; Dearing, 1997 ; Hannon et Gaillard, 1997). Néanmoins, les indices dont nous disposons sont suffisants pour détecter les changements principaux (fig. 10). Ils reflètent probablement des changements dans les précipitations ayant influencé le niveau moyen de la nappe phréatique dans le bassin versant de chaque lac, étant donné la nature relativement poreuse du substratum rocheux.
Le synchronisme relatif entre le centre et l’est de la Gaspésie traduit un changement du bilan hydrique à l’échelle régionale attribuable à une modification du patron de la circulation atmosphérique (Harrison et Metcalfe, 1985 ; Digerfeldt, 1986, 1988 ; Harrison et Digerfeldt, 1993 ; Webb et al., 1993 ; Yu, 1995). Le climat aurait donc été plus sec à l’Holocène moyen, entre 6500 et 4000-3000 BP. Ce décalage de 1000 ans à l’Holocène supérieur pourrait refléter une réponse plus lente du Petit lac Bouchard attribuable à l’influence maritime se manifestant en bordure de la péninsule, mais il pourrait aussi refléter des conditions locales. Seule la multiplication d’études similaires permettra de trancher.
Le bas niveau d’eau enregistré à nos deux lacs correspond relativement bien à un épisode où on a observé de bas niveaux d’eau au sud du Québec, entre 6100 et 4400 BP (Lavoie et Richard, 2000), au Québec subarctique, entre 6300 et 3700 BP (Payette et Filion, 1993), et au sud de l’Ontario, entre 7000 et 3000 BP (Yu, 1995 ; Yu et al., 1997). Lavoie et Richard (2000) ont expliqué l’existence de ce bas niveau par un climat plus sec attribuable à une incursion plus fréquente de la masse d’air du Pacifique, modifiée par le passage sur les Rocheuses. L’Holocène supérieur est caractérisé par une remontée généralisée du niveau d’eau dans l’ensemble de ces régions, soit depuis au moins 3500 BP, traduisant un climat plus humide. Bien que préliminaires, ces correspondances montrent que les changements du bilan hydrique sont relativement synchrones et couvrent l’ensemble du Québec (Bégin et Payette, 1988 ; Payette et Delwaide, 1991 ; Bégin, 2001).
Les changements du niveau d’eau ont par ailleurs eu des conséquences sur l’accumulation pollinique au sein du lac du Triangle et du Petit lac Bouchard, dont les influx reflètent non seulement les changements dans l’abondance des populations végétales autour des bassins, mais aussi le dépôt et le redépôt entraînés par les changements de morphométrie et, partant, de focalisation pollinique. Dans le cas de petits lacs, de tels changements taphonomiques doivent être considérés pour tirer pleinement parti, comme nous l’avons fait, des taux d’accumulation pollinique.
Climat, feux et dynamique des populations arborescentes
C’est durant l’Holocène moyen que le niveau des lacs (fig. 10) et l’influx des charbons de bois (fig. 4) furent les plus bas. Un climat moins pluvieux correspondrait ainsi à un petit nombre de feux. Ce paradoxe fut résolu par Carcaillet et Richard (2000) dans leur étude des climats saisonniers et de la fréquence des sécheresses, cause des incendies. Malgré des précipitations hivernales faibles entraînant de bas niveaux lacustres, les masses d’air étaient stables durant l’Holocène moyen et, partant, les épisodes de sécheresse, rares. Les feux aussi. Au contraire, l’instabilité des masses d’air caractérise l’Holocène inférieur et l’Holocène supérieur ; le nombre des feux s’est accru (fig. 4) en raison de la fréquence élevée des épisodes de sécheresse estivale même si les précipitations sont plus fortes, notamment durant l’Holocène supérieur.
L’établissement, puis le développement des populations arborescentes se sont effectués autour du lac du Triangle et du Petit lac Bouchard (fig. 4) alors que les feux étaient fréquents, mais c’est dans la région de Gaspé que l’afforestation s’est poursuivie le plus longtemps, jusque vers 7700 BP. Depuis environ 9500 BP, le sapin baumier abondait au coeur de la péninsule tandis que, dans la région de Gaspé, le tremble et l’aulne crispé ont dominé pendant encore plus de 1500 ans, durant une période où les feux étaient moins fréquents qu’auparavant d’après l’examen des influx de charbons de bois (fig. 4). Il se peut toutefois que la fréquence des incendies soit restée élevée. Quoi qu’il en soit, les différences entre les deux sites ne s’expliquent pas par la seule fréquence des feux. Un climat plus froid qu’à l’intérieur des terres aurait occasionné la persistance de cette afforestation, car rien n’empêchait alors les arbres d’atteindre la région. Les conditions climatiques de la péninsule gaspésienne, aujourd’hui plus clémentes sur le pourtour qu’au centre, étaient donc alors inversées. L’abondance du tremble et de l’aulne crispé signalerait l’existence d’un climat moins humide (Thompson et al., 1999a, b).
Le passage de l’Holocène moyen à l’Holocène supérieur, vers 3500 BP, est identifié par une augmentation de la fréquence des feux (fig. 4) et une élévation sans doute progressive des niveaux lacustres (fig. 10). Le climat est sans doute devenu plus froid et plus humide. Les sapinières à épinettes noires du centre de la Gaspésie ont alors surtout connu une augmentation de la proportion d’épinettes, bien que le sapin baumier ait aussi réagi positivement, mais au détriment des populations de bouleau blanc semble-t-il (fig. 7a). Dans le même temps, les bétulaies jaunes à sapins de la région de Gaspé se sont surtout enrichies de sapins baumiers et d’épinettes blanches et, dans une moindre mesure, d’épinettes noires au détriment des populations de bouleau jaune. Dans les deux régions, l’enrésinement des paysages végétaux caractérise l’Holocène supérieur. Ces changements sont semblables à ceux observés aujourd’hui entre les sous-domaines bioclimatiques de la Sapinière à bouleau jaune de l’ouest et de l’est du Québec, respectivement (Grondin, 1996). Les populations de pin blanc, d’érable à sucre et de chêne rouge, parfois présentes en abondance dès l’Holocène moyen, ont fluctué à la baisse mais se sont tout de même maintenues dans la région de Gaspé. L’augmentation de la fréquence des feux a sans doute contribué à l’appauvrissement des populations d’arbres thermophiles en région côtière.
Le cas de l’érable à sucre est intéressant, car ses populations sont maximales à la charnière entre les deux périodes et il était présent dans la région de Gaspé dès 5000 BP, 1000 ans plus tôt qu’à Mont-Saint-Pierre (Labelle et Richard, 1984). L’hypothèse de Dansereau (1944) selon laquelle les érablières à Acer saccharum de la Gaspésie seraient des groupements reliques hérités de l’optimum climatique de l’Holocène moyen, période au cours de laquelle les érablières auraient été largement répandues sur le territoire, est donc aussi réfutée pour le coeur de la Gaspésie. Plus généralement, l’hypothèse selon laquelle les taxons thermophiles aurait connu une plus grande extension durant l’optimum climatique de l’Holocène moyen, encore répandue pour expliquer certains traits de l’aire de peuplement des végétaux vasculaires dans l’Est du Canada (Clayden, 2000), pourrait devoir être révisée, du moins quand on y fait intervenir des changements à l’échelle des biomes ou des domaines écologiques. Le cas du Petit lac Bouchard montre toutefois des changements dans l’abondance de tels taxons, mais leur aire de peuplement ne paraît pas avoir été beaucoup plus étendue dans le passé.
L’établissement du pin gris dans la région du lac du Triangle durant la phase d’afforestation correspond à une période de grande fréquence des feux, ce qui est conforme à l’écologie de l’essence (Fowells, 1965). Sa persistance durant l’Holocène supérieur, période où les feux étaient peu nombreux (fig. 4), montre que la relation entre le feu et l’abondance du pin gris n’est pas simple et rappelle le cas des peuplements côtiers du Maine qui, manifestement, se reproduisent abondamment en climat maritime, en l’absence d’incendies fréquents. Il s’agit d’écotypes différents.
Conclusion
L’étude des sédiments du lac du Triangle et du Petit lac Bouchard a permis de définir la paléophytogéographie du centre et de l’est de la Gaspésie et d’en préciser le contexte paléoclimatique. La déglaciation tardive aura favorisé une colonisation rapide des terres, mais le délai dans la constitution des forêts fermées derrière Gaspé signale la persistance d’un climat côtier froid. Les arbres dominants des forêts boréales actuelles (Picea, Abies, Betula papyrifera, Larix) sont présents partout, dès le début de l’Holocène. Les essences thermophiles occupant actuellement le pourtour côtier de la péninsule n’ont jamais pénétré profondément à l’intérieur des terres. Notre hypothèse selon laquelle le bouleau jaune (Betula alleghaniensis) abondait toutefois dans la région de Gaspé durant l’Holocène moyen est donc vérifiée. Le pin gris (Pinus banksiana), aujourd’hui absent de la Gaspésie, en occupait la partie centrale, entre 10 000 et 4000 ans avant nos jours.
Durant l’afforestation postglaciaire, la zone pollinique à dominance d’Alnus crispa s’étend à l’ensemble de la Gaspésie, mais c’est dans les régions côtières que l’aulne crispé a connu sa plus forte abondance. Le rôle du feu dans le maintien, voire le développement des populations de l’aulne crispé et dans le déclin de celles des épinettes est confirmé, mais d’éventuels changements dans la fréquence des feux devront être établis, non seulement pour la période d’afforestation mais pour l’ensemble du postglaciaire, afin d’examiner la réponse éventuelle des populations arborescentes à ce facteur clé de l’écologie boréale.
Nos reconstitutions paléohydrologiques s’ajoutent aux données encore trop peu nombreuses disponibles pour le Québec. Elles montrent que les forêts du centre, autant que celles de l’est, ont connu un climat plus sec à l’Holocène moyen, entre 6000 et 3500 BP. D’autres études portant sur les niveaux lacustres sont nécessaires pour préciser la paléohydrologie régionale de la Gaspésie.
Appendices
Remerciements
Ces recherches furent menées dans le cadre du projet CSHD (Climate System History and Dynamics) soutenu par le Conseil de recherche en sciences naturelles et en génie du Canada. Nous remercions Mme Nicole Morasse et M. Alayn Larouche du Laboratoire de paléobiogéographie et de palynologie du Département de géographie de l’Université de Montréal, pour leur contribution aux analyses polliniques et macrofossiles. Nous remercions également Mme Alwynne B. Beaudoin et les deux lecteurs anonymes pour leurs commentaires et suggestions.
Références
- Aaby, B. et Berglund, B.E., 1986. Characterization of peat and lake deposits, p. 231-246. In B.E. Berglund, édit., Handbook of Holocene Palaeoecology and Palaeohydrology. John Wiley and Sons, Chichester, 869 p.
- Asnong, H., 2001. Histoire postglaciaire de la végétation, des feux et des niveaux lacustres en Gaspésie, Québec. Mémoire de maîtrise, Université de Montréal, 186 p.
- Bégin, Y., 2001. Tree-ring dating of extreme lake levels at the subarctic-boreal interface. Quaternary Research, 55 : 133-139.
- Bégin, Y. et Payette, S., 1988. Dendroecological evidence of lake-level changes during the last three centuries in subarctic Québec. Quaternary Research, 30 : 210-220.
- Bennett, K.D. et Fuller, J.L., 2002. Determining the age of the mid-Holocene Tsuga canadensis (hemlock) decline, eastern North America. The Holocene, 12 : 421-429.
- Benninghoff, W.S., 1962. Calculation of pollen and spores density in sediments by addition of exotic pollen in known quantities. Pollen et Spores, 6 : 332-333.
- Birks, H.H., 1980. Plant macrofossils in Quaternary lake sediments. Archiv für Hydrobiologie Beih., Ergebnisse Limnologiae, 15 : 1-60.
- Carcaillet, C. et Richard, P.J.H., 2000. Holocene changes in seasonal precipitation highlighted by fire incidence in eastern Canada. Climate Dynamics, 16 : 549-559.
- Clayden, S.R., 2000. History, physical setting, and regional variation of the flora, p. 35-73. In H.R. Hinds, édit., Flora of New Brunswick. 2e éd. Biology Department, University of New Brunswick, Fredericton, 699 p.
- Comtois, P., 1981. Diamètre pollinique de quelques espèces tourbicoles de Betula. Le Naturaliste canadien, 108 : 471-474.
- Comtois, P. et Payette, S., 1984. Représentation pollinique actuelle et subactuelle des peupleraies boréales au Nouveau-Québec. Géographie physique et Quaternaire, 38 : 123-133.
- Cushing, E.J., 1967. Evidence for differential Pollen Preservation in late Quaternary sediments in Minnesota. Review of Palaeobotany and Palynology, 4 : 87-101.
- Dansereau, P., 1944. Les érablières de la Gaspésie et les fluctuations du climat. Bulletin des sociétés de géographie de Québec et de Montréal, 3 : 1-18.
- Davis, M.B., 1968. Pollen grains in lake sediments: Redeposition caused by seasonal water circulation. Science, 162 : 796-799.
- ——— 1973. Redeposition of pollen grains in lake sediment. Limnology and Oceanography, 18 : 44-52.
- ——— 1981. Outbreak of forest pathogens in Quaternary history, p. 216-227. In D.C. Bharadwaj, édit., Proceedings of the 4th International Conference on Palynology (Lucknow, Inde, 1976-1977), vol. 3, Lucknow.
- Davis, M.B. et Brubaker, L.B., 1973. Differential sedimentation of pollen grains in lakes. Limnology and Oceanography, 18 : 635-646.
- Davis, M.B., Brubaker, L.B. et Webb III, T., 1973. Calibration of absolute pollen influx, p. 9-25. In H.J.B. Birks et R.G. West, édit., Quaternary Plant Ecology: The 14th Symposium of the British Ecological Society. Wiley, Toronto, 326 p.
- Davis, M.B., Moeller, R.E. et Ford, J., 1984. Sediment focusing and pollen influx, p. 261-293. In E.Y. Haworth et J.W.G. Lund, édit., Lake Sediments and Environmental History. University of Leicester Press, Leicester, 411 p.
- Dean Jr., W.E., 1974. Determination of carbonate and organic matter in calcareous sediments an sedimentary rocks by loss on ignition: Comparison with other methods. Journal of Sedimentary Petrology, 44 : 242-248.
- Dean, W.E. et Schwalb, A., 2002. The lacustrine carbon cycle as illuminated by the waters and sediments of two hydrologically distinct headwater lakes in North-Central Minnesota, U.S.A. Journal of Sedimentary Research, 72 : 416-431.
- Dearing, J.A., 1997. Sedimentary indicators of lake-level changes in the humid temperate zone: A critical review. Journal of paleolimnology, 18 : 1-14.
- Delcourt, P.A., Delcourt, H.R. et Webb III, Th., 1984. Atlas of Mapped Distributions of Dominance and Modern Pollen Percentages for Important Tree Taxa of Eastern North America. American Association of Stratigraphic Palynologists, Foundation. Contributions Series, 14, s.l., 131 p.
- Dieffenbacher-Krall, A.C. et Halteman, W.A., 2000. The relationship of modern plant remains to water depth in alkaline lakes in New England, USA. Journal of Paleolimnology, 24 : 213-229.
- Digerfeldt, G., 1986. Studies on past lake-level fluctuations, p. 127-143. In B.E. Berglund, édit., Handbook of Holocene Palaeoecology and Palaeohydrology. John Wiley and Sons, Chichester, 869 p.
- ——— 1988. Reconstruction and regional correlation of Holocene lake-level fluctuations in Lake Bysjön, South Sweden. Boreas, 17 : 165-182.
- Environnement Canada, 2003. Information climatique et hydrologique du Canada : Québec. Site internet : http://www.msc.ec.gc.ca/climate/climate_normals_1990/show_stations_f.cfm?province=QC (Page consultée le 18 mai 2004).
- Faegri, K., Kaland, P.E. et Krzywinski, K.J., 1989. Textbook of Pollen Analysis. 4e édit. John Wiley and Sons, New York, 328 p.
- Farrar, J.L., 1996. Les Arbres du Canada. Ressources naturelles Canada, Service canadien des forêts et Fides, Saint-Laurent, 502 p.
- Fowells, H.A., 1965. Silvics of Forest Trees of the United States. United States Department of Agriculture, Forest service, Washington, D.C., Agriculture Handbook 271, 762 p.
- Fuller, J.L., 1998. Ecological impact of the mid-Holocene Hemlock decline in southern Ontario, Canada. Ecology, 79 : 2337-2351.
- Gajewski, K., 1991. Représentation pollinique actuelle à la limite des arbres au Nouveau-Québec. Canadian Journal of Earth Science, 28 : 643-648.
- Glew, J.R., 1988. A portable extruding device for close interval sectioning of unconsolitated core samples. Journal of Paleolimnology, 1 : 235-239.
- ——— 1989. A new trigger mechanism for sediment samplers. Journal of Paleolimnology, 2 : 241-243.
- Grandtner, M.M., 1972. Aperçu de la végétation du Bas-Saint-Laurent, de la Gaspésie et des Îles-de-le-Madeleine. Cahiers de géographie du Québec, 16 : 116-121.
- Grimm, E.C., 1991. TILIA and TILIA-GRAPH. Illinois State Museum, Springfield, Logiciel.
- Grondin, P., 1996. Cadre bioclimatique de référence (chapitre Écologie forestière, pro parte), p. 148-159. In J.A. Bérard et M. Côté, édit, Manuel de foresterie. Ordre des ingénieurs forestiers du Québec, Les Presses de l’Université Laval, Québec, 1428 p.
- Hannon, G.E. et Gaillard, M-J., 1997. The plant-macrofossil record of past lake-level changes. Journal of Paleolimnology, 18 : 15-28.
- Harrison, S.P. et Digerfeldt, G., 1993. European lakes as palaeohydrological and palaeoclimatic indicators. Quaternary Science Reviews, 12 : 233-248.
- Harrison, S.P. et Metcalfe, S.E., 1985. Variations in lake levels during the Holocene in North America: An indicator of changes in atmospheric circulation patterns. Géographie physique et Quaternaire, 39 : 141-150.
- Heinselman, M.L., 1981. Fire and succession in the conifer forests of northern North America, p. 374-405. In D.C. West, H.H. Shugart et D.B. Botkin, édit., Forest Succession: Concepts and Application. Springer-Verlag, New York, 517 p.
- Huss-Danell, K., Sverrisson, H., Hahlin, A.-S. et Danell, K., 1999. Occurence of Alnus-infective Frankia and Trifolium-infective Rhizobium in circumpolar soils. Arctic, Antarctic and Alpine Research, 31 : 400-406.
- Hutton, M.J., MacDonald, G.M. et Mott, R.J., 1994. Postglacial vegetation history of the Mariana Lake region, Alberta. Canadian Journal of Earth Sciences, 31 : 418-425.
- Janssen, C.R., 1966. Recent pollen spectra from the deciduous and coniferous-decidous forests of Northeastern Minnesota: A study in pollen dispersal. Ecology, 47 : 804-825.
- Jetté, H. et Richard, P.J.H., 1992. Contribution à l’histoire postglaciaire de la végétation en Gaspésie méridionale, Québec. Géographie physique et Quaternaire, 46 : 273-284.
- King, G.A., 1986. Deglaciation and vegetation history of western Labrador and adjacent Québec. Thèse de doctorat, University of Minnesota, 266 p.
- Labelle, C. et Richard, P.J.H., 1981. Végétation tardiglaciaire et postglaciaire au sud-est du parc des Laurentides, Québec. Géographie physique et Quaternaire, 35 : 345-359.
- ——— 1984. Histoire postglaciaire de la végétation dans la région du Mont-Saint-Pierre, Gaspésie, Québec. Géographie physique et Quaternaire, 38 : 257-274.
- Lamb, H.F., 1980. Late-Quaternary vegetation history of southeastern Labrador. Arctic and Alpine Research, 12 : 117-135.
- Larouche, A.C., 1979. Histoire postglaciaire comparée de la végétation à Sainte-Foy et au Mont des Éboulements, Québec, par l’analyse macrofossile et l’analyse pollinique. Mémoire de maîtrise, Université Laval, 117 p.
- Lavoie, M. et Richard, P.J.H., 2000. Postglacial water-level changes of a small lake in southern Québec, Canada. The Holocene, 10 : 637-650.
- Lévesque, P.E.M., Dinel, H. et Larouche, A., 1988. Guide illustré des macrofossiles végétaux des tourbières du Canada. Agriculture Canada, Direction générale de la recherche, Centre de recherche sur les terres, Ottawa, Publication 1817, 65 p.
- Litherland, A.E. et Beukens, R.P., 1995. Radiocarbon dating by atom counting, p. 117-123. In N.W. Rutter et N.R. Catto, édit., Dating Methods for Quaternary Deposits. Geological Association of Canada, St. John’s, GEO Text 2, 308 p.
- Livingstone, D.A., 1955. A lightweight piston sampler for lake deposits. Ecology, 36 : 136-137.
- ——— 1968. Some interstadial and postglacial pollen diagrams from eastern Canada. Ecological Monographs, 38: 87-125.
- Lortie, M., 1979. Arbres, forêts et perturbations naturelles au Québec. Presses de l’Université Laval, Sainte-Foy, 172 p.
- Majcen, Z., 1981. Les forêts du parc national Forillon, Gaspésie, Québec : étude phytosociologique. Laboratoire d’écologie forestière, Études écologiques 4, Université Laval, Québec, 158 p.
- Marcoux, N. et Richard, P.H.J., 1993. A new look at Betula pollen curves. INQUA-Commission for the Study of the Holocene, Working Group on Data Handling Methods, Newsletter, 10 : 6-8.
- ——— 1995. Végétation et fluctuations climatiques sur la côte septentrionale gaspésienne, Québec. Canadian Journal of Earth Science, 32 : 79-96.
- McAndrews, J.H., Berti, A.A. et Noris, G., 1973. Key to the Quaternary Pollen and Spores of the Great Lakes Region. Royal Ontario Museum, Life Sciences Miscellaneous Publications, Toronto, 61 p.
- Ministère de l’Énergie et des Ressources du Québec, 1987. Carte minérale de la région du Bas-Saint-Laurent et de la péninsule de Gaspé. Direction générale de l’exploitation géologique et minérale, Service de la promotion et de l’aide à l’exploration minière, 1 carte (à env. 1/1 800 000).
- Moore, P.D., Webb, J.A. et Collinson, M.E., 1991. Pollen Analysis. Blackwell Scientific, Oxford, 216 p.
- Mott, R.J., 1976. A Holocene pollen profile from the Sept-Iles area, Québec. Le Naturaliste canadien, 103 : 457-467.
- ——— 1978. Populus in Late-Pleistocene pollen spectra. Canadian Journal of Botany, 56 : 1021-1031.
- Nielsen, H. et Sorensen, I., 1992. Taxonomy and stratigraphy of late-glacial Pediastrum taxa from Lysmosen, Denmark: A preliminary study. Review of Palaeobotany and Palynology, 74 : 55-75.
- Payette, S. et Delwaide, A., 1991. Variations séculaires du niveau d’eau dans le bassin de la rivière Boniface (Québec nordique): une analyse dendroécologique. Géographie physique et Quaternaire, 45 : 59-67.
- Payette, S. et Filion, L. 1993. Holocene water-level fluctuations of a subarctic lake at the tree line in northern Québec. Boreas, 22 : 7-14.
- Proulx, H., Jacques, G., Lamothe, A-M. et Litynski, J., 1987. Climatologie du Québec méridional. Ministère de l’Environnement du Québec, Direction de la météorologie, Québec, M.P. 65, 131 p.
- Richard, P.J.H., 1968. Un spectre pollinique type de la sapinière à bouleau blanc pour la Forêt Montmorency. Le Naturaliste canadien, 95 : 565-576.
- ——— 1970. Atlas pollinique des arbres et de quelques arbustes indigènes du Québec. Le Naturaliste canadien, 97 : 1-34 ; 97-161 ; 241-306.
- ——— 1976. Relations entre la végétation actuelle et le spectre pollinique au Québec. Le Naturaliste canadien, 103 : 53-66.
- ——— 1977. Histoire post-wisconsinienne de la végétation du Québec méridional par l’analyse pollinique. Ministère des Terres et Forêts du Québec, Service de la recherche, Direction générale des forêts, Publications et rapports divers, tome 1, 312 p. ; tome 2, 142 p.
- ——— 1979. Contribution à l’histoire postglaciaire de la végétation au nord-est de la Jamésie, Nouveau-Québec. Géographie physique et Quaternaire, 33 : 93-112.
- ——— 1981. Paléophytogéographie postglaciaire en Ungava, par l’analyse pollinique. Laboratoire d’archéologie de l’Université du Québec à Montréal, Montréal, Paléo-Québec, 13, 153 p.
- ——— 1993. Origine et dynamique postglaciaire de la forêt mixte au Québec. Review of Palaeobotany and Palynology, 79 : 31-68.
- ——— 1994. Postglacial palaeophytogeography of the eastern St. Lawrence River Watershed and the climatic signal of the pollen record. Palaeogeography, Palaeoclimatology, Palaeoecology, 109 : 137-161.
- Richard, P.J.H. et Labelle, C., 1989. Histoire postglaciaire de la végétation au lac du Diable, Mont Albert, Gaspésie, Québec. Géographie physique et Quaternaire, 43 : 337-354.
- Richard, P.J.H. et Larouche, A.C., 1994. Histoire postglaciaire de la végétation et du climat dans la région de Rimouski, Québec. Paléo-Québec, 22 : 51-89.
- Richard, P.J.H., Larouche, A.C. et Lortie, G., 1992. Paléophytogéographie et paléoclimats postglaciaires dans l’ouest du Bas-Saint-Laurent, Québec. Géographie physique et Quaternaire, 46 : 151-172.
- Richard, P.J.H. et Poulin, P., 1976. Un diagramme pollinique au Mont des Éboulements, région de Charlevoix, Québec. Canadian Journal of Earth Science, 13 : 145-156.
- Richard, P.J.H., Veillette, J.J., Larouche, A.C., Hétu, B., Gray, J.T. et Gangloff, P., 1997. Chronologie de la déglaciation en Gaspésie: nouvelles données et implications. Géographie physique et Quaternaire, 51 : 163-184.
- Rousseau, C., 1974. Géographie floristique du Québec-Labrador. Centre d’études nordiques, Travaux et documents 7, Les Presses de l’Université Laval, Québec, 799 p.
- Saucier, J.-P. et Grondin, P., 1996. Domaine de la sapinière à bouleau jaune (chapitre Écologie forestière, pro parte), p. 197-208. In J.A. Bérard et M. Côté, édit., Manuel de foresterie. Ordre des ingénieurs forestiers du Québec et Les Presses de l’Université Laval, Québec, 1428 p.
- Savoie, L., 1978. Paléophytogéographie de l’épisode de Saint-Narcisse dans la région de Sainte-Agathe. Mémoire de maîtrise, Université de Montréal, 106 p.
- Scoggan, H.J., 1950. The Flora of Bic and the Gaspé Peninsula, Québec. Bulletin of the National Museum of Canada (Biological series 39), 115 : 399 p.
- Smol, J.P., 1990. Freshwater Algae, p. 3-14. In B.G. Warner, édit., Methods in Quaternary Ecology. Geological Association of Canada, St. John’s, Geoscience Canada Reprint Series 5, 170 p.
- Stuiver, M. et Reimer, P.J., 1993. Extended 14C data base and revised Calib 3.0 14C age calibration program. Radiocarbon, 35 : 215-230.
- Thompson, R.S., Anderson, K.H. et Bartlein, P.J., 1999a. Atlas of Relations Between Climatic Parameters and Distributions of Important Trees and Shrubs in North America. A - Introduction and Conifers. U.S. Geological Survey, Denver, Professional Paper 1650-A, 269 p.
- ——— 1999b. Atlas of Relations Between Climatic Parameters and Distributions of Important Trees and Shrubs in North America. B - Hardwoods. U.S. Geological Survey, Denver, Professional Paper 1650-B, 423 p.
- Troëls-Smith, J., 1955. Characterization of unconsolidated sediments. Danmarks Geologiske Undersogelse, ser. IV, 3 (10) : 37-73.
- Veillette, J.J. et Cloutier, M., 1993. Géologie des formations en surface, Gaspésie, Québec. Commission géologique du Canada, carte 1804A, échelle 1/250 000.
- Webb III, T., Bartlein, P.J., Harrison, S.P. et Anderson, K.H., 1993. Vegetation, lake levels, and climate in eastern North America for the past 18,000 years. p. 415-467. In H.E. Wright, J.E. Kutzbach, F.A. Street-Perrott, W.F. Ruddiman et T. Webb III, edit., Global Climates since the Last Glacial Maximum. University of Minnesota Press, Minneapolis, 569 p.
- Webb III, T., Yeracaris, G.Y. et Richard, P.J.H., 1978. Mapped patterns in sediment samples of modern pollen form southeastern Canada and northeastern United States. Géographie physique et Quaternaire, 32 : 163-176.
- Watts, W.A., 1978. Plant macrofossils and Quaternary paleoecology, p. 53-66. In D. Walker et J.C. Guppy, édit., Biology and Quaternary Environments. Australian Academy of Sciences, Symposium on Biological Problems in the Reconstruction of Quaternary Terrestrial Environments (Canberra, 1978), Canberra, 264 p.
- Young J.A. et Young, C.G., 1992. Seeds of Woody Plants in North America. Xe éd., Dioscorides Press, Portland, 407 p.
- Yu, Z. 1995. Mid-Holocene dry period in southern Ontario? responses of lake hydrology and upland pollen to climate change, p. 97-104. In I.D. Campbell, C. Campbell, D. Lemmen et B. Vance, édit., Proceedings: Climate, Landscape, and Vegetation Change in the Canadian Prairie Provinces. Canadian Forest Service, Edmonton.
- Yu, Z., McAndrews, J.H. et Eicher, U., 1997. Middle Holocene dry climate caused by change in atmospheric circulation patterns: evidence from lake levels and stable isotopes. Geology, 25 : 251.
 10.7202/021024ar
10.7202/021024arList of figures
Figure 1
Sites des reconstitutions paléophytogéographiques réalisées en Gaspésie et localisation des sites de l’étude : (1) lac Turcotte et (2) lac à Léonard (Labelle et Richard, 1984), (3) lac du Diable (Richard et Labelle, 1989), (4) lac Harriman, (5) lac Caribou, (6) lac à Robin et (7) lac à Raymond (Jetté et Richard, 1992), (8) lac à Euloge et lac J’Arrive (9) (Marcoux et Richard, 1995), (10) lac du Triangle et (11) Petit lac Bouchard (cette étude).
Map of Gaspésie showing location of studied sites for paleophytogeographical reconstruction of this paper and other studies: (1) Lac Turcotte and (2) Lac à Léonard (Labelle et Richard, 1984), (3) Lac du Diable (Richard et Labelle, 1989), (4) Lac Harriman, (5) Lac Caribou, (6) Lac à Robin et (7) Lac à Raymond (Jetté et Richard, 1992), (8) Lac à Euloge et Lac J’Arrive (9) (Marcoux et Richard, 1995), (10) Lac du Triangle et (11) Petit Lac Bouchard (this paper).
a
Le lac du Triangle, à la tête de la rivière Saint-Jean.
Lac du Triangle, at the head of Rivière Saint-Jean.
b
Le Petit lac Bouchard, à la tête de la rivière Dartmouth.
Petit lac Bouchard, at the head of Rivière Dartmouth.
Topographie des sites d’étude
Maps that show the topography of studied sites
Figure 3
Nature des sédiments (selon Aaby et Berglund, 1986), courbes de la perte au feu à 600 et 1 000 °C et des résidus et zones polliniques du lac du Triangle (a) et du Petit lac Bouchard (b).
Sediment stratigraphy (according to Aaby and Berglund, 1986), loss on ignition curves at 600 and 1 000 °C with regional pollen zone of Lac du Triangle (a) and Petit Lac Bouchard (b).
a
b
Taux d’accumulation des charbons microscopiques (mm2/cm2/an) et zones d’assemblage pollinique (ZAP) : (a) lac du Triangle et (b) Petit lac Bouchard. Chronologie en années étalonnées BP (années sidérales).
Microscopic charcoal accumulation rates (mm2/cm2/an) and regional pollen zones assemblages: (a) Lac du Triangle and (b) Petit Lac Bouchard. Chronology is in calibrated years BP (sideral years).
a
b
Courbes de l’âge en fonction de la profondeur des sédiments du lac du Triangle (a) et du Petit lac Bouchard (b). Les dates sont en années étalonnées BP. Les carrés noirs correspondent aux dates au radiocarbone par spectrométrie de masse à l’accélérateur de particules (SMA) de TRIANGLE ou de BOUCH96, les carrés blancs, aux dates SMA de BOUCHARDB, les losanges noirs et les triangles blancs, aux dates au radiocarbone conventionnelles de BOUCHS et BOUH96 respectivement. Les cercles représentent les dates par corrélation sporopollinique.
Age/depth curves of sediment of Lac du Triangle (a) and Petit Lac Bouchard (b). Black squares correspond to dates from accelerator mass spectrometry (AMS) for TRIANGLE or BOUCH96; white squares to AMS dates for BOUCAHRDB; black lozenges and white triangles, to standard radiocarbon dates for BOUCHS and BOUCH96. Circle are dates from pollen correlation.
a
Lac du Triangle
Lac du Triangle
b
Petit Lac Bouchard
Petit Lac Bouchard
Diagrammes des pourcentages polliniques : (a) lac du Triangle (48° 42’ 36’’ N, 65° 24’ 50’’ O), (b) Petit lac Bouchard (48° 51’ 14’’ N, 64° 35’ 52’’ O).
Pollen and spore percentages diagrams: (a) Lac du Triangle (48° 42’ 36’’ N, 65° 24’ 50’’ W), (b) Petit Lac Bouchard (48° 51’ 14’’ N, 64° 35’ 52’’ W).
a
Lac du Triangle
Lac du Triangle
b
Petit Lac Bouchard
Petit Lac Bouchard
Diagrammes des influx polliniques : (a) lac du Triangle, (b) Petit lac Bouchard. Les courbes d’influx polliniques sont brutes, non corrigées.
Pollen accumulation rates diagrams: (a) Lac du Triangle, (b) Petit Lac Bouchard. Pollen accumulation rates curves were made with raw data (non-corrected data).
a
Lac du Triangle
Lac du Triangle
b
Petit Lac Bouchard
Petit Lac Bouchard
Courbes des pourcentages de Betula selon le diamètre équatorial des grains de pollen du lac du Triangle (a) et du Petit lac Bouchard (b).
Betula pollen grain size percentages diagrams of Lac du Triangle (a) and Petit Lac Bouchard (b).
a
Lac du Triangle
Lac du Triangle
b
Petit Lac Bouchard
Petit Lac Bouchard
Diagrammes des concentrations macrofossiles : a) lac du Triangle, b) Petit lac Bouchard (ath : anthère, bra : bractée, brf : bractée femelle, cac : coussinet d’aiguille de conifère, cap : capsule, col : colonie, com : cône mâle, ecf : écaille de cône femelle, ecm : écaille de cône mâle, ecr : écorce, eph : ephippia, feu : feuille, ffeu : fragment de feuille, fgra : fragment de graine, fru : fruit, gem : gemmule, gra : graine, meg : mégaspore, mes : mésoblaste, oosp : oospore, rac : racine, ram : rameau, rhi : rhizome, scl : sclérote, sta : statoblaste, tif : tige feuillée, tig : tige).
Macrofossil-concentration diagrams: a) Lac du Triangle, b) Petit Lac Bouchard (ath: anther, bra: bract, brf: female bract, cac: coniferous sterigmata, cap: capsule, col: colony, com: male cone, ecf: female cone scale, ecm: male cone scale, ecr: bark, eph: ephippia, feu: leaf, ffeu: leaf fragment, fgra: seed fragment, fru: fruit, gem: gemmule, gra: seed, meg: megaspore, mes: mesoblast, oosp: oospore, rac: root, ram: branchlet, rhi: rhizome, scl: sclerotium, sta: statoblast, tif: leafy stem, tig: stem).
Figure 10
Résumé des changements du niveau d’eau du lac du Triangle (a) et du Petit lac Bouchard (b) avec les profondeurs estimées et les bas et hauts niveaux d’eau durant l’Holocène : (c) le lac Albion, dans le sud-ouest du Québec (Lavoie et Richard, 2000), (d) le lac des Pluviers, dans le Québec nordique (Payette et Filion, 1993), (e) le lac Decoy (Yu, 1995) et (f) le lac Crawford (Yu et al., 1997), au sud de l’Ontario. Les parties noires des barres indiquent des hauts niveaux d’eau, les parties blanches, des bas niveaux et les grises, des niveaux intermédiaires.
Lake-level changes synthesis of Lac du Triangle (a) and of Petit Lac Bouchard (b) with estimated depths and Holocene lake-levels for (c) Lac Albion, southwestern Québec (Lavoie et Richard, 2000), (d) Lac des Pluviers, northern Québec (Payette et Filion, 1993), (e) Decoy Lake (Yu, 1995) and (f) Crawford Lake (Yu et al., 1997), southern Ontario. Black bars are for high lake-levels, white bars, for low lake-levels, and grey bars, for intermediate lake-levels.
List of tables
Tableau I
Données sur les lacs étudiés et leurs sédiments
Tableau II
Données chronologiques
Profondeurs ajustées par corrélation des carottes ; voir texte.
Le δ13C des dates TO- est normalisé à -25 ‰.
Abréviations des pièces macrofossiles : bra : bractée ; ecf : écaille de cône femelle ; ecm : écaille de cône mâle ; ffeu : fragment de feuille ; fru : fruit ; gra : graine ; mes : mésoblaste.
Tableau III
TRIANGLE : Zonation pollinique régionale
CP : concentration pollinique (grains/cm3) ; PA/T : rapport du pollen des arbres (incluant le genre Betula) au total des grains.
Tableau IV
BOUCHARD : Zonation pollinique régionale
CP : concentration pollinique (grains/cm3) ; PA/T : rapport du pollen des arbres (incluant le genre Betula) au total des grains.